


The multi-enzyme complex (crude extract) of white rot fungi Pleurotus ostreatus, Pleurotus eryngii, Trametes versicolor, Pycnosporus sanguineus and Phanerochaete chrysosporium were characterized, evaluated in the hydrolysis of pretreated pulps of sorghum straw and compared efficiency with commercial enzyme. Most fungi complexes had better hydrolysis rates compared with purified commercial enzyme.
Enzymes extracts from different fungi have been a strategy used in saccharification of lignocellulosic biomass, blending (2 or more extracts of different fungi) or only extract. The crude enzyme extracts offers low cost, no activities are lost in concentration/purification processes, a wide spectrum of enzyme activities is maintained and synergy among enzymes.1–6
This study produced, by solid-state fermentation (SSF) of forage sorghum straw, the crude extracts (called multi-enzyme complex) of 5 different fungi. The complexes obtained of each of the fungi were characterized to the apparent activities of cellulases and were evaluated for enzymatic saccharification of own in natura sorghum straw (not pretreated) and sorghum pretreated straw pulps, and the results compared to the hydrolysis carried out by a commercial cellulase complex.
Were evaluated the potential of crude enzymatic extracts of white rot fungi: Pleurotus ostreatus PLO06, Pleurotus eryngii PLE04, Trametes versicolor TRAM01, Pycnosporus sanguineus PYC02 and Phanerochaete chrysosporium PC and, obtained by SSF on straw forage sorghum. Fungi are from the collection of the Department of Microbiology, Federal University of Viçosa, Viçosa, Minas Gerais – Brazil.
To SSF were used straw forage sorghum BRS 655 (stem and leaves without the panicle with the grain) cultivar developed by EMBRAPA Maize and Sorghum and cultivated in the city of Sete Lagoas. Sorghum was cut with 120 days of planting and sun dried, ground and stored in a dry place away from light and moisture.
The sorghum straw was moistened for final humidity of 70%. 100g of the prepared substrate were placed in polypropylene filter bags and sealed with adhesive tape for subsequent autoclaving at 121°C for 60min. After cooled to room temperature, each bag with substrate received two discs of mycelia, with a diameter of 2 inches, of each fungus previously cultured in Petri dish of BDA for 7 days at 30°C. The bags were inoculated in a laminar flow cabinet and incubated in a BOD at 28°C until complete colonization of the substrate. Monitoring of the mycelial growth was carried out visually. After the total substrate colonization by fungi, 20 days after inoculation were obtained the crude extracts (or multi-enzyme complex).
To obtain the enzyme extract, 5g samples of substrate were placed in 250mL Erlenmeyer flasks containing 50mL of sodium citrate buffer (50mM pH 4.8) and shaken at 150rpm for 2h at 5°C. Then filtered through a sieve and placed in 2mL Eppendorf tubes following centrifugation at 12,000×g at 5°C. The supernatant was transferred to another Eppendorf tube, and subsequently identified with the fungus and incubation time, sealed and stored at −18°C.
The reagents used in this study were purchased from Sigma Chemicals companies or Vetec chemistry with analytical grade.
Total cellulase activity or Filter paper activity (FPase) was determined essentially according to the IUPAC7 instructions, and the liberated reducing sugars were estimated by the DNS method.8 FPase activity corresponds to 1μM of reducing sugars as glucose equivalents liberated per min under the assay conditions.
Endoglucanase activity (carboxymethylcellulase, EC 3.2.1.4) or carboxymethycellulase (CMCase) activity was estimated by adding 250μL of the enzyme complex in 1mL of 1% solution of carboxymethylcellulose in 0.05M citrate buffer, pH 4.8 and incubated at 50°C for 30min. One CMCase unit is the amount of enzyme necessary to produce 1μM reducing sugar as glucose equivalents per min under the standard assay conditions.
