


Cushing's syndrome is due to excess cortisol secretion and is associated to increased mortality and severe morbidity that are not fully reversible despite biochemical control. The syndrome consists of a set of systemic manifestations similar to those found in aging. Chronic stress, which also causes hyperstimulation of the hypothalamic-pituitary-adrenal axis, has been related to accelerated telomere shortening, oxidative damage, and cell aging. Although premature aging in patients with Cushing's syndrome could be related to environmental factors, the possibility that chronic exposure to hypercortisolism causes telomere shortening, and thus premature aging, cannot be ruled out. This review discusses the available evidence supporting a link between Cushing's syndrome and cell aging.
El síndrome de Cushing se debe a una hipersecreción de cortisol, asociado a una mayor mortalidad y una elevada morbilidad, que no es totalmente reversible a pesar del control bioquímico, presentando un conjunto de manifestaciones sistémicas similares a las que aparecen en el envejecimiento. El estrés crónico, que también conlleva una hiperestimulación del eje adrenal, se ha relacionado con el acortamiento telomérico acelerado, el daño oxidativo y el envejecimiento celular. A pesar de que el envejecimiento prematuro de los pacientes con síndrome de Cushing podría relacionarse con factores ambientales, no puede descartarse que la exposición crónica al hipercortisolismo determine un acortamiento telomérico y, por lo tanto, envejecimiento. En esta revisión se repasan las evidencias existentes que podrían relacionar el síndrome de Cushing y el envejecimiento celular prematuro.
Cushing's syndrome is a rare endocrine disease caused by either exogenous (iatrogenic) or endogenous cortisol hypersecretion due to an ACTH-secreting pituitary tumor (Cushing's disease) and, less commonly, a cortisol-secreting adrenal tumor or an ectopic ACTH-secreting tumor. The annual incidence of endogenous Cushing's syndrome is estimated at approximately 2/106 inhabitants. It is associated with significant morbidity and increased mortality.
The main systemic manifestations occurring in patients with Cushing's syndrome are those related to cardiovascular diseases or other problems associated with aging itself.1 Thus, phenotypically these patients usually have central obesity (moon face and limb atrophy), increased fat mass and decreased bone and lean mass, muscle weakness, increased susceptibility to infection, capillary fragility, hematomas, typical red wine striae, hirsutism, decreased libido, and metabolic syndrome components (hypertension, diabetes mellitus and insulin resistance, dislipidemia, vascular disease, atherosclerosis, etc.) with increased cardiovascular risk, as well as depression, cerebral atrophy, and impaired health-related quality of life.1,2
Although hypercortisolism may be cured after surgery (control of cortisol levels is achieved in up to 90% of operated patients) and with drugs, recent evidence shows that these patients continue to have a high risk of experiencing cardiovascular complications, osteopenia, and functional and structural brain changes associated with a poor health-related quality of life.3
On the other hand, biological aging is characterized by a progressive and predictable loss of coordinated tissue and cell functions leading to an increasing risk of vulnerability and disease. This impairment initially occurs as a loss of the ability to maintain homeostasis in stress situations, and in more advanced stages even under resting conditions.4 This process is species-specific, and changes become apparent in multiple organs and systems, although the mechanisms contributing to aging are not fully known.
Although excess hypercortisolism is probably related to these processes, the exact mechanisms responsible are not clear. This review will attempt to set out the hypotheses and mechanisms relating hypercortisolism and aging proposed to date.
