


The search for new biocatalysts has aroused great interest due to the variety of micro-organisms and their role as enzyme producers. Native lipases from Aspergillus niger and Rhizopus javanicus were used to enrich the n-3 long-chain polyunsaturated fatty acids content in the triacylglycerols of soybean oil by acidolysis with free fatty acids from sardine oil in solvent-free media. For the immobilization process, the best lipase/support ratios were 1:3 (w/w) for Aspergillus niger lipase and 1:5 (w/w) for Rhizopus javanicus lipase using Amberlite MB-1. Both lipases maintained constant activity for 6 months at 4°C. Reaction time, sardine-free fatty acids:soybean oil mole ratio and initial water content of the lipase were investigated to determine their effects on n-3 long-chain polyunsaturated fatty acids incorporation into soybean oil. Structured triacylglycerols with 11.7 and 7.2% of eicosapentaenoic acid+docosahexaenoic acid were obtained using Aspergillus niger lipase and Rhizopus javanicus lipase, decreasing the n-6/n-3 fatty acids ratio of soybean oil (11:1 to 3.5:1 and 4.7:1, respectively). The best reaction conditions were: initial water content of lipase of 0.86% (w/w), sardine-free faty acids:soybean oil mole ratio of 3:1 and reaction time of 36h, at 40°C. The significant factors for the acidolysis reaction were the sardine-free fatty acids:soybean oil mole ratio and reaction time. The characterization of structured triacylglycerols was obtained using easy ambient sonic-spray ionization mass spectrometry. The enzymatic reaction led to the formation of many structured triacylglycerols containing eicosapentaenoic acid, docosahexaenoic acid or both polyunsaturated fatty acids.
Lipases (acylglycerolacylhydrolases, EC 3.1.1.3) are biocatalysts with many industrial applications, naturally acting on carboxylic ester bonds to catalyze the hydrolysis of triacylglycerols (TAGs). In non-aqueous media, they can catalyze esterification, alcoholysis, acidolysis and transesterification reactions. The screening, kinetic characterization and immobilization of microbial lipases for use in biocatalysis has been an important focus of research in our laboratory. Previously we have described the selection by screening techniques on solid and liquid media of lipase-producing microorganisms and the characterization of the catalytic performance of fungal lipase preparations for hydrolysis and esterification reactions.1–4 Among these strains, Aspergillus niger, Rhizopus javanicus, Fusarium oxysporum and Penicillium solitum are described as lipase-producers for the first time.3,5 The chemo-, regio- and enantiospecific behavior of these enzymes are the great industrial interest. Most microbial lipases such as those from A. niger, Rhizopus delemar, Rhizopus miehei, Mucor javanicus, R. javanicus and Yerrowia lipolytica show 1,3-positional specificity releasing 2-monoacylglycerol and 1,2- and 2,3-diacylglycerol as products from the substrate.2 Lipase from A. niger gave the best yields and enantioselectivities in the esterification of racemin ibuprofen.1,6
Lipases can be employed in the modification of the chemical structure of natural TAGs to improve their physicochemical properties and/or their health benefits. For instance, long chain polyunsaturated fatty acids such as the n-3 family (n-3 LCPUFAs) may be incorporated into TAGs from oils and fats via lipase-catalyzed reactions. These n-3 LCPUFAs, especially eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), may promote specific effects related to metabolism and health and they show great promise for nutraceutical applications.
The concept of structured triacylglycerols (STAGs) implies modification of the fatty acids composition and/or fatty acids location on the glycerol backbone, and improvement of the physical and/or physiological properties of dietary lipids. Studies have also shown that enzymatically or chemically modified TAGs exhibit similar or, in some cases, greater benefits compared to blended oils with similar fatty acid compositions.7–11
Soybean oil (SO) is a cheap and abundant commodity in Brazil, representing an important source of fatty acids for regular consumption. SO presents an n-6/n-3 fatty acids ratio ranging between 7.1:1 and 11:1.12,13 Based on experimental and clinical studies, the most favorable n-6/n-3 ratio for human nutrition is proposed to range between 2:1 and 4:1.14
In a previous study, we reported that TAGs from SO enriched in EPA and DHA can be obtained by acidolysis catalyzed by a commercial R. miehei lipase immobilized on a macroporous anion-exchange resin (Lipozyme RM IM®).13 We also reported that some lipases, derived from native microorganisms, provide good results for obtaining PUFA concentrates as acylglycerols by hydrolysis of borage oil, salmon oil and sardine oil.3,5,15 There are some studies reporting the production of TAGs enriched with n-3 PUFA by enzymatic acidolysis.16 However, most of them have used commercially-available lipases in reaction media containing organic solvents and few studies exploring native lipases have been conducted. This fact highlights the importance of exploiting the capacities of novel microbial lipases to increase the content of n-3 LCPUFAs in vegetable oils.