Exoglucanase activity (EC 3.2.1.91) or AVICELase consisted of adding 250μL of crude enzyme complex in 1mL of 1% solution of microcrystalline cellulose (Avicel) in 0.05M citrate buffer, pH 4.8 and incubated at 50°C for 30min. Periodically, the enzyme-substrate system was stirred in order to maintain the pulp in suspension. One AVICELase unit is the amount of enzyme necessary to produce 1μM reducing sugar as glucose equivalents per min under the standard assay conditions.
Xylanase activity (endo-1,4-β-xylanase, EC 3.2.1.8) was determined in the mixture of 1mL of the enzyme complex, 1mL of xylan solution (1% xylan birchhood – SIGMA) in citrate buffer 0, 05M, pH 4.8 and incubated at 50°C for 30min. Periodically, the enzyme-substrate system was stirred in order to maintain xylan suspension. One Xylanase unit is the amount of enzyme necessary to produce 1μM reducing sugar as glucose equivalents per min under the standard assay conditions. The liberated reducing sugars were estimated by the DNS method.7
The β-glucosidase (EC 3.2.1.21) activity was determined by incubating 1mL of p-nitrophenyl-β-d-glucopyranoside (PNPG) substrate 0.005M, 0.05M citrate buffer, pH 4.8 with 100μL of the enzyme complex (crude extract), for 15min at 50°C. The reaction was stopped by adding 2.0mL of 1.0M sodium bicarbonate and absorbance was measured at 410nm. The unit of β-glucosidase activity was defined as the amount of enzyme capable of releasing 1μmol of p-nitrophenol per minute under the test conditions.
Laccase activity was determined by the oxidation of 2,2′-azino-bis (3-ethylbenzthiazoline-6-sulfonic acid) (ABTS; SIGMA, St. Louis, USA) at 37°C according to Buswell et al.9 The reaction mixture (1mL) contained 600μL enzyme extract, 300μL sodium acetate buffer pH 5.0 (0.1M) and 100μL ABTS solution (1mM). Oxidation was followed via the increase in absorbance at 420nm (γ 420=36.000M−1cm−1). One unit of enzyme activity was defined as the amount of enzyme oxidizing 1μmol of ABTS per minute.
Quantification of protein used the method of Bradford.10
The apparent activity of enzymes FPase (Total Cellulase) AVICELase, CMAase and β-glucosidase were characterized as to pH and temperature, as well as the thermostability of maximal enzyme activity in the temperature through tests to determine activity of those enzyme with varying pH of the buffer or the reaction temperature.
The pH ranged from 3 to 8. The buffer systems used were 50mM citrate buffer (pH 3.0–6.0) and 50mM phosphate buffer (pH 6.0–8.0). The incubation temperature for the measurement of enzyme activity ranged from 30 to 80°C. To estimate the thermostability, the enzymatic complex (samples) was stored in the apparent optimum temperature (results of temperature characterization) of each enzyme in the extract crude, and then made to measure the residual activity of the enzyme 12 in 12h for 48h.
The saccharification experiments were conducted in Erlemeyers 125mL in shaker (Tecnal – TE-421) stirred at 120rpm at 50°C. 0.5mL samples were collected every 6h to 24h, and after 12 12-h to 72h, and each sample was heated at 100°C for 5min to inactivate the enzymes, centrifuged and subsequently was determined the concentration of reducing sugars and glucose. Saccharification was performed in natura sorghum straw (not pre-treated) and pulps of the forage sorghum obtained pretreatments as Cardoso et al.11 For comparison with the commercial enzyme (enzymatic complex Genencor Multifect GC) were conducted saccharification of the pulp obtained pretreatment acid/delignified of the forage sorghum as Cardoso et al.11
LAP 008 protocol used as enzyme saccharification method by NREL,12 suggests an enzyme load in the order of 25FPUg−1 (25 FPase per gram of cellulose). However in this work, because it was not utilized extracts purified, that is more concentrated in relation U/mL, there were changes in load enzymes to about 8FPUg−1 biomass (dry basis) for each fungus and the commercial enzyme. The load of substrate was 0.3g of material lignocellulosic for 1% w/w (dry biomass) in each flask (straw of forage sorghum in natura, pulp acid, pulp delignified and pulp acid/delignified, all the pulps obtained by pretreatment of forage sorghum)11 for each saccharification experiments.
