






Introduction
The global ecology of West Nile virus (WNV) has experienced noteworthy changes in recent years. From an endemic pattern of infection in specific areas dotted with sporadic local outbreaks every 25-30 years1,2, this flavivirus has spread to non-endemic areas over the past 10 years, causing a significant burden of human and animal disease3-8. This expansion, the causes of which are not yet totally understood, has been featured by the forthcoming of human outbreaks preceded by severe and highly mortal epizootia affecting mainly indigenous birds9,10. An increase in WNV virulence has also been noted in these outbreaks, confirmed by human mortality ranging between 4.3% in Romania in 1996 and 16.7% in Israel in 20004,6. Because of these issues, WNV is now considered a paradigm of emerging pathogen1,3.
The presence of circulating WNV strains has also increased in the Mediterranean basin in the past years, in countries geographically and climatologically closely related to Spain, such as Italy, France, Tunisia, Algeria, and Morocco11,12. Many factors concur in our environment to enable the introduction and expansion of WNV in Spain: a) the ubiquitous presence of wetlands (particularly the Doñana National Park in southwestern Spain, one of the most extensive in Europe), and stable populations of various ornitophilic Culex spp. mosquitoes; b) the strategic geographical location for numerous migratory birds coming from endemic areas of Africa (e.g., storks and flamingos), which annually share these natural areas and wetlands with indigenous birds; and c) recent animal and human outbreaks occurring in many bordering countries, configuring a virtual "ring" of WNV circulation around Spain11,12.
Despite these issues, there is little data assessing the ecobiology and possible circulation of WNV in sylvatic cycles occurring in our geography. With respect to humans, no cases of confirmed acute infection have been declared, and the few existing seroepidemiologic studies were performed with techniques having poor specificity (indirect hemagglutination)13,14. Thus, the current impact of human exposure to WNV in Spain remains uncertain. The aim of the present study was to analyze the prevalence of past and recent WNV infections and the possible risk factors associated with exposure to WNV in a representative population from southern Spain.
Material and methods
Geographical area and study population
Individuals from the city of Seville and the northern, western, and southern parts of Seville province (1,194,570 inhabitants). The province of Seville (southwestern Spain) covers an area of 14,000 square kilometers and has 1,758,720 inhabitants (65% of whom live in urban areas). The capital of the province, Seville, has 705,000 inhabitants. This area is located close to numerous wetlands, in particular Doñana National Park, one of the most extensive wetlands in Europe, and the delta of the Guadalquivir river, both of which shelter a large variety of indigenous and migratory avian species. Numerous livestock exploitations (mainly of equine cattle) are established in the vicinity of the park. The area has a mild Mediterranean climate, with a mean temperature of 19.6 °C (12.9-26.3), and mean rainfall of 327 L/m2/year. The topography is divided into farms and crops (66%), woods (19%), pastures (7.5%), and settled areas (7.5%).
Sample size/stratification
Sample size was established for an estimated prevalence of past WNV infections of 5 ± 2.5% (confidence level 99%) in 504 subjects. A pre-stratification was performed using the Annual Statistics Report of Andalusia (2001 census)15, according to age distribution (0-14 [n = 110], 15-29 [n = 135], 30-44 [n = 98], 45-64 [n = 101], and > 64 [n = 60] years), and place of residence of the population (> 50,000 = urban [n = 310], 5000-50,000 = suburban [n = 157], < 5000 = rural [n = 37]).
Inclusion criteria and sample collection
Subjects of all age groups were selected by random telephone solicitation, with the exception of the youngest age group (0-14 years). Subjects from this last group were pre-selected in our two central laboratories, which receive blood samples from the entire area (primary care, outpatient clinics, and hospital). Based on this pre-selection, we ultimately collected samples originating from primary care centers and pre-anesthetic assessment in outpatient clinics (to avoid selection of hospitalized, chronically ill, or immunosuppressed children). Subsequently, legal guardians were asked by phone for informed consent to use the remaining serum for our study, and were questioned to detect possible exclusion criteria. Once informed consent had been granted, the serum sample was stored at 80 °C and an epidemiologic survey was performed on all participating subjects. The data collected included gender, age, profession, place of residence, outdoor activities, travel over the past 12 months to other parts of Spain and Europe, contact with animals, and insect bites in the past month. Risk professions were defined as those involving close contact with animals (farmers, stockbreeders, veterinarians), nature and wetlands (rangers, foresters), or mosquitoes (extermination and pest control workers). The following persons were excluded: those unable to answer the epidemiologic survey, those who had traveled to endemic areas in the previous 12 months, and persons previously vaccinated against yellow fever, hospitalized in the last 30 days, with febrile disease in the last 30 days, or with any type of immunosuppression.
