


El objetivo principal de la presente investigación fue evaluar los efectos del agonista 5-HT2C Ro 60-0175 en la expresión de la sensibilización locomotora inducida por la administración de etanol. Además, también se evaluaron los efectos del antagonista 5-HT2C SB 242084 sobre los efectos del Ro 60-0175 en la sensibilización locomotora producida por etanol para determinar si los efectos del Ro 60-0175 resultan de una acción específica sobre los receptores 5-HT2C. Diferentes grupos de ratas se sometieron al desarrollo de sensibilización locomotora producida por etanol. En las pruebas de expresión de la sensibilización locomotora se evaluaron los efectos del Ro 60-0175 y del SB 242084 sobre la sensibilización locomotora producida por etanol. Adicionalmente se evaluó el pretratamiento con SB 242084 sobre los efectos del Ro 60-0175 en la sensibilización locomotora producida por etanol. Los resultados mostraron que el Ro 60-0175 disminuyó la sensibilización locomotora producida por etanol y que este efecto fue prevenido por el pretratamiento con SB 242084. Estos resultados sugieren que los receptores 5-HT2C juegan un papel modulatorio en la sensibilización locomotora producida por etanol.
The main goal of the present research was to evaluate the effects of 5-HT2C receptor agonist Ro 60-0175 on the expression of ethanol-induced locomotor sensitization. In order to determine if these effects result from a specific action of Ro 60-0175 on 5-HT2C receptors, we also examined the effects of the selective 5-HT2C receptor antagonist SB 242084 on Ro 60-0175's effects on the ethanol-induced locomotor sensitization. Different groups of rats were subjected to development of ethanol-induced locomotor sensitization. On the expression tests of the locomotor sensitization the effects of the Ro 60-0175 and SB 242084 on the ethanol-induced locomotor sensitization were evaluated. In addition, the pretreatment with SB 242084 on the effects of Ro 60-0175 on the ethanol-induced locomotor sensitization was also evaluated. The results showed that Ro 60-0175 produced a dose-dependent prevention of the expression of ethanol-induced locomotor sensitization. This effect was reversed by administration of SB 242084. These data suggest that 5-HT2C receptors play a regulatory role on ethanol-induced locomotor sensitization.
El abuso y la dependencia del alcohol etílico o etanol, también conocida como alcoholismo, es un problema de salud pública en el mundo (World Health Organization, 2014) y en México (Villatoro et al., 2016). El consumo crónico de etanol puede conducir a la tolerancia, la sensibilización, el síndrome de retirada y la adicción al etanol (Diana et al., 2003). Estos efectos se deben a que el etanol modula numerosos sistemas de neurotransmisión cerebral como la DA (Koob, 2013), el GABA (Koob et al., 1998), el Glu (Krystal, Petrakis, Mason, Trevisan y D'Souza, 2003) y la serotonina (LeMarquand, Pihl y Benkelfat, 1994; Sari, 2013), entre otros, en el sistema de la gratificación del cerebro.
El etanol tiene afinidad por el sitio alostérico de la subunidad α de los receptores GABAA localizados en las interneuronas GABAérgicas del área tegmental ventral (ATV) (Enoch, 2008), entre otros sitios. La activación de estos receptores por el etanol produce la inhibición de la actividad de las interneuronas GABAérgicas del ATV (Stobbs, Ohran, Lassen, Allison, Brown y Steffensen, 2004); como consecuencia de esta inhibición, las neuronas DAérgicas del ATV incrementan su liberación de DA en el núcleo accumbens (nAcc). Este incremento de DA en el nAcc es el responsable de las propiedades adictivas del etanol (Weiss, Lorang, Bloom y Koob, 1993). Recientemente, se ha planteado que los receptores 5-HT2C podrían modular los efectos adictivos del etanol y otras drogas de abuso (Wu et al., 2015). Estos receptores se encuentran localizados, entre otros sitios, en los cuerpos celulares de las interneuronas GABAérgicas del ATV (Bubar y Cunningham, 2007) y su activación disminuye la tasa de disparo de las neuronas DAérgicas del ATV, dando como resultado una reducción en la liberación de DA en las zonas terminales de la vía mesocorticolímbica de la DA, incluyendo el nAcc (Gobert et al., 2000; Millan, Dekeyne y Gobert, 1998; Di Matteo, di Giovanni, di Mascio y Esposito, 2000). En línea con lo anterior, se ha reportado que la activación o inhibición de los receptores 5-HT2C disminuye o incrementa, respectivamente, la autoadministración de etanol en ratas (Tomkins et al., 2002).
Un modelo conductual con ratas de laboratorio que podría agregar información adicional del papel de los receptores 5-HT2C en las propiedades adictivas del etanol es la sensibilización locomotora inducida por las drogas de abuso. La sensibilización locomotora se ha definido como el aumento progresivo y persistente de los efectos locomotores de las drogas de abuso, incluido el etanol, después de su administración repetida (Kalivas y Stewart, 1991), y también se ha evidenciado que refleja las neuroadaptaciones en el circuito de la gratificación que contribuyen a la adicción a las drogas de abuso y que modela algunos aspectos de las conductas adictivas como el deseo por la droga (Robinson y Berridge, 1993). La sensibilización locomotora puede separarse en 2 fases diferentes: el desarrollo (o iniciación) y la expresión. Durante el desarrollo se producen las alteraciones celulares e inmediatas que inducen la sensibilización locomotora, y la expresión es el producto de las consecuencias a largo plazo de estos eventos iniciales. Por lo tanto, el propósito del presente estudio fue investigar si el agonista de los receptores 5-HT2C Ro 60-0175 podría prevenir la expresión de la sensibilización locomotora inducida por etanol. Además, con la finalidad de averiguar si los efectos producidos por el Ro 60-0175 están regulados por los receptores 5-HT2C, también evaluamos si la administración del antagonista selectivo de los receptores 5-HT2C SB 242084 puede prevenir los efectos del Ro 60-0175 sobre la expresión de la sensibilización locomotora inducida por etanol.