Immobilization of the native lipases is an important part of the process insofar as the immobilized enzymes can be easily recovered and reused thereby making the process economically viable. On account of the relatively high surface hydrophobicity of lipases, the process of simple adsorption onto suitably hydrophobic supports has been the more used strategy for lipase immobilization. Therefore, this work has three main objectives: (i) to immobilize the native lipases from A. niger (AN) and R. javanicus (RJ) on Amberlite MB-1 using different lipase/support ratios; (ii) to determine the activity, stability and degree of incorporation of n-3 LCPUFAs by acidolysis of SO with a mixture of free fatty acids from sardine oil (sardine-FFA) in solvent-free media, by free and immobilized lipases; and (iii) to investigate some of the variables which may influence the incorporation of n-3 LCPUFA into SO by enzymatic acidolysis.
Materials and methodsChemicalsAmberlite MB-1 and chemical and culture media reagents were obtained from Merck & Co., Inc. (Darmstadt, Hesse, Germany) and from Sigma–Aldrich Co. (St. Louis, MO, USA). Olive oil (Carbonell®) (Deoleo, Madrid, Spain), soybean oil (Liza®) (Cargill, Inc., São Paulo, São Paulo, Brazil) and sardine oil (Catalent Pharma Solutions, Inc., Sorocaba, São Paulo, Brazil) were purchased in the local market. The preparation of free fatty acids from sardine oil (sardine-FFA) was carried out using alkaline hydrolysis followed by acidification with acetic acid.17 Its major component fatty acids were stearic (5.7%), myristic (7.4%), palmitoleic (8.1%), palmitic (16.5%) and oleic (15.3%) acids, EPA (19.8%) and DHA (11.4%).
Production and determination of lipases activitiesThis study was performed using AN and RJ lipases produced in our laboratory as previously described.6 The production was carried out in a basal medium with an initial pH value of 6.0 that consisted of 2% peptone, 0.5% yeast extract, 0.1% NaNO3, 0.1%, KH2PO4 0.05% MgSO4. 7H2O and 2% of olive oil. Cultures were carried out in Erlenmeyer flasks (500mL) containing 120mL of the growth medium. The cultures were inoculated with 1mL of spores suspension (105–106 spores/mL) and the flasks were agitated on a rotary shaker (130rpm) at 35°C for 72h. After that, the cultures were filtered and cleared filtrates were treated with ammonium sulphate (80% saturation). The precipitates were dialyzed overnight in sodium phosphate buffer, pH 7.0, and lyophilized for use as a crude lipase preparation in powder form. The residual water of the lyophilized lipases was 0.84% (w/w). Hydrolytic activity was determined by triolein, with olive oil as the substrate and employing a standard oleic acid curve. One activity unit (U) was defined as 1μmol of oleic acid released per minute of reaction. The specific activity was expressed as U/mg of total protein. The amount of protein was determined by the method of Lowry et al.18 AN lipase with 7.5 and RJ lipase with 6.8U/mg were used, without further purification.
Immobilization by adsorptionThe crude lipases were immobilized by adsorption onto Amberlite MB-1 support at different lipase/support ratios (w/w). The crude lipase preparation (0.07–0.17g) was dissolved in 5mL of sodium phosphate buffer (50mM, pH 7.0), mixed with 500mg of dried support and shaken at 200rpm for 12h using a Tecnal orbital shaker TE-421 (Tecnal, Ltd., Piracicaba, São Paulo, Brazil). After vacuum filtration, the mixture was washed three times with deionized water and rinsed with sodium phosphate buffer. The lipase preparations were freeze dried and stored at 4°C until further use.
Immobilization yieldImmobilization yield was defined as follows:
where aimm is the total hydrolytic activity of immobilized enzyme (U/mg of protein) and afree is the total hydrolytic activity of initial enzyme preparation (U/mg of protein). The amount of protein bound to the support was calculated by subtracting the amount of protein recovered after washing the Amberlite MB-1-lipase complex from the amount of protein used for immobilization.Acidolysis reaction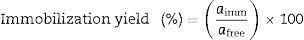
The free and immobilized lipases (10%, w/w) were added to the reaction medium (13g) composed of sardine-FFA and SO in a molar ratio of 3:1, in 50mL conical flasks with silicone-capped stoppers. The reaction was carried out under nitrogen atmosphere and 0.001% BHT was added to the reaction medium to avoid degradation of the PUFAs. The reaction mixture was incubated at 40°C using a Tecnal orbital shaker TE-421, at 160rpm for 12h, and the reaction was stopped by filtering the lipase. The reaction product was flushed out with a stream of nitrogen and stored at −20°C until analyzed. The STAGs were purified and analyzed by GC and EASI-MS as described below. The reactions and analyses were carried out in triplicate.
Experimental designResponse surface methodology (RSM) was adopted as it provides a set of mathematical and statistical methods that make it possible to model phenomena and find combinations of experimental variables that will lead to optimal responses. A 23 full factorial design with 4 center points to determine the residual variance (Table 2) was used in the statistical optimization assays. This process made it possible to determine the best conditions for the acidolysis reaction for each lipase. The independent variables (or factors) studied were: reaction time (hours, X1), sardine-FFA:SO mole ratio (X2) and initial water content of lipase (% w/w, X3). The dependent variable investigated was the n-6/n-3 FAs ratio of the STAGs. The experimental matrix was designed using Statistica 9.0 software (StatSoft, Inc.) (Tulsa, OK, USA), and the significance of data was accessed by the analysis of variance (ANOVA) statistical test.
