



Las áreas motoras corticales no solo son influenciadas por aferencias sensitivas periféricas y áreas de asociación prefrontales, sino también por los ganglios basales, específicamente el estriado. El estriado dorsomedial (EDM) y el estriado dorsolateral están involucrados en el aprendizaje espacial y el aprendizaje estímulo-respuesta; sin embargo, cada una de estas zonas pudiera mediar distintos componentes del aprendizaje. El propósito del estudio es determinar el efecto de la lesión electrolítica del EDM sobre el aprendizaje y ejecución de la conducta locomotora y sexual en ratas macho.
MétodoUna vez que los sujetos aprendieron a ejecutar las pruebas motoras de equilibrio, laberinto, rampa de ascenso y la conducta sexual, se realizó la lesión electrolítica del EDM. Cinco días después se realizaron las pruebas en 2 ocasiones más y se compararon las latencias de ejecución de cada prueba.
ResultadosDespués de la lesión, los valores promedio de latencia, incrementaron durante la ejecución de las pruebas de laberinto y equilibrio. Sin embargo, los valores promedio en la prueba rampa y conducta sexual, no aportaron efectos contrastantes entre los grupos.
ConclusionesLa lesión electrolítica del EDM modifica la ejecución de la actividad locomotora (prueba de laberinto y equilibrio), pero no la ejecución de la conducta sexual.
Cortical motor areas are influenced not only by peripheral sensory afferents and prefrontal association areas, but also by the basal ganglia, specifically the striatum. The dorsomedial striatum (DMS) and dorsolateral striatum are involved in both spatial and stimulus-response learning; however, each of these areas may mediate different components of learning. The aim of the study is to determine the effect of electrolytic lesion to the DMS on the learning and performance of sexual behaviour and locomotor activity in male rats.
MethodOnce the subjects had learned to perform motor tests of balance, maze navigation, ramp ascent, and sexual behaviour, they underwent electrolytic lesion to the DMS. Five days later, the tests were repeated on 2 occasions and researchers compared performance latencies for each test.
ResultsAverage latency values for performance on the maze and balance tests were higher after the lesion. However, the average values for the ramp test and for sexual behaviour did not differ between groups.
ConclusionsElectrolytic lesion of the DMS modifies the performance of locomotor activity (maze test and balance), but not of sexual behaviour.
Existe evidencia que indica que una de las funciones no motoras de los ganglios basales, involucra la participación del aprendizaje y memoria1,2. El papel de los ganglios basales en aprendizaje y memoria se ha descrito basado en hallazgos de estudios que han empleado la lesión eléctrica y farmacológica en animales de experimentación3-9. En algunos mamíferos las funciones del estriado dorsal, incluyendo el núcleo caudado y putamen, son generalmente consideradas en el contexto de control motor10. Sin embargo, otras funciones se han vuelto más evidentes en aspectos de integración sensoriomotora11, control cognitivo12, aprendizaje2,13 y algunos aspectos de función atencional14,15.
Diversos estudios han indicado que el estriado dorsal puede ser importante para el aprendizaje asociado (condicionamiento clásico) a un estímulo-respuesta (E-R)1,16,17. En particular se ha demostrado que las manipulaciones de esta región inducen déficits en una variedad de tareas de aprendizaje que requieren el procesamiento de información espacial en las pruebas de laberinto acuático de Morris18 y en el laberinto elevado en cruz19.
Sin embargo, el estriado no es una estructura homogénea y puede ser diferenciado en términos de su bioquímica intrínseca compartimental20,21 y en términos de su conectividad, sobre las bases de diversas proyecciones aferentes y eferentes16,22.