MétodoAnimalesSe utilizaron 150 ratas macho Wistar de aproximadamente 120 días de edad y con un peso de 200 a 250g, provenientes del Bioterio General de la FES Iztacala-UNAM. Las ratas se alojaron individualmente en cajas-hogar de acrílico de 30×20×20cm, con acceso libre a agua y comida (Teklad LM485 Rat Diet by Harlan) y bajo un ciclo luz-oscuridad controlado (luz 8:00 am-8:00 pm) y a una temperatura ambiente de 21±1°C. Los procedimientos de cuidado y manejo de animales se realizaron conforme a la Norma Oficial Mexicana (NOM-062-00-1999), titulada Especificaciones técnicas para la producción, cuidado y uso de los animales de laboratorio.
FármacosEtanol al 99.95% (J. T. Baker, México), (S)-2-(6-cloro-5-fluoroin-dol-1-yl)-1-metiletilamina (Ro 60-0175, agonista 5-HT2C), 6-cloro-5-metil-1-[2-(2metilpiridil-3-oxi)-pirida-5-il carbamoil] indolina (SB 242084, antagonista selectivo 5-HT2C). Estos fármacos fueron adquiridos en Tocris Bioscience, Estados Unidos. Todos los fármacos se prepararon diariamente, se disolvieron en solución salina al 0.9% y se administraron ip a un volumen de 1.0ml/kg.
AparatosLa actividad locomotora de las ratas se cuantificó en cajas de actividad de 40×40×30cm (modelo ENV-515, MED-Associates, St. Albans, VT, Estados Unidos) equipadas con una matriz de 8×8 emisores de luz infrarroja colocada 4cm por encima del piso de la caja. Las interrupciones de la luz infrarroja se detectaron a través de una interfase que registró en intervalos de 5min en un ordenador de escritorio compatible con IBM.
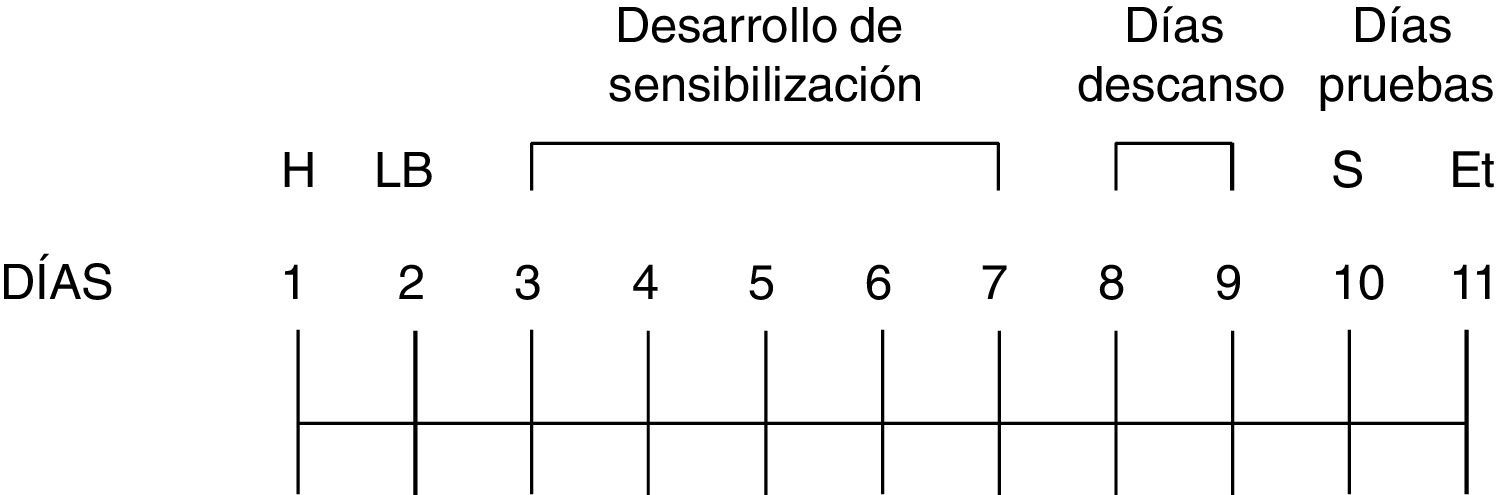
Procedimiento experimentalCada experimento se llevó a cabo en 3 fases: habituación a las cajas de actividad motora, desarrollo de la sensibilización motora y retos farmacológicos o días de prueba (fig. 1). En todos los ensayos, las ratas se colocaban en la caja de actividad durante 15min antes de cada sesión para disminuir la conducta exploratoria. Una vez transcurridos los 15min, las ratas fueron regresadas a su caja-hogar para la administración de los compuestos, dependiendo del experimento.