Values are expressed as means±S.D. (Standard Deviation). Comparing the percentages of enzymatic saccharification acid pulp/delignified by enzyme extracts of fungus and the commercial enzyme it was used.
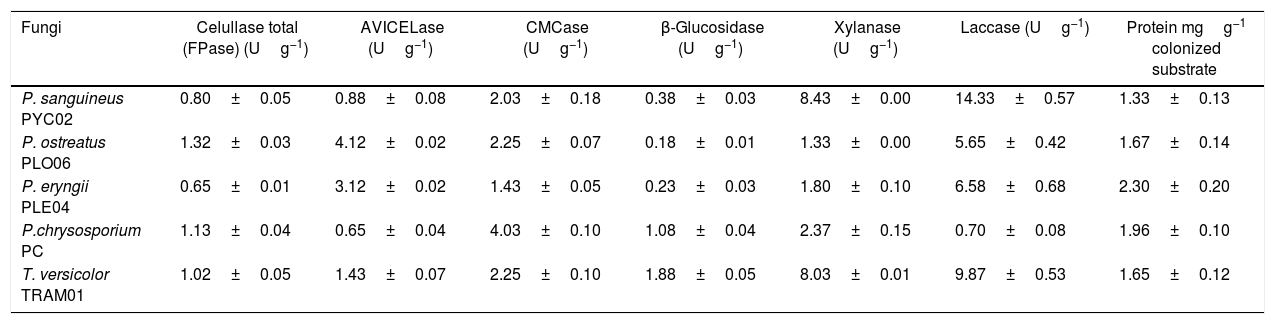
The apparent total cellulase activities (FPase), CMCase, AVICELase and β-glucosidase, xylanase and laccase in the crude extract of white rot fungi, after 20 days of fermentation of sorghum straw with 70% humidity at a temperature of 28°C are shown in Table 1.
Apparent enzymatic activities (Ug−1dm) of the extracts of sorghum straw, fermented by white rot fungi, after 20 days, with 70% humidity and incubated at 28°C. U is the amount of enzyme required to generate μmol products per minute.
| Fungi | Celullase total (FPase) (Ug−1) | AVICELase (Ug−1) | CMCase (Ug−1) | β-Glucosidase (Ug−1) | Xylanase (Ug−1) | Laccase (Ug−1) | Protein mgg−1 colonized substrate |
|---|---|---|---|---|---|---|---|
| P. sanguineus PYC02 | 0.80±0.05 | 0.88±0.08 | 2.03±0.18 | 0.38±0.03 | 8.43±0.00 | 14.33±0.57 | 1.33±0.13 |
| P. ostreatus PLO06 | 1.32±0.03 | 4.12±0.02 | 2.25±0.07 | 0.18±0.01 | 1.33±0.00 | 5.65±0.42 | 1.67±0.14 |
| P. eryngii PLE04 | 0.65±0.01 | 3.12±0.02 | 1.43±0.05 | 0.23±0.03 | 1.80±0.10 | 6.58±0.68 | 2.30±0.20 |
| P.chrysosporium PC | 1.13±0.04 | 0.65±0.04 | 4.03±0.10 | 1.08±0.04 | 2.37±0.15 | 0.70±0.08 | 1.96±0.10 |
| T. versicolor TRAM01 | 1.02±0.05 | 1.43±0.07 | 2.25±0.10 | 1.88±0.05 | 8.03±0.01 | 9.87±0.53 | 1.65±0.12 |
The solid state fermentation of sorghum straw was capable of generating complex multi cellulases, xylanase and laccase, with variations of values for each fungus.