Is specific morbidity reversible after the resolution of hypercortisolism?The resolution of hypercortisolism was initially thought to be followed by normalization of the morbidity caused by excess cortisol levels, but in the past decade it has been shown that normalization does not occur despite the resolution of hypercortisolism. Cardiovascular risk continues to be high in biochemically cured Cushing's syndrome even five years after cure.5 Complete recovery of bone mineral density (BMD) is more controversial. Some studies show improvement, or even normalization,6,7 but others do not.8,9 It has been shown that the duration of both endogenous hypercortisolism and replacement therapy with exogenous glucocorticoids after effective surgery for Cushing's syndrome negatively correlates to BMD in women 11 years after syndrome remission as compared to controls.10 Persistent central fat accumulation has been shown in cured patients with an unfavorable adipokine profile (decreased adiponectin levels, increased plasma levels of sTNF-R1 and interleukin-6), leading to a low-grade inflammation state.11 This “inflammatory state” may cause vascular damage, contributing to atherosclerosis and cardiovascular disease in these patients years after remission. Increased oxidative stress and endothelial dysfunction have been seen in Cushing's syndrome.12 High levels of superoxide anions, cyclo-oxygenase-1 (COX-1) and endothelial nitric oxide synthetase, causing endothelial dysfunction and increased peripheral vascular resistance, have been shown in microvessels.13 It has also recently been shown that glucocorticoids may directly activate the signaling pathway of mineralocorticoid receptors, regardless of circulating aldosterone levels.14 This suggests that they may contribute to this vascular and endothelial damage found in these patients through the profibrotic properties of aldosterone.15
In addition, as previously stated, hypercortisolism affects behavior, mood, neuronal activity, memory, and other central nervous system processes.1,2 Studies with 3-Tesla MRI have shown greater brain atrophy as compared to normal controls of similar age which does not appear to be fully reversible despite cortisol normalization. Psychopathology (particularly atypical depression), highly prevalent in active disease, improves in the first year following treatment, but residual symptoms often persist.16 High cortisol levels have been associated with changes in the function of some neurotransmitters, such as decreased cerebral serotonin synthesis, increased noradrenergic activity, and low levels of 5-hydroxyindoleacetic acid in the cerebrospinal fluid, all of them related to the pathogenesis of depression.17 Cognition 18and health-related quality of life do not appear to normalize either following the cure of hypercortisolism, suggesting that all these changes are not fully reversible.19,20 Several studies have reported that cortisol hypersecretion has an aging effect on cognitive behavior,21,22 but the mechanisms through which glucocorticoids affect the cognitive system are not clear. Several hypothetical mechanisms have been postulated, including hippocampal changes found in rats and the anatomical location of a large number of glucocorticoid receptors in the