. En el día 11 se llevó a cabo la prueba de expresión de la sensibilización locomotora. En ese día se administró el etanol y alguno o algunos de los compuestos a evaluar, dependiendo del experimento. Et: etanol; H: habituación; LB: línea base de la actividad locomotora; S: salina.")
Diagrama esquemático que ilustra la línea del tiempo del procedimiento de la sensibilización locomotora inducida por etanol (0.25mg/kg). En el día 11 se llevó a cabo la prueba de expresión de la sensibilización locomotora. En ese día se administró el etanol y alguno o algunos de los compuestos a evaluar, dependiendo del experimento.
Et: etanol; H: habituación; LB: línea base de la actividad locomotora; S: salina.
El primer día, las ratas fueron introducidas en las cajas de actividad durante 60min (sesión de habituación). En el segundo día, las ratas recibieron una inyección ip de 1.0ml/kg de salina; después fueron introducidas en las cajas de actividad motora durante 60min para obtener el registro de la actividad motora (línea base). Del día 3 al 7 las ratas recibieron la administración de una inyección ip de 0.25mg/kg de etanol y posteriormente fueron introducidas en las cajas de actividad durante 60min para obtener el registro de la actividad motora (fase de desarrollo de la sensibilización locomotora). En el día 10 las ratas recibieron la administración de una inyección ip de 1.0ml/kg de salina y fueron introducidas en las cajas de actividad motora para contabilizar su actividad durante 60min, con el fin de verificar si existía actividad condicionada significativa (prueba de salina). En el día 11, las ratas recibieron la administración de una inyección ip de etanol y alguno o algunos de los compuestos a evaluar, dependiendo del experimento (prueba de la expresión de la sensibilización locomotora). Posteriormente, fueron introducidas en las cajas de actividad durante 60min para obtener el registro de la actividad motora (fig. 1).
Efectos del agonista 5-HT2C Ro 60-0175 en la expresión de la sensibilización locomotora inducida por etanol. Durante los días 3-7 del experimento, los animales recibieron una inyección ip de etanol (0.25mg/kg). Los días 8-9, los animales permanecieron en sus cajas-hogar libres de cualquier tratamiento. En el día 10 se llevó a cabo la prueba con salina (1.0ml/kg), y el día 11, los animales recibieron diferentes inyecciones ip de acuerdo con el grupo al que pertenecían: salina (1.0ml/kg; -2min)+salina (1.0ml/kg; -1min), salina (1.0ml/kg; -2min)+etanol (0.25mg/kg; -1min), Ro 60-0175 (0.3mg/kg; -2min)+etanol (0.25mg/kg; -1min), Ro 60-0175 (1.0mg/kg; -2min)+etanol (0.25mg/kg; -1min), Ro 60-0175 (3.0mg/kg; -2min)+etanol (0.25mg/kg; -1min). Cada combinación de los compuestos representa un grupo de 10 ratas. En todas las sesiones se midió la actividad locomotora durante 60min.
Efectos del antagonista 5-HT2C SB 242084 en la expresión de la sensibilización locomotora inducida por etanol. Durante los días 3-7 del experimento, los animales recibieron una inyección ip de etanol (0.25mg/kg). Los días 8-9, los animales permanecieron libres de etanol en sus cajas-hogar. En el día 10, se hizo la prueba con salina (1.0ml/kg), y el día 11, los animales recibieron diferentes inyecciones ip de acuerdo con el grupo al que pertenecían: salina (1.0ml/kg; -10min)+salina (1.0ml/kg; -1min), salina (1.0ml/kg; -10min)+etanol (0.25mg/kg; -1min), SB 242084 (0.1mg/kg; -10min)+etanol (0.25mg/kg; -1min), SB 242084 (0.3mg/kg; -10min)+etanol (0.25mg/kg; -1min), SB 242084 (1.0mg/kg; -10min)+etanol (0.25mg/kg; -1min). Cada combinación de los compuestos representa un grupo de 10 ratas. En todas las sesiones se midió la actividad locomotora durante 60min.
Efectos del antagonista 5-HT2C SB 242084 sobre los efectos de agonista 5-HT2C Ro 60-0175 en la expresión de la sensibilización locomotora inducida por etanol. Durante los días 3-7 del experimento, los animales recibieron una inyección ip de etanol (0.25mg/kg). Los días 8-9, los animales permanecieron libres de etanol en sus cajas-hogar. En el día 10, se hizo la prueba con salina (1.0ml/kg), y el día 11, los animales recibieron diferentes inyecciones ip de acuerdo con el grupo al que pertenecían: salina (1.0ml/kg; -10min)+salina (1.0ml/kg; -2min)+salina (1.0ml/kg; -1min), salina (1.0ml/kg; -10min)+salina (1.0ml/kg; -2min)+etanol (0.25mg/kg; -1min), SB 242084 (0.1mg/kg; -10min)+Ro 60-0175 (3.0mg/kg; -2min)+etanol (0.25mg/kg; -1min), SB 242084 (0.3mg/kg; -10min)+Ro 60-0175 (3.0mg/kg; -2min)+etanol (0.25mg/kg; -1min), SB 242084 (1.0mg/kg; -10min)+Ro 60-0175 (3.0mg/kg; -2min)+etanol (0.25mg/kg; -1min). Cada combinación de los compuestos representa un grupo de 10 ratas. En todas las sesiones se midió la actividad locomotora durante 60min.
En los 3 experimentos previos, los números después de las dosis entre paréntesis indican los tiempos antes de introducir las ratas en las cajas de actividad.