Serological techniques
Samples were processed and tested for WNV IgG by enzyme-linked immunosorbent assay (ELISA) and for WNV IgM by antibody-capture ELISA (MAC-ELISA) (Focus Technologies, Rendon, VA, USA) according to the manufacturer's guidelines. Assays were performed in duplicate. All the positive samples were retested and the final result was the mean value of all determinations. When available, IgM-positive samples were tested with a confirmatory ELISA procedure recommended by the manufacturer (background subtract procedure).
Positive or indeterminate sera in both the IgG and IgM ELISA tests were assayed by a plaque reduction microneutralization test against WNV (PRNT). For this purpose, samples were tested in duplicate and assayed twice. Two-fold dilutions (25 μL) of the samples ranging from 1:16 to 1:256 were placed in a 96-well tissue culture microplate (Nunc A/S, Roskilde, Denmark) and mixed with 25 μL containing 100 tissue culture infectivity doses (100 TCID50)16 of a reference E101 WNV preparation. After 1 hour of incubation at 37 °C, 50 μL of a Vero E6 cell suspension containing 4x105 cells/μL was added to each well. Cultures were maintained for 7 days at 37 °C and 5% CO2, and then fixed with a solution containing 10% formaldehyde and 1% naphtol blue-black dye. The endpoint titer was defined as the highest serum dilution that showed no cytopathic effect in 50% of cell cultures. All these procedures were performed in a level-3 biosafety facility.
Sera showing an IgG/IgM titer against WNV underwent further commercial serological analyses against other flaviviruses (dengue and tick-borne encephalitis [TBE]). The assays for dengue IgG (Panbio Diagnostics, Queensland, Australia) and for TBE IgG and IgM (Dade Behring, Deerfield, IL, USA) were based on indirect ELISA; the method for dengue IgM was a μ -chain capture ELISA (Panbio Diagnostics).
Definition of WNV infection
Subjects with any ELISA IgG titer were considered as having possible past WNV infection; subjects with any MAC-ELISA IgM titer were considered as having possible recent WNV infection. Those with any ELISA IgG or MAC-ELISA IgM titer, subsequently confirmed by PRNT assay, were considered as having definitive past or recent WNV infection, respectively.
Statistical analysis
Seroprevalence was determined in the overall sample, and by place of residence and age group. Continuous variables are expressed as mean ± standard deviation. Univariate analysis was performed to determine possible risk factors associated with past infection. Univariate group comparisons were performed using the chi-square, Fisher, and Student t tests and correlations were determined with Spearman's Rho. Multivariate analysis of factors showing significant differences on univariate analysis was performed using a forward stepwise logistic regression model. Group differences were determined with the odds-ratio (OR) and 95% confidence interval (CI). Significance was set at a P-value of < 0.05.
Results
Five subjects (nearly 1% of the 504 included) presented anti-WNV IgG titers with a mean ELISA absorbance ratio of 2.72 ± 1 (table 1); none of the remaining sera had indeterminate absorbance ratios. Prevalence of definitive past WNV infection confirmed by PRNT was 0.6% (3 positive samples, with PRNT titers of 64, 128, and > 256, respectively (table 1), and was significantly higher in rural areas (5.4%), than in suburban (0.6%; P = 0.09), or urban areas (0%; P = 0.01). Mean age of subjects with past WNV infections was higher than that of uninfected subjects (65 ± 23 vs. 34 ± 22 years, P = 0.01). Comparisons of demographic and epidemiological features between subjects with and without past WNV infection are shown in table 2. No differences were detected with the exception of activity in any risk profession (P = 0.048), which was associated with prior WNV exposure. Multivariate analysis detected no independent risk factors associated with past WNV infection.