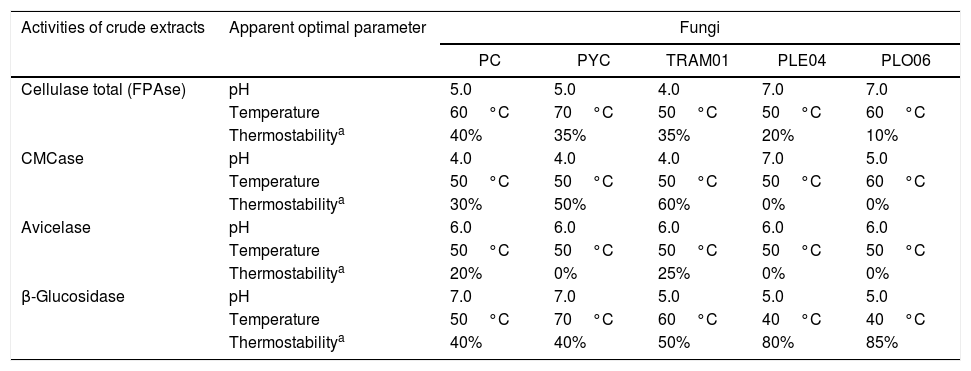
The characterization of the apparent Cellulase Total Activity, CMCase, AVICELase and β-glucosidase from white rot fungi, in relation to pH, temperature and thermostability are shown in Table 2.
The characterization of the apparent total cellulase activity, CMCase, AVICELase and β-glucosidase of white rot fungi in forage sorghum straw SSF. aThe apparent thermostability was evaluated at the apparent optimum temperature of each enzyme.
| Activities of crude extracts | Apparent optimal parameter | Fungi | ||||
|---|---|---|---|---|---|---|
| PC | PYC | TRAM01 | PLE04 | PLO06 | ||
| Cellulase total (FPAse) | pH | 5.0 | 5.0 | 4.0 | 7.0 | 7.0 |
| Temperature | 60°C | 70°C | 50°C | 50°C | 60°C | |
| Thermostabilitya | 40% | 35% | 35% | 20% | 10% | |
| CMCase | pH | 4.0 | 4.0 | 4.0 | 7.0 | 5.0 |
| Temperature | 50°C | 50°C | 50°C | 50°C | 60°C | |
| Thermostabilitya | 30% | 50% | 60% | 0% | 0% | |
| Avicelase | pH | 6.0 | 6.0 | 6.0 | 6.0 | 6.0 |
| Temperature | 50°C | 50°C | 50°C | 50°C | 50°C | |