Análisis de los datosPara analizar la sensibilización locomotora inducida por etanol se cuantificó la distancia recorrida en centímetros (media±EEM) en las diferentes fases de la investigación. En la fase del desarrollo de la sensibilización locomotora se utilizó un ANOVA factorial 2×5 de medidas repetidas con efectos principales de días (día 3 vs. día 7; la medida repetida), grupos y la interacción días×grupos. En las fases de prueba o retos farmacológicos, se utilizó un ANOVA de un factor. Los ANOVA aplicados en cada fase fueron con datos independientes de los mismos grupos que corresponden a las diferentes fases del experimento. Para las comparaciones a posteriori de los resultados que mostraron diferencias significativas en los ANOVA se utilizó la prueba de Tukey (p<0.05).
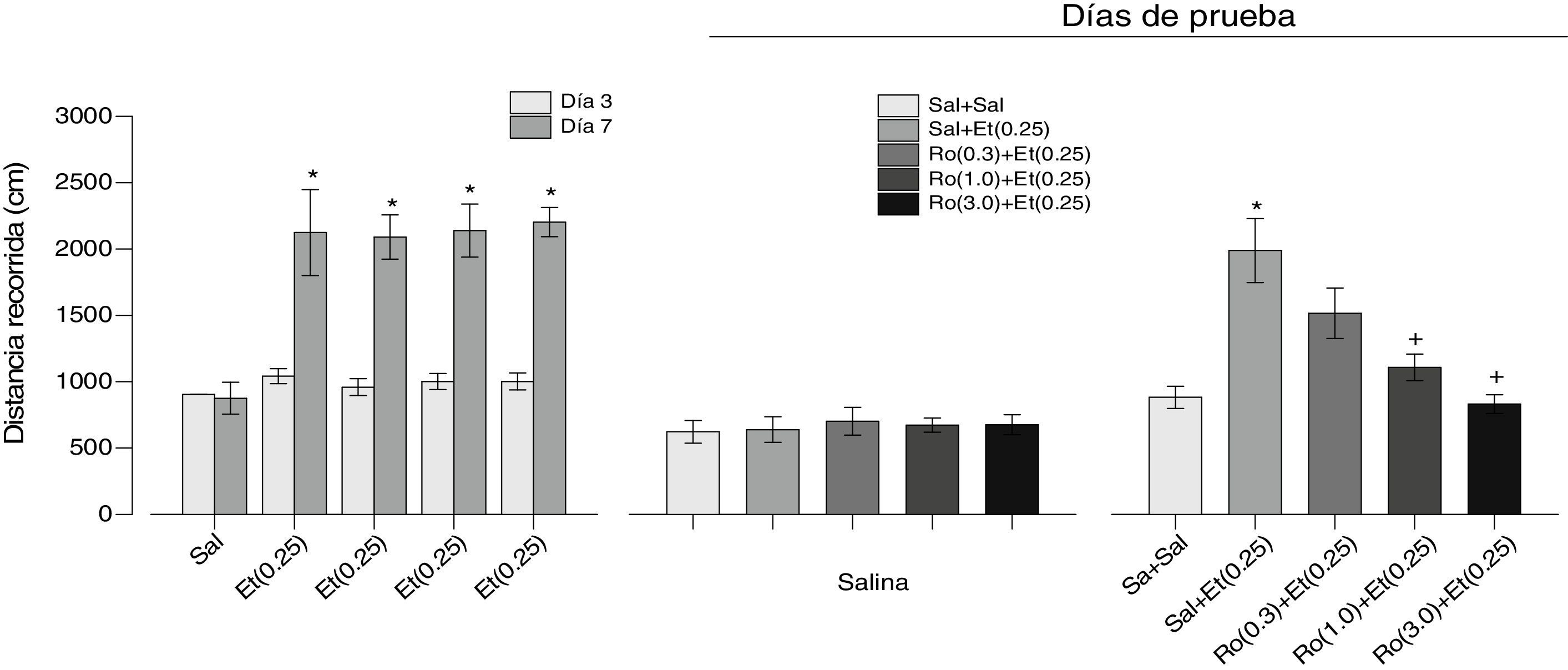
ResultadosEfectos del agonista 5-HT2C Ro 60-0175 en la expresión de la sensibilización locomotora inducida por etanol. Los resultados del tratamiento repetido con etanol se muestran en el panel izquierdo de la figura 2. Como puede observarse, la administración repetida de etanol condujo al desarrollo de la sensibilización locomotora en todos los grupos, excepto en el grupo salina. Un ANOVA factorial de medidas repetidas mostró un efecto significativo de Grupos (F[4.45]=15.464, p=0.0001), Días (F[1.45]=143.298, p=0.0001) y de la interacción Grupos×Días (F[4.45]=9.603, p=0.0001). Las comparaciones posteriores con la prueba de Tukey revelaron diferencias significativas en la distancia recorrida entre el día 7 y el día 3 en todos los grupos tratados con etanol. Los resultados de la prueba con salina (día 10) se muestran en el panel central de la figura 2; como puede observarse, no hay diferencias entre los grupos (F[4.45]=0.138, p=0.967). Los resultados del tratamiento con diferentes dosis del agonista 5-HT2C Ro 60-0175 más etanol (día 11) se muestran en el panel derecho de la figura 2. El Ro 60-0175 redujo la expresión de la sensibilización locomotora provocada por la administración de etanol en función de la dosis del Ro 60-0175 (F[4.45]=10.157, p=0.0001). La prueba de comparaciones posteriores Tukey reveló que la actividad locomotora en los grupos Ro(1.0)+Et(0.25) y Ro(3.0)+Et(0.25) fue significativamente diferente del grupo Sal+Et(0.25). La misma prueba también reveló que la actividad locomotora del grupo Sal+Et(0.25) fue significativamente diferente de la del grupo Sal+Sal.