Purification of the STAGsThe STAGs and FFAs obtained after the acidolysis reaction were separated using a previously described method, with modifications: the reaction mixture was dissolved in n-hexane to 200mL, 150mL of 0.8N KOH (hydro-alcoholic solution, 30% ethanol) were added, and the mixture was agitated.19 The hydro-alcoholic phase (containing FFAs as potassium salts), and the hexane phase (containing the TAGs) were decanted, then the hydro-alcoholic phase was extracted twice more with 20mL of n-hexane. Both n-hexane solutions were mixed together, hexane was evaporated off, the STAGs were weighed and the recovery percentage was calculated. After the purification step, it was possible to recover about 80% of the STAGs. A thin-layer chromatography (TLC) of the hexane extract was performed in order to confirm the efficiency of the process. The presence of acylglycerols and FFAs was identified by TLC on plates of silica-gel (TLC plates, SIL G-25; Aldrich Chemical Co., Milwaukee, WI, USA) activated by heating at 105°C for 20min. The samples were spotted on the plate (dissolved in hexane, 7%, v/v) with authentic standards and then developed in chloroform/acetone/methanol (95:4.5:0.5, v/v/v). Spots of each lipid were visualized by spraying the plate with iodine vapor in a nitrogen stream.
Gas chromatography (GC) and easy ambient sonic-spray ionization mass spectrometry (EASI-MS)The STAG and FAs profiles were obtained by direct mass spectrometry using the easy ambient sonic-spray ionization (EASI-MS) and by gas chromatography (GC) as previously described.13 The FAs from STAGs were converted into FAME (fatty acid methyl ester) by treatment with BF3-methanol.20 The analysis were performed by gas chromatography with a flame ionization detector using a Chrompack Gas Chromatograph (Chrompack, São Paulo, São Paulo, Brazil) and a 50-m fused silica capillary column (WCOT CP-Sil 88, Chrompack, Chromtech, MN, USA). The fatty acids were identified by comparison of the relative retention times with authentic standards from Sigma Chemical Company and expressed as a relative percentage (% total fatty acids). A single-quadrupole LCMS 2010 mass spectrometer (Shimadzu Corp., Kyoto, Japan) with a homemade EASI source was used to obtain the spectra in the positive ion mode. The analysis conditions were: N2 nebulizing gas at 3Lmin−1, surface angle of ca. 30° and methanol flow rate of 20μLmin−1. The samples (2μL) were placed on the paper surface (brown Kraft paper). The mass spectra were accumulated over 60s and scanned in the 50–1200m/z range.
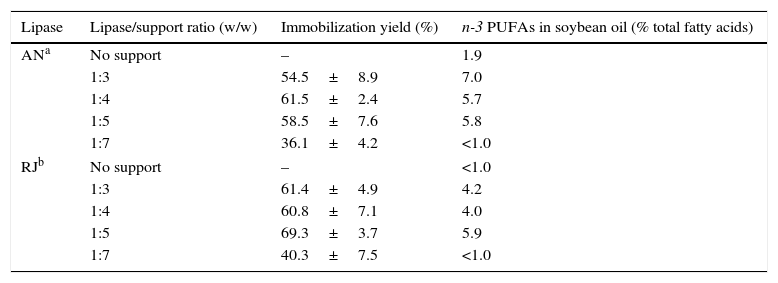
Results and discussionImmobilization of lipases: best lipase/support ratio and stability of enzymesThe capacity of incorporation of n-3 LCPUFAs (EPA+DHA) was dependent on the lipase source and lipase/support ratios for immobilization (Table 1). When the lipases were used in the free form, insignificant incorporation of n-3 LCPUFAs into SO was achieved in 12h of reaction. The total n-3 PUFAs content in the acidolysis products was 4.0–7.0% when immobilized lipases were used, except for a lipase/support ratio of 1:7 (<1.0%) which gave less than 50% of immobilization yield for both lipases. The higher content was 7.0% when immobilized AN was employed (1:3 lipase/support ratio). Larger amounts of enzyme did not further improve the incorporation of n-3 LCPUFAs. This may be due to enzyme aggregation, which leads to an increase in substrate diffusion inhibition, causing the saturation of the reaction rate. These results indicate that immobilized lipases have more access to the substrate molecules than free lipases and, thus, the immobilized system showed higher incorporation of n-3 LCPUFAs into SO. During the immobilization process, multiple binding sites are created as the protein molecules, initially folded and able to contact the surface on only one side, begin to unfold following contact.21
Influence of the lipase/support ratios on the percentage of n-3 PUFAs in soybean oil after acidolysis reactions catalyzed by free and immobilized lipases.