brain, especially in the hippocampus, amygdala, and prefrontal lobes, which would promote morphological changes in these areas as occurs in aging,23 or a marked reduction in cerebral glucose metabolism (visible as the atrophy and neuronal death typical of aging). This impaired cognitive function is evident even after eliminating the effect caused by depression, which is highly common in hypercortisolism and may markedly interfere with cognitive function. The reason why chronic hypercortisolism has these long-term consequences despite biochemical normalization not being clear, but a poorly reversible neurotoxic effect is suggested.
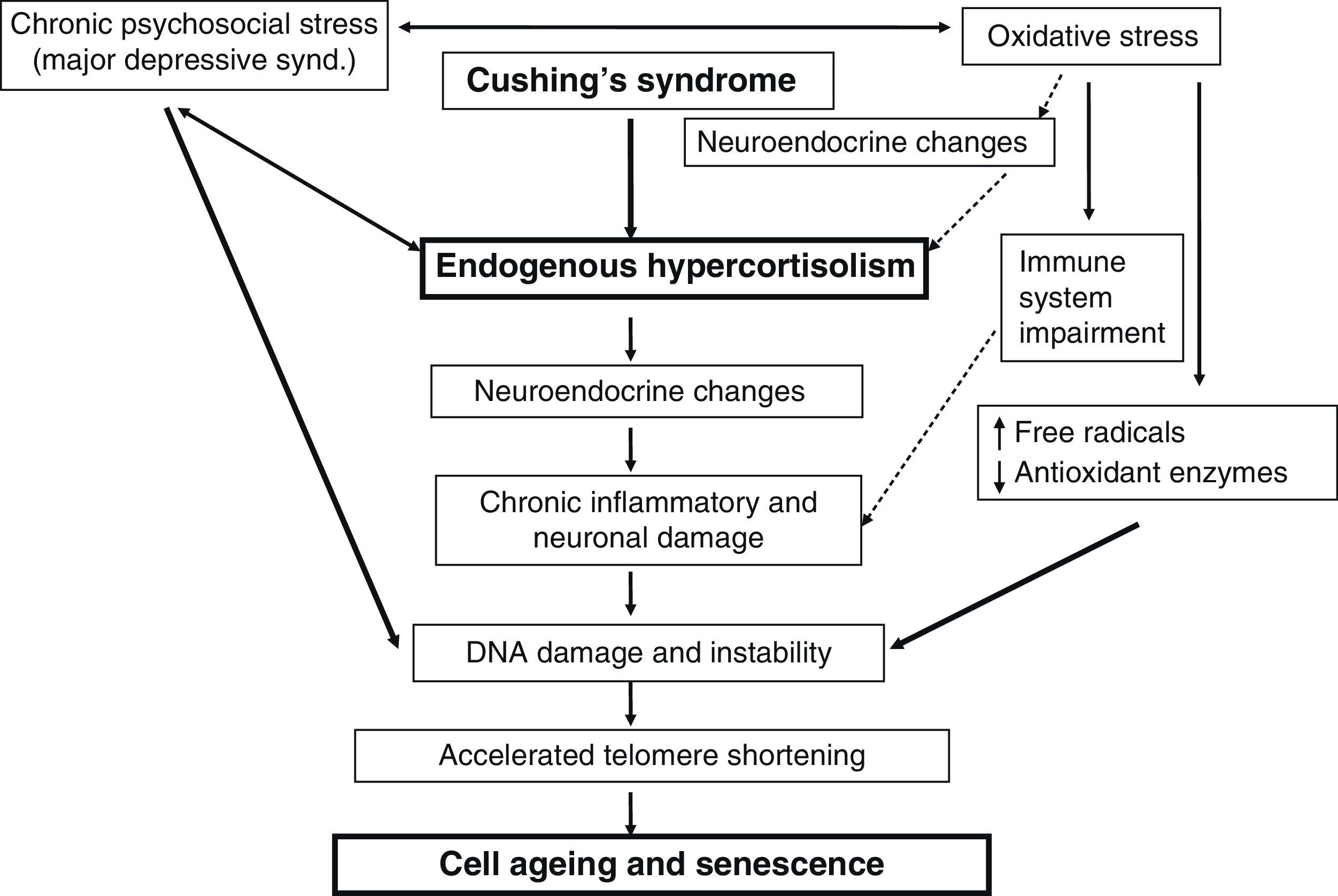
Chronic stress and hypercortisolismChronic stress causes overstimulation of the hypothalamic-pituitary-adrenal axis and endogenous hypercortisolism. In addition, it is known that more than half of the patients with chronic depression have adrenal axis overactivity, representing another model of endogenous hypercortisolism.24
At the hypothalamic level, chronic stress inferred by the brain cortex also results in corticotropin-releasing hormone (CRH) secretion leading to activation of the hypothalamic-pituitary-adrenal axis and, thus, cortisol release into the bloodstream. High CRH levels in the cerebrospinal fluid and high CRH mRNA levels in the limbic regions of the brain have been found in patients with chronic major depression.25,26 It has also been noted that clinical remission of major depressive disorder is often associated with the reversibility of cortisol abnormalities. However, depressive symptoms may persist despite the resolution of Cushing's syndrome.27 By contrast, it has also been suggested that chronic cortisol elevations, particularly nocturnal elevations (characteristic of endogenous hypercortisolism) may play a significant role in the pathogenesis of depression and in chronic stress.28 On the other hand, the role of 11-beta-hydroxysteroid reductase (11b-HSD), an enzyme that regulates glucocorticoid metabolism at intracellular level, has been suggested as one of the potential mechanisms by which hypercortisolism could exert harmful effects on the nervous system and behavior. There are two different isoforms of 11b-HSD: type 1, which would increase intracellular cortisol levels (in liver, fatty tissue, and brain), and type 2, which would inactivate glucocorticoids (from cortisol to an inactive cortisone molecule).29 Hippocampal cells only express 11b-HSD type 1, and cortisol levels are therefore increased. Since 11b-HSD type 2 is not expressed in the hippocampus, cortisol binds to the mineralocorticoid receptors (located in the hippocampus and limbic regions) with a high affinity (10-fold higher than that of the glucocorticoid receptors) and may have a significant effect in the pathogenesis of depression and in chronic stress,30 as well as contributing to the pathogenesis of metabolic syndrome (due to the increased action of the type 1 isoform seen in liver and fatty tissue).31
Chronic stress is also associated with a low health index, including an increase in cardiovascular risk factors and impaired immune function, similar to that which occurs in patients with Cushing's syndrome. However, the mechanisms by which this occurs have not been elucidated yet.