| Thermostabilitya | 20% | 0% | 25% | 0% | 0% | |
| β-Glucosidase | pH | 7.0 | 7.0 | 5.0 | 5.0 | 5.0 |
| Temperature | 50°C | 70°C | 60°C | 40°C | 40°C | |
| Thermostabilitya | 40% | 40% | 50% | 80% | 85% | |
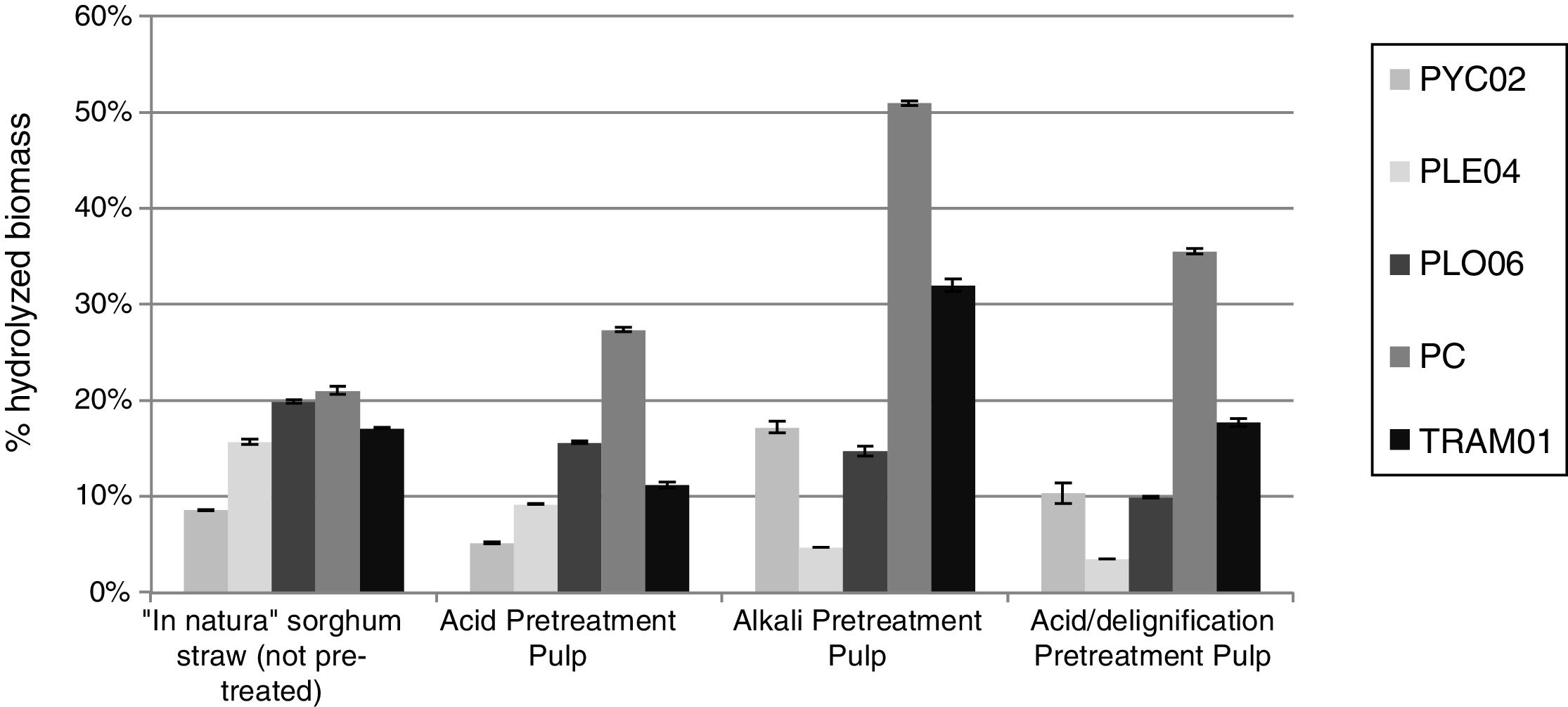
Fig. 1 shows the percentage of hydrolysis of the sorghum in natura and pretreat pulps by multi-enzyme complexes (crude extracts) of white rot fungi, generated by SSF of the sorghum straw.
 and of the pulps obtained from pretreatment according to Cardoso et al.,11 by crude extracts of fungi, P. chrysosporium PC, P. sanguineus PYC02, T. versicolor TRAM01, P. eryngii PLE04 e P. ostreatus PLO06, produced by SSF of forage sorghum straw.")
Saccharification of forage sorghum straw “in natura” (without pre-treatment) and of the pulps obtained from pretreatment according to Cardoso et al.,11 by crude extracts of fungi, P. chrysosporium PC, P. sanguineus PYC02, T. versicolor TRAM01, P. eryngii PLE04 e P. ostreatus PLO06, produced by SSF of forage sorghum straw.