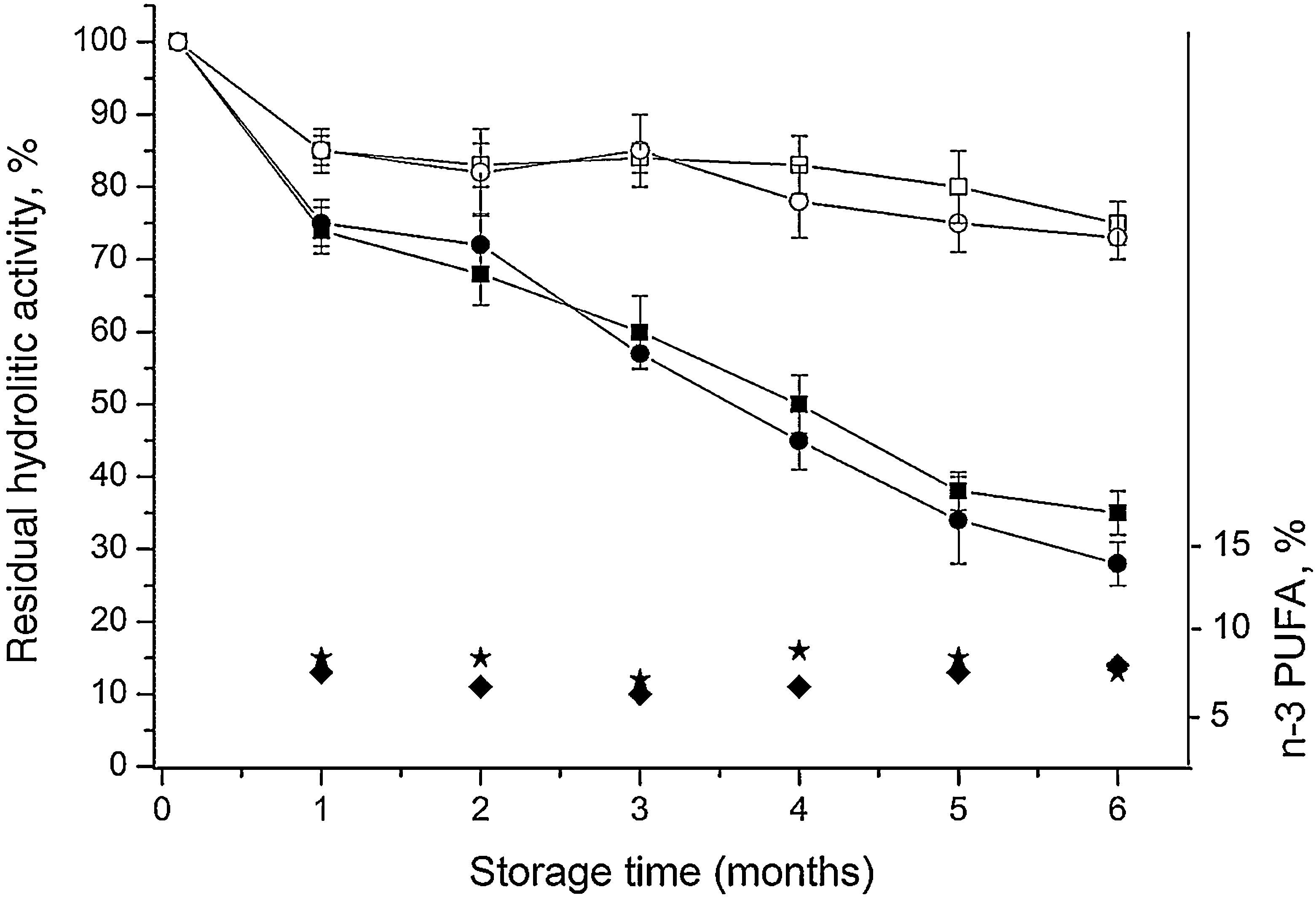
Fig. 1 shows the storage stability of the immobilized lipases compared to that of free lipases. After 6 months of storage at 4°C, the residual hydrolytic activity of immobilized AN and RJ lipases was 75 and 73%, respectively, while that of the free lipases was 35 and 28%, respectively. These data show that the immobilization procedure may increase the storage stability and that proves the significant advantage of Amberlite as an enzyme immobilization resin. Furthermore, the capacity of incorporation of n-3 LCPUFAs into SO was kept constant after 6 months of storage at 4°C. The incorporation of n-3 LCPUFAs into SO showed little variation (5–8%) for either lipase as shown on the right-hand y axis of the graph.
, 166mg of AN lipase or 100mg of RJ lipase, 12h. AN, Aspergillus niger; RJ, Rhizopus javanicus.")
Stability of lipases during 6 months of storage at 4°C. ■ free AN lipase; □ immobilized AN lipase; ● free RJ lipase; ○ immobilized RJ lipase. ★ and ♦ represent the total n-3 PUFAs content on soybean oil after acidolysis using immobilized AN and RJ lipases. Each data point represents the average of three experiments and the error bars show the standard deviation. Operational conditions: 13g of sardine-FFA+soybean oil (3:1), 166mg of AN lipase or 100mg of RJ lipase, 12h. AN, Aspergillus niger; RJ, Rhizopus javanicus.