It appears that chronic stress could affect cell health and aging by different non-mutually exclusive and still undefined mechanisms. This would result in immune function impairment through oxidative stress (neuroendocrine response with hypercortisolism secondary to stress which would increase neuron damage and decrease antioxidant enzymes), and would finally cause an impairment in telomerase activity) (Fig. 1).32
Telomere activity and cell aging
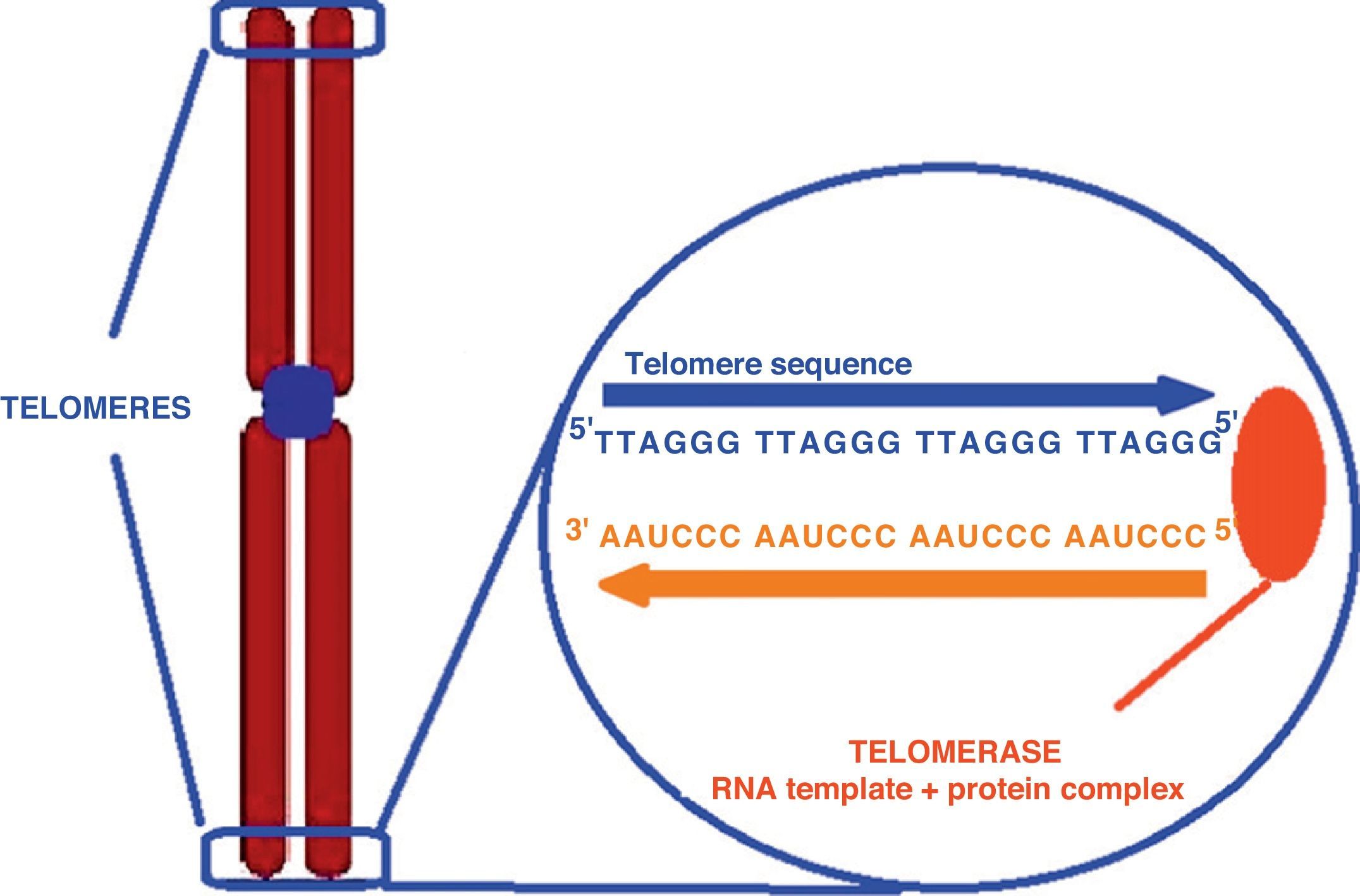
Oxidative cell damage and severe psychosocial stress have been related to telomere shortening. Chronic stress associated with mood changes may therefore contribute to excess vulnerability to diseases of aging such as cardiovascular diseases (through an accelerated aging of the body). The measurement of telomere length is currently a widely accepted means of assessing cell aging.33 Telomeres are non-coding DNA sequences in humans and other vertebrates which consist of tandem repetitions of a guanine-rich sequence TTAGGG (5′→3′). Telomeres are located at the chromosome ends forming a structure that protects them from erosion naturally occurring during the replication of linear DNA molecules in each division of eukaryotic cells (Fig. 2).34 Without telomeres, genetic material could be lost every time a cell divides.35 When telomere length becomes critically short, cell division stops and the cells become senescent and eventually die. To prevent this telomere attrition, germ cells, hemopoietic cells, and especially adult stem cells produce telomerase (found at high levels in the embryo and at decreasing levels after birth), an enzyme that catalyzes DNA synthesis to maintain telomere length and thus avoid cell senescence and apoptosis.26,35 By contrast, telomerase activity levels are very low and inconsistent in somatic cells and all other tissues.
.")
Telomeres may be shortened as the result of cumulative exposure to oxidative stress, genotoxic stressors and noxious stimuli and after chronic psychosocial stress, but this shortening has not been found to be consistent for all cell types and tissues.37
Telomere dysfunction involves significant clinical consequences. Diseases caused by an impaired or abnormal telomere system encompass a wide spectrum of clinical manifestations: acquired aplastic anemia (mutations in the telomerase complex), congenital dyskeratosis (mutation in dyskerin, an essential protein for telomere function), idiopathic pulmonary fibrosis (mutations in the telomerase complex in 15% of cases), etc.35