En ratas con lesión del EDL se mostró disminución en la eficiencia de discriminación durante una prueba de condicionamiento operante y una deficiencia para responder ante la presencia de CS+ y CS–. Este déficit es consistente con la noción de que el EDL está involucrado en el aprendizaje E-R y el aprendizaje de preferencia de lugar condicionado23,24. Sin embargo, la lesión del estriado dorsomedial (EDM) no muestra ningún efecto en el aprendizaje de las pruebas de preferencia de lugar condicionado,23 pero existe evidencia que la poslesión (después de haber aprendido una tarea de discriminación), si tiene participación en el aprendizaje E-R16,23.
El estriado es una estructura central clave para la adquisición y control del movimiento y más recientemente se ha reconocido que también para el aprendizaje E-R, preferencia de lugar, y procesamiento de información espacial, todos éstos involucrados durante la cópula. Por lo tanto, falta elucidar si el EDM subyace el aprendizaje de los patrones locomotores de la conducta sexual. Debido a esto, el propósito del presente estudio fue evaluar efecto de la poslesión electrolítica del EDM sobre la expresión de conductas locomotoras y sexuales previamente aprendidas.
Materiales y métodosSujetos de estudio y alojamientoRatas macho Wistar sexualmente expertas (250-350g) y hembras ovariectomizadas (200-250g) fueron usadas. Los machos seleccionados para los experimentos habían eyaculado al menos en 2 de 4 pruebas y exhibieron latencias de eyaculación de menos de 15 min. La receptividad de las hembras fui inducida mediante esteroides exógenos diluidos en aceite de canola. Inyecciones subcutáneas de benzoato de estradiol (10μg) y progesterona (500μg) se administraron 48 y 4h antes de cada prueba, respectivamente. Las ratas se alojaron en cajas colectivas de acrílico transparente (50×30×20cm) conteniendo una cama de aserrín, bajo condiciones de ciclo invertido de luz-obscuridad durante periodos de luz-oscuridad de 12×12h (las luces se activaron a las 08:00h) y mantenidos con alimento (LabDiet, Prolab 2500, RMH Rodent Diet. Fabricado por PMI Nutrition International, USA) y agua (Xallapan) a libre demanda. Todos los procedimientos quirúrgicos y las manipulaciones realizadas a las ratas se llevaron a cabo bajo las políticas de la Norma Oficial Mexicana NOM-062-ZOO-1991 y de la Society for Neuroscience sobre el uso de animales de experimentación.
Grupos experimentalesLas ratas (21 en total) fueron organizadas aleatoriamente en 3 grupos: se seleccionaron 7 animales como grupo control, 7 que recibieron una lesión electrolítica anodal bilateral en el EDM (grupo lesión) y 7 más con cirugía pero sin lesión (grupo testigo). Todos los machos seleccionados fueron sexualmente expertos.
Pruebas de laberintoEl laberinto octagonal usado en esta prueba fue diseñado en el laboratorio con plexiglás transparente. Todos los pasillos (35×10×20cm) estuvieron unidos a un fragmento central octagonal de plexiglás (10cm). La hembra receptiva se colocó siempre en el mismo pasillo, separada mediante una malla de plástico para prevenir el contacto con el macho. Los machos fueron probados cada 3 días durante una semana para adquirir las habilidades apropiadas para alcanzar a la hembra. Una vez expertos, los machos fueron probados 2 veces por semana durante 2 semanas. La primera semana representaron las pruebas antes de la cirugía y la segunda semana después de la cirugía. En todas las pruebas, la hembra se introdujo primero en su caja. Cinco minutos, después el macho se introdujo en el pasillo. Desde el inicio de la prueba se grabaron los movimientos del macho mediante una cámara (Sony DRC-SR20) y se analizaron mediante el programa The observer XT (Noldus Information Technology, USA). En esta prueba se analizó la latencia para alcanzar la hembra.