, que se sometieron primero a una fase de desarrollo de la sensibilización locomotora inducida por etanol (panel izquierdo), después a una prueba de salina (panel central) y, finalmente, a una prueba de la expresión de la sensibilización locomotora inducida por etanol (panel derecho). Entre paréntesis se indica la dosis en mg. Los * indican diferencias significativas con el día 3 y en el panel derecho, con el grupo Sal+Sal. Los + indican diferencias con el grupo Sal+Et(0.25) (Tukey, p<0.05 después de los ANOVA respectivos). Et: etanol; Ro: Ro 60-0175; Sal: salina.")
Se muestran los resultados de los efectos del agonista 5-HT2C Ro 60-0175 sobre la expresión de la sensibilización locomotora inducida por etanol. Las barras representan la media±EEM. Se utilizaron 5 grupos de ratas (n=10), que se sometieron primero a una fase de desarrollo de la sensibilización locomotora inducida por etanol (panel izquierdo), después a una prueba de salina (panel central) y, finalmente, a una prueba de la expresión de la sensibilización locomotora inducida por etanol (panel derecho). Entre paréntesis se indica la dosis en mg. Los * indican diferencias significativas con el día 3 y en el panel derecho, con el grupo Sal+Sal. Los + indican diferencias con el grupo Sal+Et(0.25) (Tukey, p<0.05 después de los ANOVA respectivos).
Et: etanol; Ro: Ro 60-0175; Sal: salina.
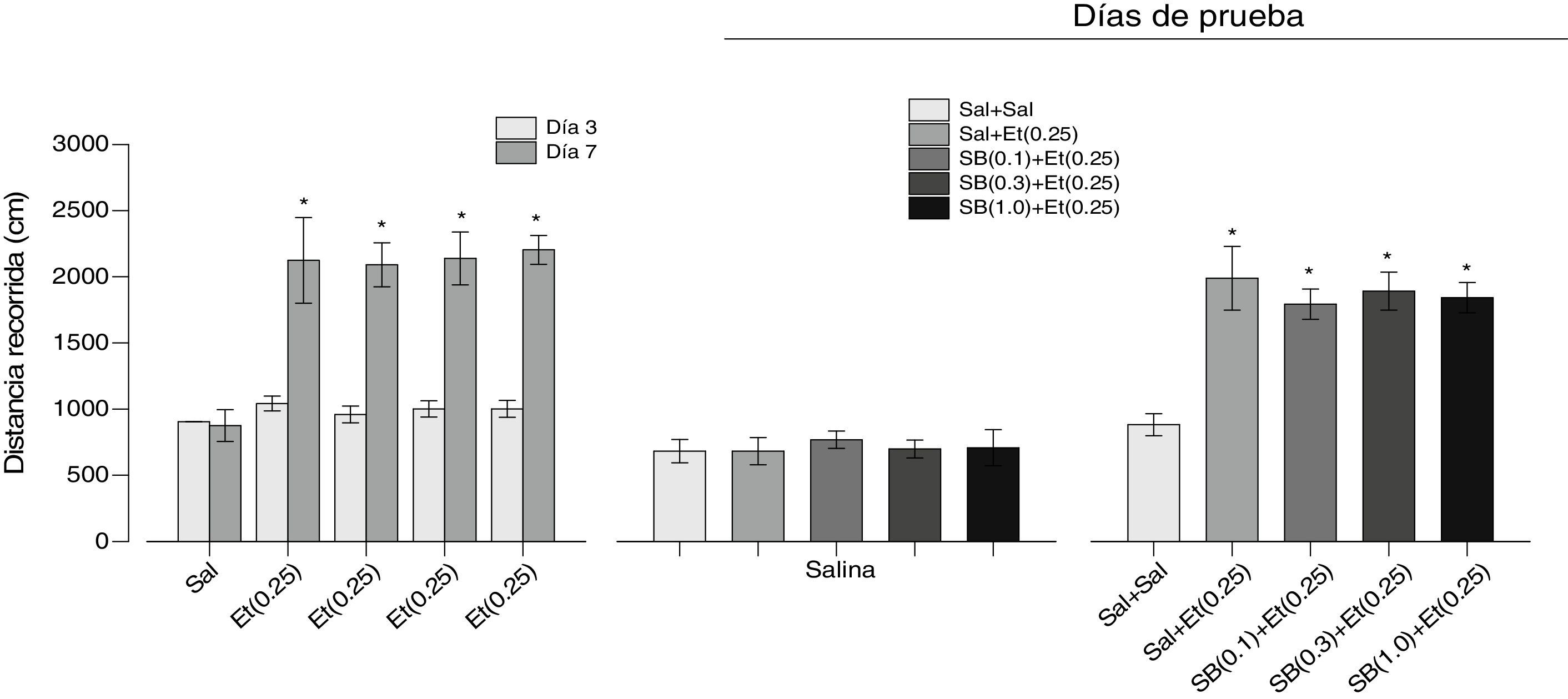
Efectos del antagonista 5-HT2C SB 242084 en la expresión de la sensibilización locomotora inducida por etanol. La administración repetida de etanol produjo el desarrollo de la sensibilización locomotora (fig. 3, panel izquierdo). Un ANOVA factorial de medidas repetidas mostró un efecto significativo de Grupos (F[4.45]=39.938, p=0.0001), Días (F[1.45]=467.812, p=0.0001) y de la interacción Grupos×Días (F[4.45]=31.286, p=0.0001). Las comparaciones posteriores con la prueba de Tukey revelaron diferencias significativas en la distancia recorrida entre el día 7 y el día 3 en todos los grupos tratados con etanol. Los resultados de la prueba con salina (día 10) se muestran en el panel central de la figura 3; como puede observarse, no hay diferencias entre los grupos (F[4.45]=0.141, p=0.966). Los resultados del tratamiento con diferentes dosis del antagonista 5-HT2C SB 242084 más etanol (día 11) se muestran en el panel derecho de la figura 3. Como se puede observar, la coadministración del SB 242084 y etanol no alteró la expresión de la sensibilización locomotora provocada por el etanol (F[4.45]=9.086, p=0.001). La prueba de comparaciones posteriores Tukey reveló que la actividad locomotora en los grupos a los cuales se les administró etanol fue significativamente diferente a la del grupo que solo recibió salina. La misma prueba también reveló que no hubo diferencias significativas en la actividad locomotora entre los grupos a los cuales se les administró el antagonista SB 242084 y el etanol.