In addition to increasing storage stability, the immobilization often provides the extra advantage of increasing the catalytic activity of the lipase compared with that of free enzymes. An immobilization procedure for A. niger lipase by covalent binding on chitosan-coated magnetic nanoparticles was recently reported.22 Immobilized derivatives retained above 80% of their initial activity after 15 hydrolytic cycles, and high storage stability during 50 days was observed. Also, lipase from Rhizomucor miehei and M. javanicus immobilized in microemulsion-based organogels maintained good enantioselectivities in the hydrolysis of ketoprofen vinyl ester and their activities increased 12.8-fold and 7.8-fold, respectively, compared with their free forms.23
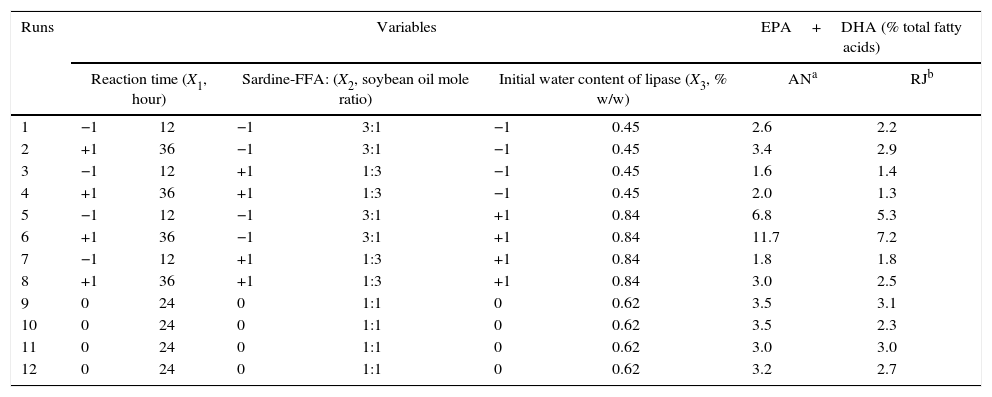
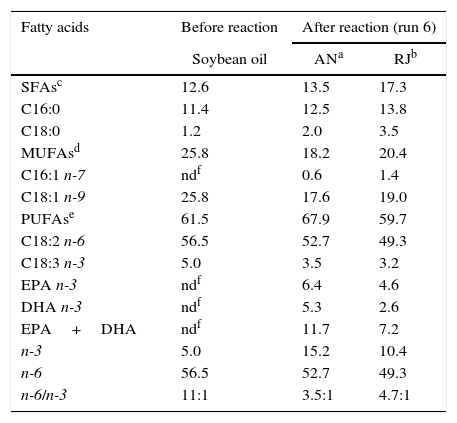
Response surface methodology (RSM)The highest n-3 LCPUFA incorporations (11.7 and 7.2% of EPA+DHA for AN and RJ, respectively) into SO by both lipases were obtained in run 6 (Tables 2 and 3), using a sardine-FFA:SO mole ratio of 3:1, an initial water content of lipase of 0.84% (w/w) and after 36h of reaction. Table 3 shows the FAs composition of SO (before hydrolysis) and of the purified STAGs obtained by acidolysis. The major FAs in SO were linoleic acid (18:2 n-6, 56.5% of the total FAs), oleic acid (18: 1 n-9, 25.8%) and palmitic acid (16:0, 11.4%). The enzymatic acidolysis showed a positive change in the n-6/n-3 FAs ratio, increasing the n-3 family and decreasing the n-6 FAs due to the incorporation of n-3 LCPUFAs into SO. The n-6/n-3 FAs ratio decreased from 11:1 in the original SO to approximately 3.5:1 and 4.7:1 after acidolysis catalyzed by AN and RJ lipases, respectively. The data showed that in the conditions of run 6 the TAGs show lower oleic acid (18:1 n-9) and linoleic acid (18:2 n-6) content when compared to the original SO. Therefore, the incorporation of EPA and DHA increased palmitic acid (16:0) and stearic acid (18:0) content. For both lipases, EPA incorporation was higher than DHA incorporation, which may be due to the EPA content of the fish oil used in the acidolysis reaction (19.8%) being higher than its DHA content (11.4%).
Experimental model and levels of the variables used for optimizing the acidolysis of soybean oil by immobilized AN and RJ lipases.