Prueba de barra horizontal (equilibrio)Esta prueba se realizó como se describe por Ortiz-Pulido et al.25. Este equipo fue elaborado en el laboratorio y consistió de dos cajas (40×30×15cm) de acrílico transparente, elevadas a 20cm del piso. Las cajas estuvieron separadas por una distancia de 45cm e interconectadas con una barra horizontal de madera de 2,5cm de ancho. En una de las cajas se colocó una hembra receptiva, separada mediante una malla de plástico para prevenir que cruzara la barra; el macho se colocó en la otra caja. A esta distancia, los machos eran capaces de detectar las señales sexuales provenientes de la hembra y así salir de la caja y caminar sobre la barra para alcanzar la caja de la hembra. Dentro de las conductas analizadas, se incluyó la latencia para salir de la caja, latencia para cruzar la barra y alcanzar la caja donde se encontraba la hembra. El mismo procedimiento se siguió en la prueba de rampa (barra de ascenso).
Prueba de rampa (barra de ascenso)El aparato usado para la prueba fue diseñado en el laboratorio y consistió de 2 cajas de acrílico (40×30×15cm), una colocada a nivel del piso y la otra en una posición elevada (30cm arriba del nivel del piso). La prueba se realizó como se describe en Ortiz-Pulido et al.25.
Prueba de conducta sexualAl final de cada una de las pruebas de laberinto, barra horizontal y rampa, los machos realizaron la cópula, la cual se realizó en el tercer periodo de oscuridad. En cada prueba, los machos se colocaron en una arena cilíndrica de Plexiglass (50×50cm, diámetro×altura). Después de un periodo de adaptación de 5min, se introdujo una hembra receptiva. Se analizaron los parámetros de número de montas (NM), intromisión (NI), eyaculación, sus latencias y se calculó el índice de intromisión NI/(NM+NI). Cuando el patrón eyaculatorio fue observado, finalizó la prueba de cópula y entonces los animales fueron regresados a sus cajas.
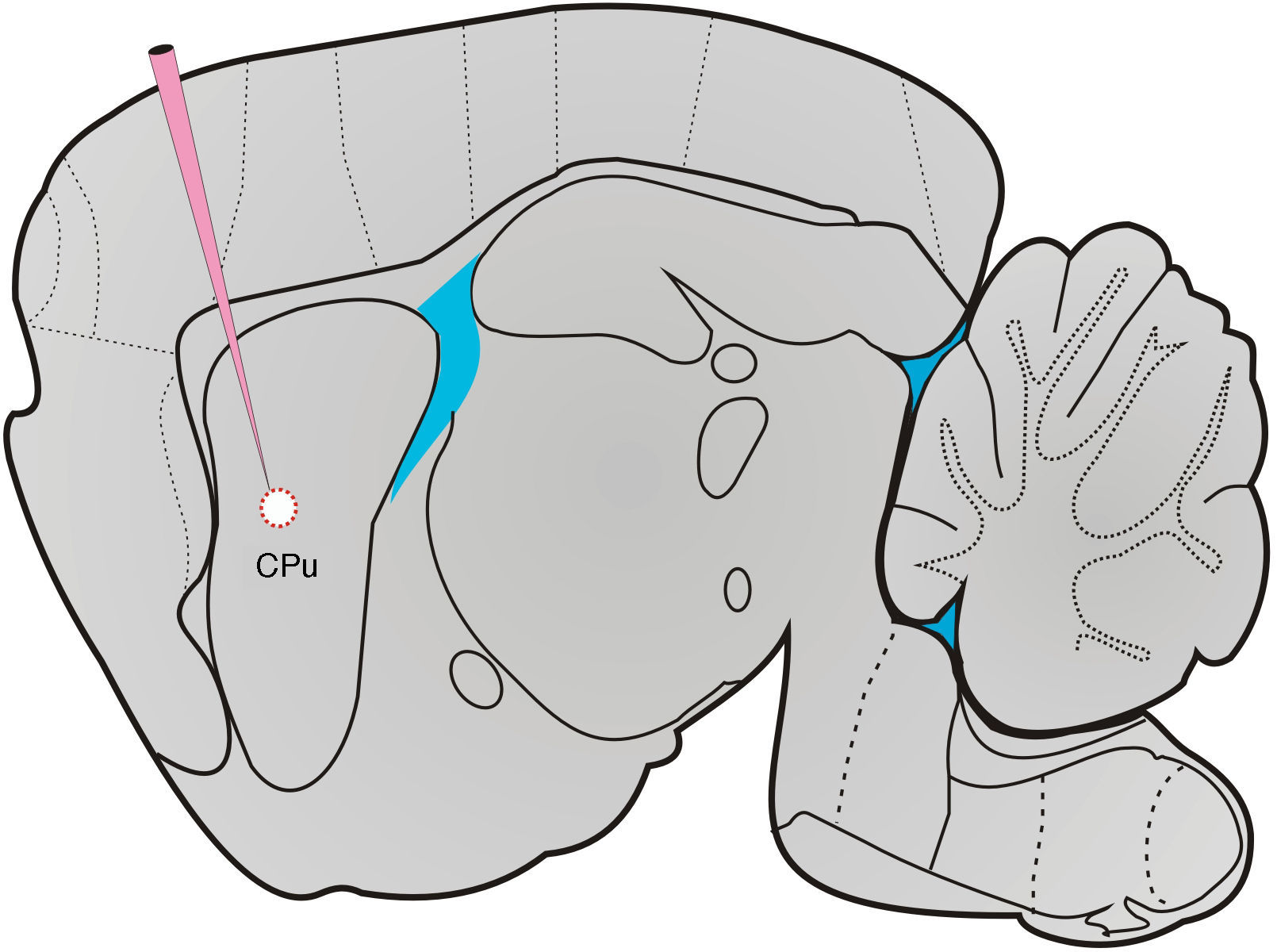