, que se sometieron primero a una fase de desarrollo de la sensibilización locomotora inducida por etanol (panel izquierdo), después a una prueba de salina (panel central) y, por último, a una prueba de la expresión de la sensibilización locomotora inducida por etanol (panel derecho). Entre paréntesis se indica la dosis en mg. Los * indican diferencias significativas con el día 3 y en el panel derecho, con el grupo Sal+Sal (Tukey, p<0.05 después de los ANOVA respectivos). Et: etanol; SB: SB 242084; Sal: salina.")
Se muestran los resultados de los efectos del antagonista 5-HT2C SB 242084 en la expresión de la sensibilización locomotora inducida por etanol. Las barras representan la media±EEM. Se utilizaron 5 grupos de ratas (n=10), que se sometieron primero a una fase de desarrollo de la sensibilización locomotora inducida por etanol (panel izquierdo), después a una prueba de salina (panel central) y, por último, a una prueba de la expresión de la sensibilización locomotora inducida por etanol (panel derecho). Entre paréntesis se indica la dosis en mg. Los * indican diferencias significativas con el día 3 y en el panel derecho, con el grupo Sal+Sal (Tukey, p<0.05 después de los ANOVA respectivos).
Et: etanol; SB: SB 242084; Sal: salina.
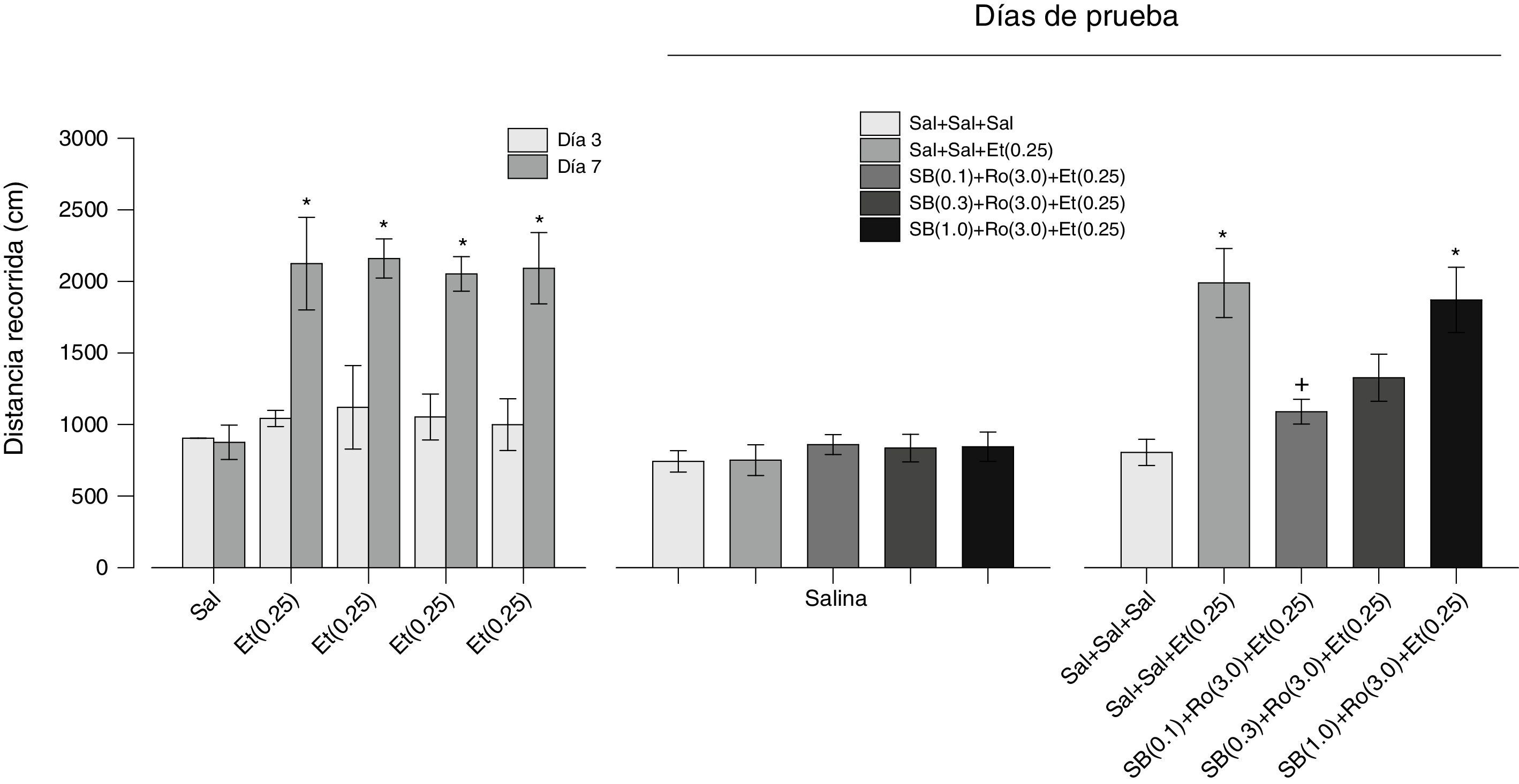
Efectos del antagonista 5-HT2C SB 242084 sobre los efectos del agonista 5-HT2C Ro 60-0175 en la expresión de la sensibilización locomotora inducida por etanol. Los resultados del tratamiento repetido con etanol se muestran en el panel izquierdo de la figura 4. Como puede observarse, la administración repetida de etanol condujo al desarrollo de sensibilización locomotora en todos los grupos, excepto en el grupo de salina. Un ANOVA factorial de medidas repetidas mostró un efecto significativo de Grupos (F[4.45]=12.786, p=0.001), Días (F[1.45]=95.949, p=0.001) y de la interacción Grupos×Días (F[4.45]=6.513, p=0.001). Las comparaciones posteriores con la prueba de Tukey revelaron diferencias significativas en la distancia recorrida entre el día 7 y el día 3 en todos los grupos tratados con etanol. Los resultados de la prueba con salina (día 10) se muestran en el panel central de la figura 4; como puede observarse, no hay diferencias entre los grupos (F[4.45]=0.368, p=0.830). Los resultados del tratamiento con diferentes dosis del antagonista 5-HT2C SB 242084 más el agonista 5-HT2C Ro 60-0175 más etanol (día 11) se muestran en el panel derecho de la figura 4. El SB 242084 previno el efecto del Ro 60-0175 sobre la actividad locomotora provocada por etanol (F[4.45]=9.337, p=0.0001). La prueba de comparaciones posteriores de Tukey reveló que la actividad locomotora en los grupos Sal+Sal+Et(0.25) y SB(1.0)+Ro(3.0)+Et(0.25) fue significativamente diferente a la del grupo Sal+Sal+Sal. La misma prueba también reveló que la actividad locomotora del grupo SB(0.1)+Ro(3.0)+Et(0.25) fue significativamente diferente a la de los grupos Sal+Sal+Et(0.25) y SB(1.0)+Ro(3.0)+Et(0.25).