| Runs | Variables | EPA+DHA (% total fatty acids) | ||||||
|---|---|---|---|---|---|---|---|---|
| Reaction time (X1, hour) | Sardine-FFA: (X2, soybean oil mole ratio) | Initial water content of lipase (X3, % w/w) | ANa | RJb | ||||
| 1 | −1 | 12 | −1 | 3:1 | −1 | 0.45 | 2.6 | 2.2 |
| 2 | +1 | 36 | −1 | 3:1 | −1 | 0.45 | 3.4 | 2.9 |
| 3 | −1 | 12 | +1 | 1:3 | −1 | 0.45 | 1.6 | 1.4 |
| 4 | +1 | 36 | +1 | 1:3 | −1 | 0.45 | 2.0 | 1.3 |
| 5 | −1 | 12 | −1 | 3:1 | +1 | 0.84 | 6.8 | 5.3 |
| 6 | +1 | 36 | −1 | 3:1 | +1 | 0.84 | 11.7 | 7.2 |
| 7 | −1 | 12 | +1 | 1:3 | +1 | 0.84 | 1.8 | 1.8 |
| 8 | +1 | 36 | +1 | 1:3 | +1 | 0.84 | 3.0 | 2.5 |
| 9 | 0 | 24 | 0 | 1:1 | 0 | 0.62 | 3.5 | 3.1 |
| 10 | 0 | 24 | 0 | 1:1 | 0 | 0.62 | 3.5 | 2.3 |
| 11 | 0 | 24 | 0 | 1:1 | 0 | 0.62 | 3.0 | 3.0 |
| 12 | 0 | 24 | 0 | 1:1 | 0 | 0.62 | 3.2 | 2.7 |
Fatty acid composition (% of the total fatty acids) of the purified STAGs obtained by acidolysis of soybean oil under different conditions.
| Fatty acids | Before reaction | After reaction (run 6) | |
|---|---|---|---|
| Soybean oil | ANa | RJb | |
| SFAsc | 12.6 | 13.5 | 17.3 |
| C16:0 | 11.4 | 12.5 | 13.8 |
| C18:0 | 1.2 | 2.0 | 3.5 |
| MUFAsd | 25.8 | 18.2 | 20.4 |
| C16:1 n-7 | ndf | 0.6 | 1.4 |
| C18:1 n-9 | 25.8 | 17.6 | 19.0 |
| PUFAse | 61.5 | 67.9 | 59.7 |
| C18:2 n-6 | 56.5 | 52.7 | 49.3 |
| C18:3 n-3 | 5.0 | 3.5 | 3.2 |
| EPA n-3 | ndf | 6.4 | 4.6 |
| DHA n-3 | ndf | 5.3 | 2.6 |
| EPA+DHA | ndf | 11.7 | 7.2 |
| n-3 | 5.0 | 15.2 | 10.4 |
| n-6 | 56.5 | 52.7 | 49.3 |
| n-6/n-3 | 11:1 | 3.5:1 | 4.7:1 |
Some lipases have been frequently used to discriminate between EPA and DHA in concentrates containing both of these fatty acids, thus providing the possibility of producing n-3 PUFA concentrates with dominance of either EPA or DHA. The selectivity of lipases is influenced by the positional distribution of the fatty acids and the glyceride structure due to the regiospecificity and triglyceride specificity of the lipase. A. niger lipase increased DHA and EPA concentration in cod liver oil, but did not increase them in menhaden oil.24,25 In the model system, the lipase from Candida rugosa showed the highest discrimination against DHA, while the lipases from Pseudomonas fluorescens and Pseudomonas cepacia discriminated against EPA the most. In the oils, the Pseudomonas lipases also discriminated against EPA the most, while DHA was initially discriminated the most by the lipase from Thermomyces lanuginosus. However, after longer reaction times the enrichment of DHA in the glyceride fraction of the oils was greatest for the lipase from C. rugosa.26
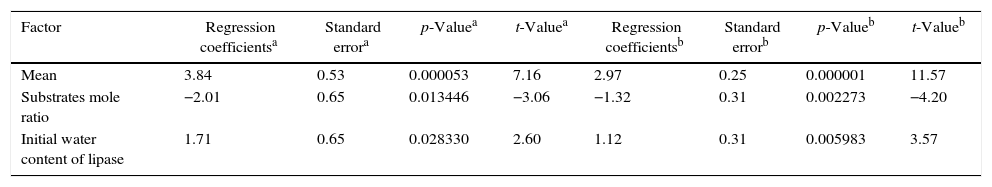
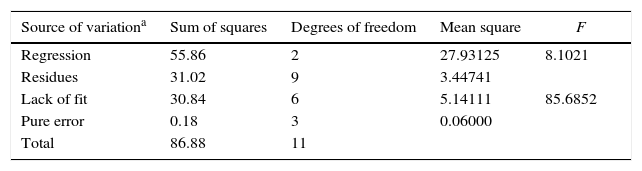
The statistical analysis showed that only the effects of the variables sardine-FFA:SO mole ratio and initial water content of lipase were statistically significant with a 95% confidence level (p<0.05). The estimated regression coefficients for both lipases are shown in Table 4 and the ANOVA for both lipases are shown in Table 5. The results were statistically validated by means of the F test. The R2 values were equal to 0.77 and 0.64. The models are described in Eqs. (1) and (2). With these models, it is possible to predict the optimal response (percentage of EPA and DHA) of the process in any conditions covered by the range of the experiment (Table 2).