Cirugía y lesión electrolíticaLos machos lesionados y testigo fueron anestesiados con una mezcla de ketamina (50mg/kg) xilacina (8mg/kg). Inyecciones subcutáneas de solución salina fisiológica (0,9%) fueron administradas cada 30min para ser hidratados. Durante el periodo de anestesia profunda, se colocaron en un aparato estereotáxico (Stoelting Co. USA) y se realizó una incisión en la línea media para exponer la región del cráneo de lambda al borde posterior. Se realizaron 2 trépanos en las áreas correspondientes para descender bilateralmente electrodos unipolares de acero inoxidable de 250 micras (FHC) y realizaron lesiones anodales en la zona dorsomedial del núcleo estriado de acuerdo con las coordenadas estereotáxicas del atlas de Paxinos y Watson26, anteroposterior (AP)=0,0mm, lateral (L) +/–=3,0mm, ventral (V)=–5,5mm desde el punto Bregma. En los sujetos testigo, solo se descendió el electrodo pero no se aplicó corriente. En los sujetos experimentales, una corriente continua (0,5mA por 20 s) fue aplicada con una unidad CCU1 (Grass Astro-Med, Inc. USA), activada por pulsos de un estimulador S40 (Grass Astro-Med, Inc. USA). De 5 a 10 min después de la introducción del electrodo (testigo) y de aplicar la corriente (experimentales), se retiró el electrodo y se suturó la piel de la cabeza. Los animales se colocaron en una incubadora hasta su recuperación de la anestesia y posteriormente se hospedaron en cajas individuales en el bioterio.
HistologíaAl final de los experimentos, las ratas testigo y lesionadas fueron anestesiadas profundamente con pentobarbital sódico (60mg/kg) e inmediatamente perfundidas transcardialmente. El cráneo fue removido, se extrajo el cerebro y se colocó en una solución fijadora durante 24h. Se realizaron cortes coronales y sagitales del cerebro a 40μm y a una temperatura de –24°C. Los cortes fueron teñidos con violeta de cresilo, montados en portaobjetos gelatinizados y se fijaron con un cubreobjetos mediante Permount. Se tomaron imágenes con la ayuda de un microscopio (Olympus Provis, Ax70, Olympus, Co. Japan) para corroborar los sitios y las dimensiones de las lesiones (fig. 1).