, que se sometieron primero a una fase de desarrollo de la sensibilización locomotora inducida por etanol (panel izquierdo), después a una prueba de salina (panel central) y, finalmente, a una prueba de la expresión de la sensibilización locomotora inducida por etanol (panel derecho). Entre paréntesis se indica la dosis en mg. Los * indican diferencias significativas con el día 3 y en el panel derecho, con el grupo Sal+Sal+Sal. Los + indican diferencias con el grupo Sal+Sal+Et(0.25) (Tukey, p<0.05 después de los ANOVA respectivos). Et: etanol; Ro: Ro 60-0175; SB: SB 242084; Sal: salina.")
Se muestran los resultados de los efectos del antagonista 5-HT2C SB 242084 sobre los efectos del agonista 5-HT2C Ro 60-0175 en la expresión de la sensibilización locomotora inducida por etanol. Las barras representan la media±EEM. Se utilizaron 5 grupos de ratas (n=10), que se sometieron primero a una fase de desarrollo de la sensibilización locomotora inducida por etanol (panel izquierdo), después a una prueba de salina (panel central) y, finalmente, a una prueba de la expresión de la sensibilización locomotora inducida por etanol (panel derecho). Entre paréntesis se indica la dosis en mg. Los * indican diferencias significativas con el día 3 y en el panel derecho, con el grupo Sal+Sal+Sal. Los + indican diferencias con el grupo Sal+Sal+Et(0.25) (Tukey, p<0.05 después de los ANOVA respectivos).
Et: etanol; Ro: Ro 60-0175; SB: SB 242084; Sal: salina.
El presente estudio examinó la influencia del agonista 5-HT2C Ro 60-0175 en la sensibilización locomotora provocada por etanol. Adicionalmente, también se examinaron los efectos del antagonista selectivo de los receptores 5-HT2C SB 242084 sobre los efectos del Ro 60-0175 en la sensibilización locomotora inducida por etanol, con el fin de determinar si los efectos del Ro 60-0175 resultan de una acción específica sobre los receptores 5-HT2C. Nosotros reportamos que la administración del Ro 60-0175 disminuyó la expresión de la sensibilización locomotora provocada por etanol y que este efecto fue revertido por la administración del antagonista SB 242084. Estos resultados proporcionan evidencia adicional de que la activación de los receptores 5-HT2C regula las conductas relacionadas con la administración de etanol, como la sensibilización locomotora.