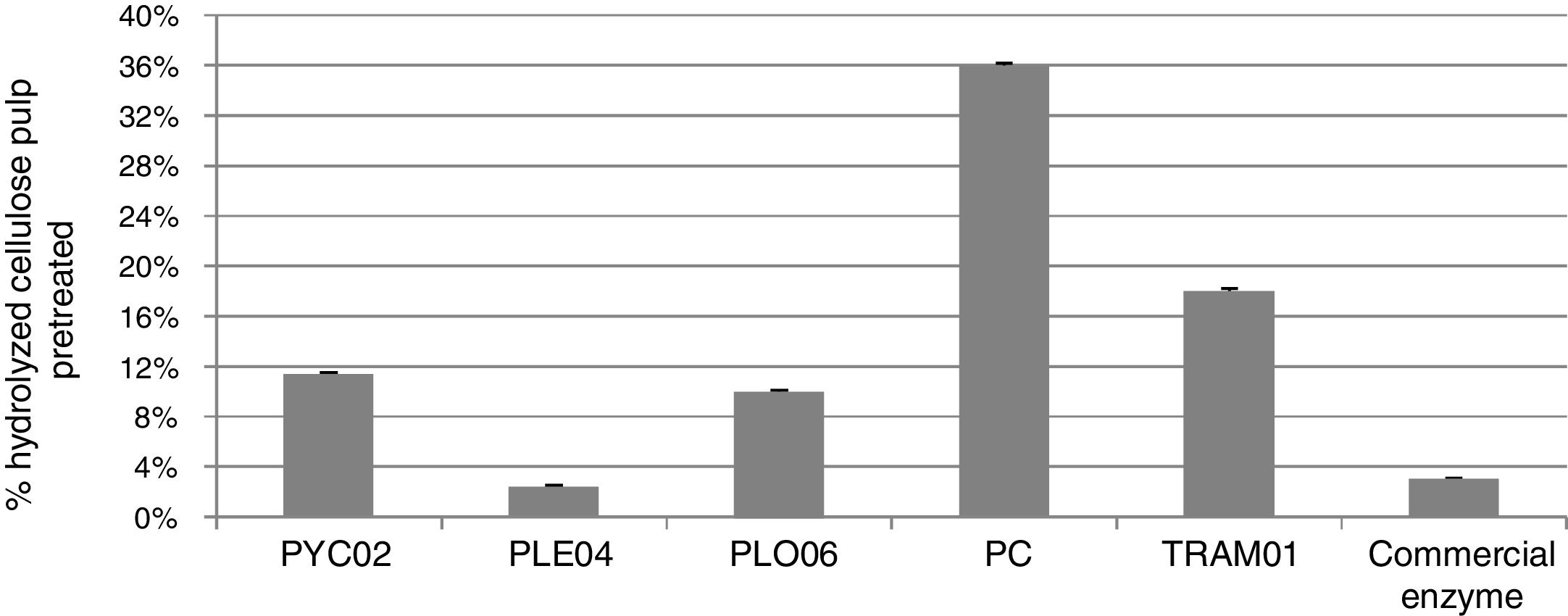
To compare the multi-enzyme complexes of the fungi with a complex of commercial enzymes was performed hydrolysis of acid/delignified pulp (higher percentage of cellulose, less than 1% of lignin and hemicellulose).11 For commercial enzyme were kept the same hydrolysis conditions applied to the extracts of fungi, the commercial enzyme (about 273 FPU/mL) was diluted 130-fold to load of enzymes of the 8.66FPUg−1 of cellulose e and 8.40FPUg−1 of biomass for pretreatment acid/delignified pulp (altered protocol LAP 00812). Fig. 2 shows the results of saccharification of the acid/delignified pulp by multi-enzyme complex of five fungi and commercial enzyme.
.")
Saccharification of the pulp obtained from straw forage sorghum pretreated by acid and delignification, by fungus of multi-enzyme complexes, P. chrysosporium PC, P. sanguineus PYC02, T. versicolor TRAM01, P. eryngii PLE04 and P. ostreatus PLO06, produced by SSF, and commercial enzyme (Multifect GC).
The present study was not carried out any enrichment or pretreatment of the substrate, only sterilization, control of the incubation temperature (28°C) and humidity (70%). The low yield in cellulases may be associated with the insoluble substrate (straw) and low nitrogen levels in the culture medium. Other authors have reported FPase values well above obtained in this work. In research conducted by Elisashvili and Kachlishvili,13 the fungus P. ostreatus reached values near 12.0Ug−1 for FPase and CMCase activity between 6.25 and 325Ug−1, in wheat straw fermentation enriched with yeast extract.