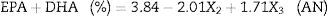
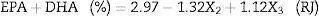
Regression coefficients for the analysis of the acidolysis reactions using AN and RJ lipases.
| Factor | Regression coefficientsa | Standard errora | p-Valuea | t-Valuea | Regression coefficientsb | Standard errorb | p-Valueb | t-Valueb |
|---|---|---|---|---|---|---|---|---|
| Mean | 3.84 | 0.53 | 0.000053 | 7.16 | 2.97 | 0.25 | 0.000001 | 11.57 |
| Substrates mole ratio | −2.01 | 0.65 | 0.013446 | −3.06 | −1.32 | 0.31 | 0.002273 | −4.20 |
| Initial water content of lipase | 1.71 | 0.65 | 0.028330 | 2.60 | 1.12 | 0.31 | 0.005983 | 3.57 |
Analysis of variance (ANOVA) for EPA+DHA incorporation (% total fatty acids) after acidolysis reactions using AN and RJ lipases.
| Source of variationa | Sum of squares | Degrees of freedom | Mean square | F |
|---|---|---|---|---|
| Regression | 55.86 | 2 | 27.93125 | 8.1021 |
| Residues | 31.02 | 9 | 3.44741 | |
| Lack of fit | 30.84 | 6 | 5.14111 | 85.6852 |
| Pure error | 0.18 | 3 | 0.06000 | |
| Total | 86.88 | 11 |
| Source of variationb | Sum of squares | Degrees of freedom | Mean square | F |
|---|---|---|---|---|
| Regression | 24.17000 | 2 | 12.08500 | 15.2492 |
| Residues | 7.13250 | 9 | 0.79250 | |
| Lack of fit | 6.74500 | 6 | 1.12417 | 8.7032 |
| Pure error | 0.38750 | 3 | 0.12917 | |
| Total | 31.30250 | 11 |
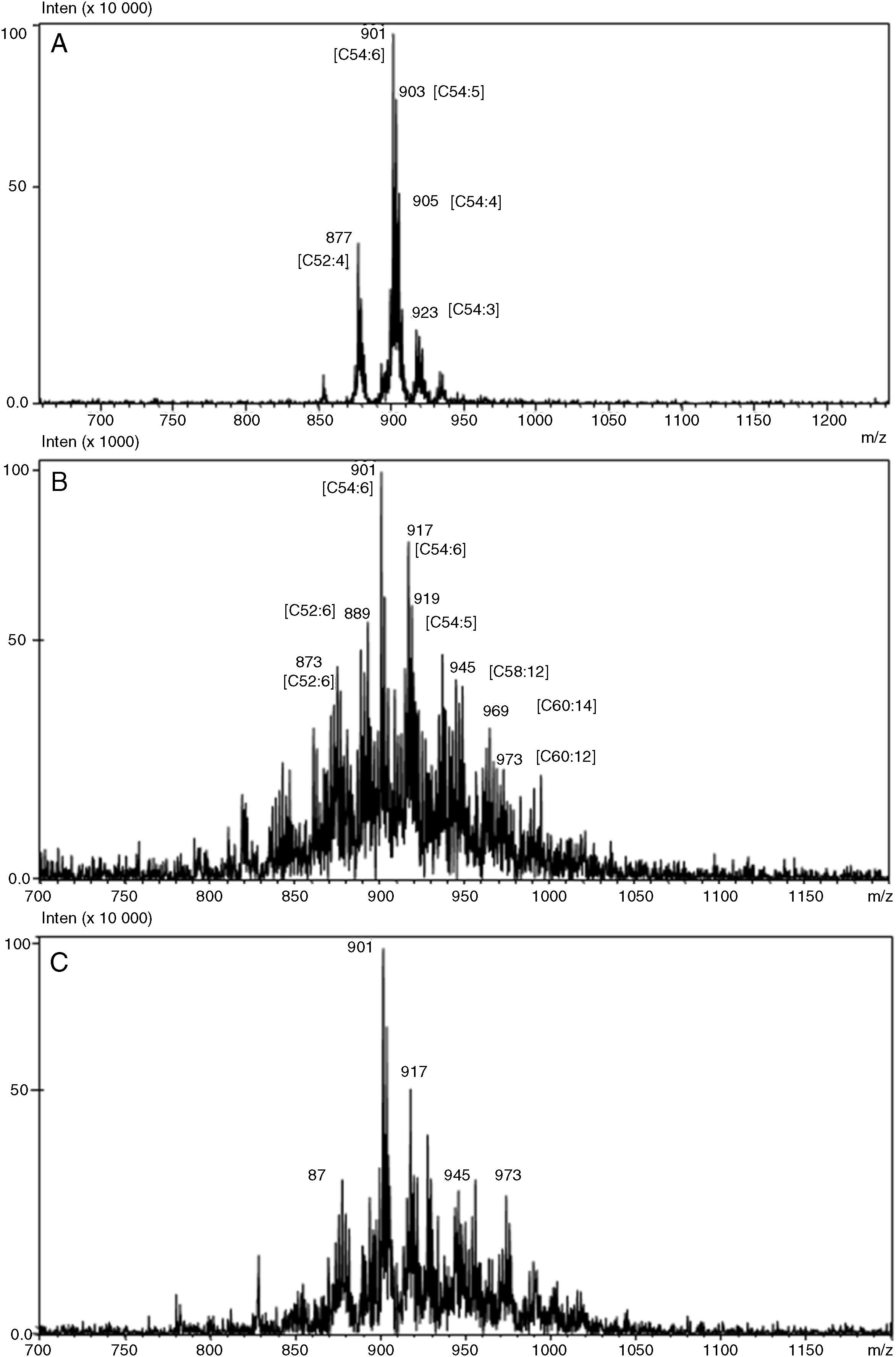
EASI-MS provides faster analysis with no sample preparation, using a more simple apparatus compared to other techniques. Fig. 2 shows TAG profiles obtained by EASI-MS for SO before (A) and after acidolysis catalyzed by AN (B) and RJ (C) lipases. Characteristic TAG of SO which, containing mostly linoleic (L), oleic (O), linolenic (Ln), palmitic (P) and stearic (S) acids, were detected mainly as [TAG+Na]+ ions and attributed to C52:4 (PLL, m/z 877), C52:3 (PLO, m/z 879), C54:7 (LLLn or OLnLn, m/z 899), C54:6 (LLL or OLLn, m/z 901), C54:5 (OLL or OOLn, m/z 903), C54:4 (OOL, m/z 905), C54:3 (OOO, m/z 907), C54:2 (SOO, m/z 909) and C54:1 (SSO, m/z 911) (Fig. 2A). Correspondent [TAG+K]+ ions of m/z 893 (PLL), m/z 895 (PLO), m/z 915 (LLLn or OLnLn), m/z 917 (LLL or OLLn), m/z 919 (OLL or OOLn), m/z 921 (OOL) and m/z 923 (OOO) were also detected. The lipase-catalyzed acidolysis of SO with sardine-FFA resulted in a substantially modified TAG profile mainly in the form of higher m/z (Fig. 2B and C). New, highly unsaturated TAG species were detected, showing a possible addition of DHA and EPA in the TAG molecules of SO, such as C52:6 of m/z 873 [MPDHA+Na]+, C54:6 of m/z 878 [POEPA+Na]+, C52:6 of m/z 889 [MPDHA+K]+, C54:6 of m/z 917 [LLL or OLLn or POEPA+K]+, C54:5 of m/z 919 [PSEPA+K)+, C58:12 of m/z 945 [LEPAEPA or MDHADHA+Na]+, C60:14 of m/z 969 [LnEPADHA+Na)+, C60:12 of m/z 973 [PDHADHA+Na]+ and C60:11 of m/z 975 (SEPADHA+Na]+. The data indicate a significant amount of LC-PUFA esterified into STAGs, which may be relevant in the development of alternative PUFA-formulations for clinical nutrition or incorporation into food products.