Análisis estadístico
La variable de respuesta (latencia) fue procesada aplicando un análisis de varianza (ANOVA) con medias repetidas, en el que fueron analizadas las primeras 4 sesiones de entrenamiento locomotor en las pruebas de laberinto, equilibrio y rampa. Posteriormente, se analizaron la última sesión previa a la lesión electrolítica y las 2 subsecuentes. El modelo estadístico fue de un factor fijo (latencia) con 3 niveles, los cuales fueron un grupo de ratas con lesión eléctrica del EDM, otro testigo y un grupo de animales intactos, obtenido en las pruebas de laberinto equilibrio y rampa. Se verificaron los supuestos de homogeneidad de varianza y normalidad para cumplir con los lineamientos de la prueba estadística.
ResultadosLa respuesta de las ratas que fueron asignadas a los 3 grupos, y que fueron sometidas a 4 sesiones de entrenamiento a distintas pruebas locomotoras y entrenamiento de la conducta sexual, dio como resultado lo siguiente: la respuesta de las ratas a la prueba de equilibrio no difirió entre los factores fijos sesión y grupo como se esperaba, y la interacción entre sesiones por grupo tampoco aportó efecto. Durante la prueba de laberinto, las ratas tuvieron la misma latencia en respuesta a los factores fijos y la interacción entre sesiones por grupo no contribuyó con efecto alguno. En la pruebas de rampa (equilibrio), se registró el mismo tipo de respuesta en la latencia de las ratas, puesto que tampoco hubo efecto de los factores y de la interacción. La interacción entre sesiones por grupo en el índice de intromisión tampoco aportó efecto (tabla 1).
Latencia de respuesta e índice de intromisión en las pruebas locomotoras y de conducta sexual, respectivamente, durante 4 sesiones por los grupos a los que se asignaron los individuos después de la fase de entrenamiento