Telomere shortening has also been related to premature aging processes (e.g. atherosclerosis, Alzheimer's disease, osteopenia, or type 2 diabetes mellitus), an increased risk of cardiovascular disease, inflammation, and malignant transformation (e.g. colorectal cancer, inflammatory bowel disease, Barrett's esophagus, etc.), increased mortality in patients with coronary disease, and early myocardial infarction, and it has also been related to carotid intima-media thickness and early mortality.38,39
The role of changes in telomere biology in human disease is probably underestimated, and much research on the subject is still needed.
Are stress and aging related to telomeres?Major depressive disorder is associated with high morbidity and mortality, including an increase in diseases related to aging (cardiovascular disease, cancer, etc.). Stress caused by these diseases is considered a threat for body homeostasis and induces a chronic response (to restore balance) that may induce adaptations and changes leading to long-term biological damage.
The hypothesis that stress has an impact on health by modulating the cell aging rate is also supported by evidence associating psychological stress with high oxidative stress (autonomic activation and neuroendocrine response to stress), decreased telomerase activity, and telomere length shortening, all of them known determinants of cell senescence and longevity.40 These effects become evident the longer the duration of and exposure to stress. Chronic psychosocial stress has been shown to affect telomere length. Studies of patients with severe chronic stress (mothers caring for chronically ill children) found that a longer stress duration and a greater perception of stress levels significantly correlated to telomere length shortening and decreased telomerase activity as compared to age- and sex-matched controls, representing the equivalent of approximately 10 years of accelerated biological aging.41 Similar results have been found in patients with chronic major depression (who often have hypercortisolism). An additional study of 647 women with sisters with breast cancer assessed the relationship between telomere length (a marker of cell aging), the degree of stress perceived by these women, and stress hormone levels (cortisol and metanephrine) in urine. A trend to increased telomere shortening was found in women with a higher index of perceived stress, but these differences were only significant in women over 55 years, with significant recent psychosocial stress, and with higher urinary catecholamine levels. A non-significant trend to greater telomere shortening was seen with higher urinary cortisol levels.42 The biological impact of these telomere length results is not clear because of the wide range of changes in telomere length in the general population, which makes interpretation difficult. There are still many questions pending as regards the relationship between telomere length, aging, and the risk of disease.