-MS for original soybean oil (A) and soybean oil modified by the acidolysis reactions catalyzed by Aspergillus niger (B) and Rhizopus javanicus (C) lipases.")
TAGs containing n-3 LCPUFAs can be obtained by lipase-catalyzed acidolysis or interesterification with fatty acid ethyl esters, either in organic or in solvent-free media, as reported in several studies.19,27–29 STAGs enriched with unsaturated fatty acids and low in palmitic acid content via acidolysis of microalgae oil (Schizochytrium sp) with oleic acid were produced using Lipozyme RM IM.30 The unsaturated fatty acids content increased from 70.20% to 90.9% at the sn-1,3 positions in the STAGs under the optimal condition (i.e., lipase load of 7%, molar ratio of microalgae oil TAGs to oleic acid of 1:3, and temperature of 65°C). In most of these studies, high-cost sn-1,3, regioselective, immobilized, commercial, microbial lipases have been used. The high cost of commercial lipases and the low operational stability of some of them have been recognized as the main constraints to the use of lipase-catalyzed processes for STAGs production. Therefore, the search for novel biocatalysts, presenting both high activity and operational stability, is a way to make enzymatic processes competitive.
ConclusionSTAGs containing EPA and DHA, showing a n-3/n-6 ratio within the proportions recommended as ideal for clinical nutrition, were successfully obtained by acidolysis catalyzed by native AN and RJ lipases immobilized on Amberlite MB-1. Under the conditions optimized by RSM, the acidolysis reactions using both lipases led to an advantageous exchange between the acyl radicals from TAGs of SO and FFAs obtained from Brazilian sardine oil. The immobilization on Amberlite MB-1 probably exerts a significant influence on the stabilization of the conformation of lipases and on their resistance to denaturation, favoring their storage time, especially for AN lipase. One of the greatest advantages of immobilization is to facilitate an enzyme's reusability, which is essential to render the process technologically and economically viable. Little information is available on the potential health benefits of the STAG molecules that are rich in n-3 LCPUFAs. Further work is needed to get to know the physiological effects of the use of STAGs produced by incorporation of EPA and/or DHA into SO for clinical nutrition.
Conflicts of interestThe authors declare no conflicts of interest.
The authors wish to thank the Brazilian Financing Agencies: Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). The authors also thank the Laboratório ThoMSon de Espectrometria de Massas, Instituto de Química (UNICAMP) and the Centro de Micologia da Universidade de Lisboa (Portugal) for the taxonomic identification of isolates.