The production xylanase by solid-state fermentation of sorghum straw was the most significant among the hydrolytic enzymes.
In apparent activity of laccase, the highlight was again the P. sanguineus PYC02 with the value of 14.33Ug−1(dm). There is an increasing trend in employment of laccase in biotechnological processes,14,15 and P. sanguineus has been used in Kraft bleaching effluent16 and degradation different dyes.17 However, the value obtained by P. sanguineus PYC02 was lower than that reported by other strains of P. sanguineus, as in the work of Vikineswary et al.,18 where the production of laccase in SSF organic residues reached 48.7Ug−1 (dm).
The extracellular protein values of fungi tested in this study (Table 1) were lower than those reported in other studies,19,20 possibly due to lack of enrichment with nitrogen and also the fact that the biomass used is low protein level.
Results of the thermostability of β-glucosidases, the one that deserves to be highlighted was of the P. sanguineus PYC02 with optimum temperature of 70°C and thermal stability (70°C) for 48h with residual activity about 30%. Thermostable cellulases are considered ideal for biotechnological applications.21 Another β-glucosidase, of the T. versicolor TRAM01, incubated at 60°C remained over 50% activity after 48h.
The alkali pulp reached values close to 50% of saccharified biomass, followed by acid/delignified pulp with the percentage of 36.2% after 72h of saccharification (Fig. 1). Because these materials more easily digestible due to pretreatments, this was expected. It was also expected higher proportion of hydrolysis of the acid/delignified pulp by containing lower content of lignin and hemicellulose and higher cellulose.
Siqueira20 reported that in untreated bagasse hydrolysis by crude extract of Aspergillus awamori achieved is about 30% degradation after 24h of incubation, and about 60% after 96h, a result above obtained in this work, which reached a maximum of 20% after 72h.
The fungi Pleurotus PLE04 and PLO06 highlighted by the lower yield in the saccharification with increased cellulose content of the pretreated materials. Such fact can be linked to β-glucosidase enzyme load of fungi PLE04 and PLO06 (Table 1), which are smaller compared to other fungi. The final accumulation of cellobiose or glucose will inhibit the cellulose hydrolysis reactions, and cellobiose is a more effective inhibitor than glucose.22,23 No hydrolysis of cellobiose due to the low level of β-glucosidase can reach low levels of saccharification.
The enzymatic complex of the P. chrysosporium PC showed the highest results in the hydrolysis of the straw in natura and of the three pulps. Highlighted for alkali pulp with 50% of the biomass hydrolyzed. In Mayrink24 the extract of Trichoderma spp. C012 was obtained in about 50% hydrolysis of the AVICEL (5FPUg−1) at just over 10h of reaction. A similar result to the PC, although the load of the P. chrysosporium PC was slightly higher (8FPUg−1).
As can be observed in Fig. 2, most of the fungi obtained significantly better results than the commercial enzyme that only reached 3%, in relation a cellulose material. It must be remembered that this complex of commercial enzymes (only cellulases) was diluted 130 times. Under normal conditions the same commercial enzymes (load 50FPUg−1 biomass) achieved 95% yield in the hydrolysis of acid/delignified pulp of sorghum straw11. Falkoski et al.25 compared the extract produced by fungus Chrysoporthe cubensis with a commercial enzyme product and found that the produced extract was more efficient for hydrolyzing alkali pretreated sugarcane bagasse per FPU of enzyme applied.
At the end is possible to evaluate that the use of enzymatic complexes obtained from these fungi have the potential to saccharification, and the concentration of these extracts can further increase yields.
FundingThis study was funded with financial aid by Fundação de Amparo a Pesquisa do Estado de Minas Gerais (FAPEMIG) and schorlarship by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Conflict of interestAuthors declare that have no conflict of interest.
The authors acknowledge IFES, FAPEMIG and CNPq for financial support.