It has been reported that there are people who have physiological resistance to oxidative stress, very likely due to still not well known genetic variants, which would increase their life expectancy. On the other hand, it has also been postulated that this physiological resistance to oxidative stress (which would lead to less hypercortisolism, less impairment in the telomere and immune systems, etc.) is related to physiological resistance to stress, in which case it would not be so evident that telomere shortening could be a sequela of prolonged exposure to psychological stress.40
Despite the high number of studies relating oxidative stress, chronic psychosocial stress, and hypercortisolism to median telomere length, some authors have recently considered that the percentage of short telomeres could possibly be a more sensitive marker of cell aging than mean telomere length in acute stress situations.41 This statement is based on the fact that telomere length is not consistent for all cells and tissues, or even for chromosomes of the same cell. Using a recently introduced technique (Universal STELA: Single Telomere Length Assay), it has been found in in vitro cell cultures that measurement of the percentage of short telomeres positively correlates to well known markers of cell senescence such as beta-galactosidase, but not to mean telomere length.42 This new method therefore appears to provide more information, contributing to a better understanding of telomere biology and its role in aging and cancer.
Hypercortisolism and telomere shorteningSome studies show that exposure to high cortisol levels is associated with a significant reduction in the telomerase activity of T lymphocytes.36 This may be one of the mechanisms causing patients with adrenal axis overstimulation by chronic stress, or by Cushing's syndrome, to have an impaired cell immunity and, thus, increased susceptibility to infection.
Little is known about the biological mechanisms of excess morbidity and mortality associated with mood changes, but the evidence from major depressive disorder shows that changes in the biological systems related to stress (neuroendocrine system, telomere system, etc.) are among the mechanisms contributing to such excess morbidity and mortality.
As occurs in endogenous Cushing's syndrome, chronic stress also leads to overstimulation of the hypothalamic-pituitary-adrenal axis and hypercortisolism. More than half the patients chronically depressed with significant chronic stress are known to have adrenal axis overactivity, and represent a model of endogenous hypercortisolism.35,43,44
Thus, since mood changes (depressive disorders) are associated with a high morbidity, increased mortality, and cell aging, it could be hypothesized that telomere shortening may underlie increased morbidity and premature aging, both paradigms of endogenous hypercortisolism.45 Interestingly, the shortening of white blood cell telomere length has been associated with high nocturnal urinary cortisol excretion.46 More recently, an in vitro study where cortisol was applied to T lymphocytes found a significant dose-dependent reduction in telomerase activity.36 However, to our knowledge the telomere system has not yet been evaluated in Cushing's syndrome, which is the most typical situation of endogenous hypercortisolism. It has only been evaluated in pseudo-Cushing's syndrome (chronic major depression and chronic psychosocial stress).
Role of telomere shortening in complications of Cushing's syndromeThe biological link between Cushing's syndrome (characterized by the abovementioned cluster of systemic complications), increased cardiovascular risk, and neuropsychological distress is cortisol hypersecretion, which is able to induce different pathogenetic mechanisms resulting in cardiovascular damage, premature aging, and increased mortality. High glucocorticoid levels may therefore contribute to premature aging. The possibility that environmental factors cause premature aging cannot be ruled out either.
Could prior chronic exposure to hypercortisolism in these patients cause premature aging through telomere shortening? Could the telomere attrition mechanism seen in chronic stress or in severe mood changes also occur in patients with Cushing's syndrome and contribute to their premature aging and increased residual morbidity and mortality? To our knowledge, telomere system function has not been evaluated yet in Cushing's syndrome, the most typical paradigm of endogenous hypercortisolism
To sum up, Cushing's syndrome is a rare condition which, despite biochemical control of hypercortisolism through treatment, is associated with premature aging with increased morbidity and mortality. The exact mechanisms by which this occurs are not clear. Although environmental factors may play a role in these untoward consequences, the possibility that chronic exposure to hypercortisolism induces accelerated telomere shortening and, thus, premature aging cannot be ruled out. The elucidation of these mechanisms could be the first step toward the prevention and improvement of the clinical consequences in these patients.
Conflicts of interestThe authors state that they have no conflicts of interest.
Please cite this article as: Aulinas A, et al. Telómeros, envejecimiento y síndrome de Cushing: ¿están relacionados? Endocrinol Nutr. 2013;60:329–35.