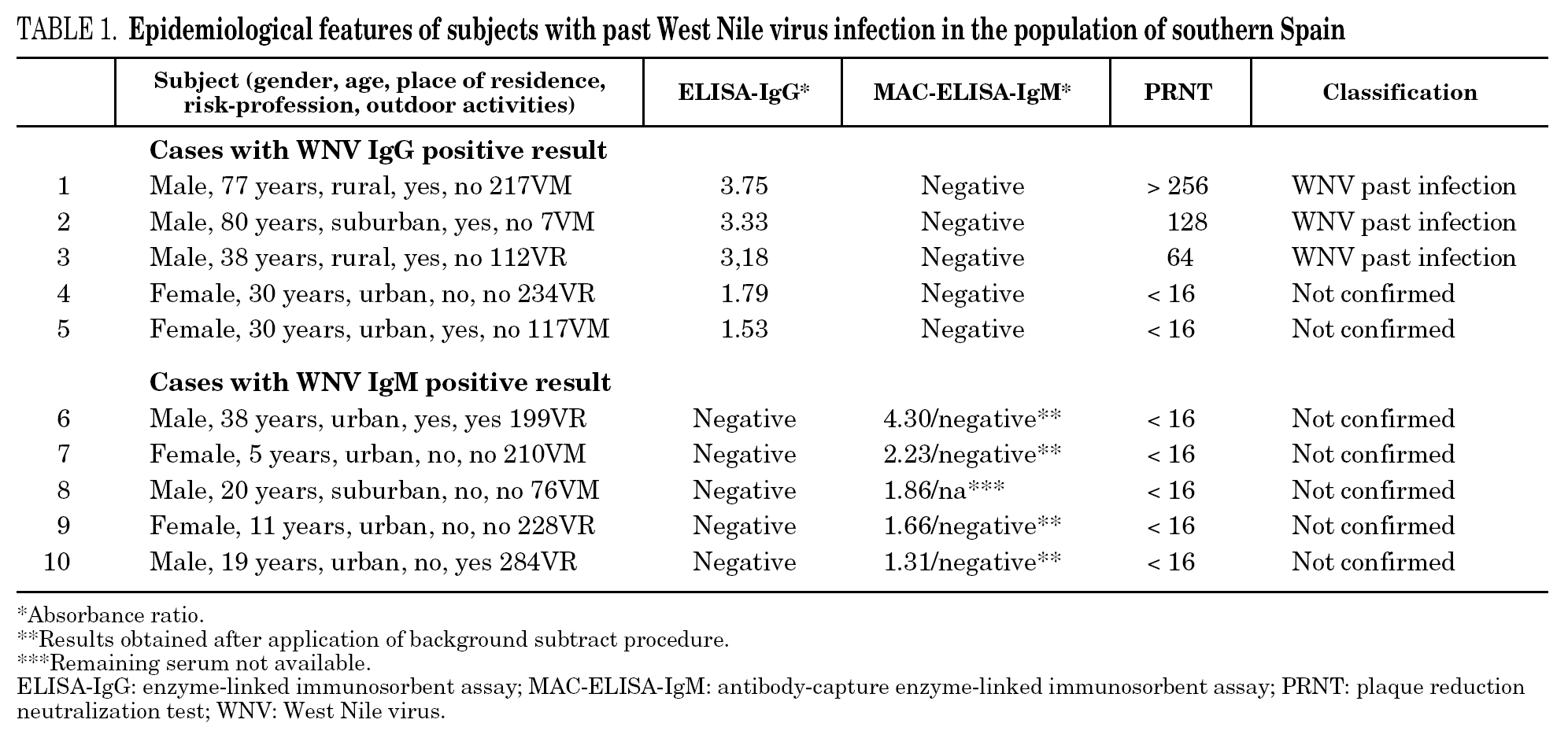

Five subjects (nearly 1% of the 504 included) showed IgM titers against WNV with a mean MAC-ELISA absorbance ratio of 2.03 ± 1.15 (table 1); WNV IgG antibodies were not detected in any of these subjects, and none of the remaining sera had indeterminate absorbance ratios. None of these individuals presented specific anti-WNV neutralizing antibodies when PRNT assay was performed (table 1). Application of a background subtract procedure to four of the remaining sera also yielded negative results.
Analysis of the IgG immune response against dengue and TBE viruses in the 5 subjects with anti-WNV IgG titers by ELISA showed IgG titers against dengue in sera from 3 subjects (subjects 1, 2, and 3 in table 1); no IgG immune response was detected against TBE virus. Similarly, analysis of IgM immune response against dengue and TBE viruses in the 5 subjects with anti-WNV IgM titers by ELISA detected IgM titers against dengue in the serum of one subject (subject 6 in table 1). After application of a subtract background assay to this serum sample (in the presence and absence of specific dengue antigen), the specific IgM immune response was not confirmed, and therefore ruled out. No IgM immune response against TBE virus was detected in any of the 5 subjects.