Los resultados mostrados anteriormente son consistentes con estudios previos que demuestran que el agonista 5-HT2C Ro 60-0175 juega un papel modulatorio en algunos efectos conductuales relacionados con la administración de etanol. Por ejemplo, se ha reportado que la administración de Ro 60-0175 redujo la autoadministración de etanol en ratas bajo un programa operante de razón fija 4. Este efecto fue revertido por la administración del antagonista selectivo de los receptores 5-HT2C SB 242084 (Tomkins et al., 2002). También se ha reportado que el Ro 60-0175 disminuye el consumo voluntario de etanol en ratas (Kasper et al., 2013). Adicionalmente, se ha reportado que la administración de la lorcaserina, agonista que activa a los receptores 5-HT2C, reduce el consumo de etanol en ratas que prefieren el etanol (Rezvani, Cauley y Levin, 2014).
De manera adicional, se ha reportado que los agonistas 5-HT2C Ro 60-0175 y lorcaserina atenúan las conductas relacionadas con diferentes drogas de abuso. Por ejemplo, se ha reportado que la lorcaserina disminuye la sensibilización locomotora inducida por la administración de heroína o morfina, así como los síntomas de retirada precipitados por naloxona en ratones tratados con heroína (Wu et al., 2015) o morfina (Zhang et al., 2016). Algunos estudios han señalado que el Ro 60-0175 disminuye la conducta impulsiva provocada por anfetamina (Fletcher, Rizos, Noble y Higgins, 2011), la autoadministración de nicotina bajo diferentes programas de reforzamiento (Fletcher et al., 2012) y la hiperlocomoción inducida por la administración de 15mg/kg de cocaína (Cathala et al., 2015).
Los datos previos evidencian que los receptores 5-HT2C juegan un papel importante en las conductas relacionadas con el consumo de etanol y otras drogas adictivas (Higgins y Fletcher, 2015). El mecanismo que podría explicar por qué en la presente investigación se observa una disminución de los efectos locomotores inducidos por la administración de etanol después de la administración del agonista 5-HT2C Ro 60-0175 podría involucrar la activación de los receptores 5-HT2C en las interneuronas GABAérgicas del ATV. Este planteamiento está basado en los datos que se exponen a continuación. Primero, todas las drogas adictivas, incluido el etanol, estimulan directa o indirectamente el sistema de la gratificación del cerebro, el cual en su forma más elemental incluye a las neuronas DAérgicas del ATV que envían proyecciones al nAcc donde liberan el neurotransmisor DA (Koob, 2013). Segundo, el etanol tiene afinidad por el sitio alostérico de la subunidad α de los receptores GABAA localizados en las interneuronas GABAérgicas del ATV (Enoch, 2008). La activación de estos receptores por el etanol produce la inhibición de la actividad de las interneuronas GABAérgicas del ATV (Stobbs et al., 2004); como consecuencia de esta inhibición, las neuronas DAérgicas del ATV incrementan su liberación de DA en el nAcc. Este incremento de DA en nAcc es el responsable de las propiedades adictivas del etanol (Weiss et al., 1993). Tercero, el control inhibitorio de las interneuronas GABAérgicas sobre las neuronas DAérgicas del ATV esta modulado por diversos receptores, entre ellos, los 5-HT2C, que se encuentran localizados en los cuerpos de las interneuronas GABAérgicas del ATV (Bubar y Cunningham, 2007; Bubar, Stutz y Cunningham, 2011). La estimulación de estos receptores con el agonista Ro 60-0175 disminuye, de manera dependiente de la dosis, la tasa de disparo de las neuronas DAérgicas del ATV y la liberación de DA en el nAcc (Di Matteo, di Giovanni, di Mascio y Esposito, 1999); la administración del Ro 60-0175 dentro del ATV disminuye el aumento de DA en el nAcc provocado por la administración de cocaína (Navailles, Moison, Cunningham y Spampinato, 2008). También se ha reportado que la administración sistémica de Ro 60-0175 disminuye la hiperactividad inducida por la administración de cocaína y este efecto fue bloqueado por el pretratamiento con el antagonista 5-HT2C SB 242084 (Grottick, Fletcher y Higgins, 2000). Estos datos, junto con los reportados en el presente estudio, proporcionan evidencia de que los receptores 5-HT2C modulan las conductas relacionadas no solo con las drogas adictivas como los psicoestimulantes y otras, sino también las relacionadas con el etanol, y podrían representar un blanco en el tratamiento de las adicciones a las drogas de abuso, incluyendo el etanol.
En conclusión, la presente investigación mostró que la administración del agonista 5-HT2C Ro 60-0175 disminuyó la expresión de la sensibilización locomotora provocada por etanol, efecto que fue revertido por el pretratamiento con el antagonista selectivo de los receptores 5-HT2C SB 242085. Estos datos, junto con los de otros estudios, proporcionan evidencia del papel modulatorio de los receptores 5-HT2C en las conductas relacionadas con las drogas adictivas, incluyendo el etanol.
FinanciaciónEste estudio fue financiado por PAPITT IN301717 (DGAPA-UNAM).
Conflicto de interesesLos autores declaran no tener ningún conflicto de intereses.
Juan C. Jiménez es estudiante de doctorado del Programa de Maestría y Doctorado en Psicología de la Universidad Nacional Autónoma de México (UNAM) y recibió una beca de CONACyT.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.