Discussion
The data from the present study show the presence of definite past WNV infection in the general population of southern Spain, confirming exposure of the population to this pathogen. These data complete previous preliminary studies carried out in Spain with less specific techniques that reported past infection rates of 8% in the human population of the northwestern part of the country and 3.1% in small mammals collected from several parts13,14,17,19. In addition, the findings are in accordance with the results of a similar recent study performed in northeastern areas of the country, which detected about 0.2% of confirmed WNV past infection in the general population20.
Definitive past WNV infection was characterized by affecting older persons, subjects living in rural areas, and those having risk professions. This epidemiological pattern probably reflects the means of acquisition, which may have been in the former past (older people are exposed during longer periods of time), and in subjects with closer contact to the wild cycle of WNV, since rural areas are located closer to wetlands, and therefore to reservoirs and potential vectors; the same is true for persons with risk professions.
As other authors have pointed out with respect to European countries, the present study raises questions as to the absence of human outbreaks of WNV disease in Spain, despite documented circulation of the virus, population exposure to WNV, and even declaration of sporadic outbreaks affecting equine cattle in surrounding countries8,21. This epidemiologic feature contrasts with the dynamics of WNV disease in North America, in which WNV has expanded considerably, causing more than 7,000 cases of human disease since 199922. Several factors may be related to this difference. Natural immunization of the majority of the population due to early exposure to WNV is hardly admissible, because the prevalence rates found in the present study are far from those reported in endemic areas, which range from 8% to 15%3,23. Partial immunization conferring some kind of protection against WNV due to population exposure to antigenically-related flaviviruses is also unlikely, because no indigenous cases or evidence of circulation of the Japanese encephalitis virus group or dengue virus have been reported in Spain, and yellow fever was eradicated in the final years of the 19th century. Exposure to other flaviviruses (mainly TBE virus, which is endemic in vast territories of Europe) could highlight this argument explaining the discordance between seroprevalences determined by ELISA (1%) and the definitive PRNT assay (0.6%) in the present study, and could also explain the recent human cases imported by Irish tourists after a visit to southern Portugal (an area close to Andalusia), whereas there is no evidence of native human cases despite detection of WNV in indigenous mosquitoes24,25. This explanation is not supported by our data, which showed a lack of specific antibodies against TBE virus in the sera tested. We hypothesize that the main reason may lie in climatic factors. The relatively warm, dry summer weather, together with the migratory pattern of avian species originating from endemic areas, which arrive to our wetlands in winter when the activity of potential indigenous biologic vectors is lower (they emerge from aquatic stages in spring and bite until fall), and when female mosquitoes begin diapause and feed less frequently, establish unfavorable environmental conditions for significant amplification of the cycle to "jump" to bridging vectors (mosquitoes that bite both birds and humans), and to indigenous avian species. Nonetheless, this pattern can easily change in the future, due to the global climatic changes we are witnessing26,27.
Another issue our data raises is the value of ELISA-IgG and MAC-ELISA in the diagnosis of recent and past WNV infection. The lack of specificity of these methods for accurate diagnosis of WNV infection has been discussed28,29, and discordance between the high specificity to detect immune response against WNV endowed by the technique's manufacturers and the well-recognized serological cross reactions against flaviviral antigens is a frequent and unsolved issue29-32. In fact, we detected IgG antibodies against dengue virus in the 3 subjects with specific neutralizing IgG titers against WNV, confirming cross-reactivity of the ELISA techniques with respect to closely related flaviviruses (dengue is not an endemic disease in Spain, and subjects with past WNV infections reported no travel to dengue endemic areas). Subjects with a single IgG against WNV, unconfirmed by neutralization are difficult to explain according to current knowledge. A plausible explanation might be prior exposure to an antigenic-related flavivirus unrecognized in our area. Clearly, future studies in this area are necessary, focusing on the impact of WNV (and other antigenic-related flavivirus) circulation in indigenous and migratory fauna, and on mosquitoes and other potential vectors (e.g., ixodidae and argasidae) to fully understand local epidemiology of WNV in humans33.
In conclusion, our data have documented the presence of definite past WNV infection in the general population of southern Spain. These infections were associated with older age, rural areas, and risk professions.
Acknowledgements
This study was financially supported by the Consejería de Salud, Junta de Andalucía (44/03), Spain.
Correspondence: Dr. M. Bernabeu-Wittel.
Servicio de Medicina Interna. Hospitales Universitarios Virgen del Rocío.
Avda. Manuel Siurot, s/n. 41013 Seville. Spain.
E-mail: MAXBW@telefonica.net
Manuscript received on October 9, 2006; accepted for publication on January 2, 2007.