| F sesiones×grupo | p | |
|---|---|---|
| Equilibrio | 0,65 | 0,6 |
| Laberinto | 0,57 | 0,7 |
| Rampa base | 0,36 | 0,8 |
| Rampa subir | 0,04 | 0,9 |
| Índice de intromisión | 0,45 | 0,8 |
En cambio, después de las 4 sesiones de entrenamiento se realizaron los análisis de las variables de respuesta nuevamente para comparar 2 nuevas sesiones, posterior al daño por lesión eléctrica al EDM y se incluyó además la cuarta sesión de entrenamiento para así realizar el contraste antes y después de la lesión.
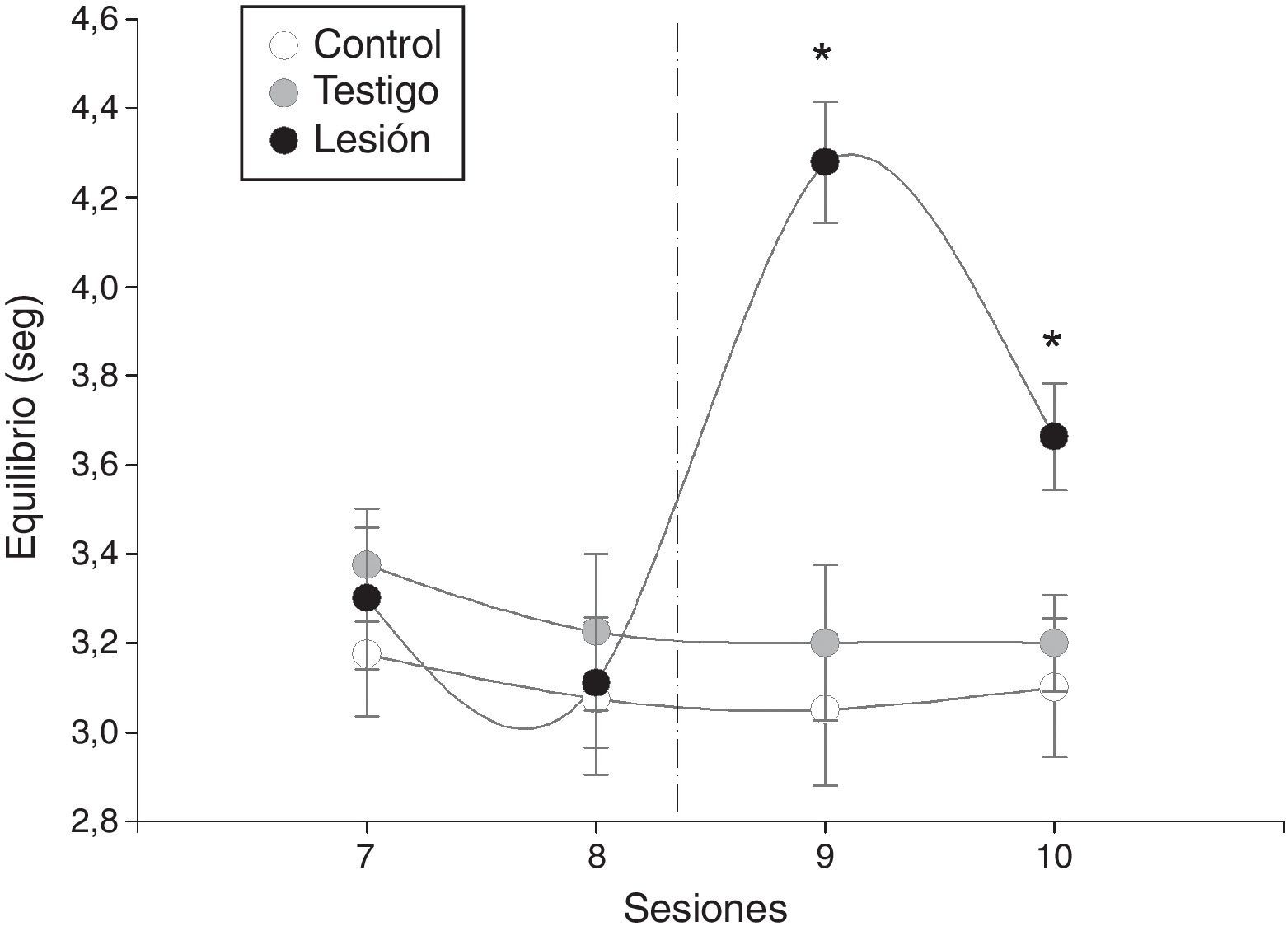
La latencia promedio de las individuos durante la prueba de equilibrio sí aportó efecto contrastante entre los grupos de ratas (F=16, p = 0,0001). Además, hubo diferencias entre las sesiones (F=10, p = 0,0003), y la interacción sesión por grupo muestra cómo los valores promedio difieren antes y después de la lesión en las ratas y no varía en los grupos control y testigo (fig. 2). Sin embargo, fue distinguida la repuesta de las ratas en el grupo que tuvo lesión eléctrica en el EDM debido a que el valor promedio de la latencia en la sesión inmediata al daño incrementó drásticamente y en la última sesión se redujo (fig. 2).
. En las pruebas previas a la lesión electrolítica la latencia que los machos requieren para cruzar la barra (y encontrar a la hembra receptiva) va disminuyendo, después de la lesión aumentan significativamente en comparación con los grupos control y testigo.")
Latencia promedio ± DE de los individuos durante la prueba de barra horizontal (equilibrio). En las pruebas previas a la lesión electrolítica la latencia que los machos requieren para cruzar la barra (y encontrar a la hembra receptiva) va disminuyendo, después de la lesión aumentan significativamente en comparación con los grupos control y testigo.
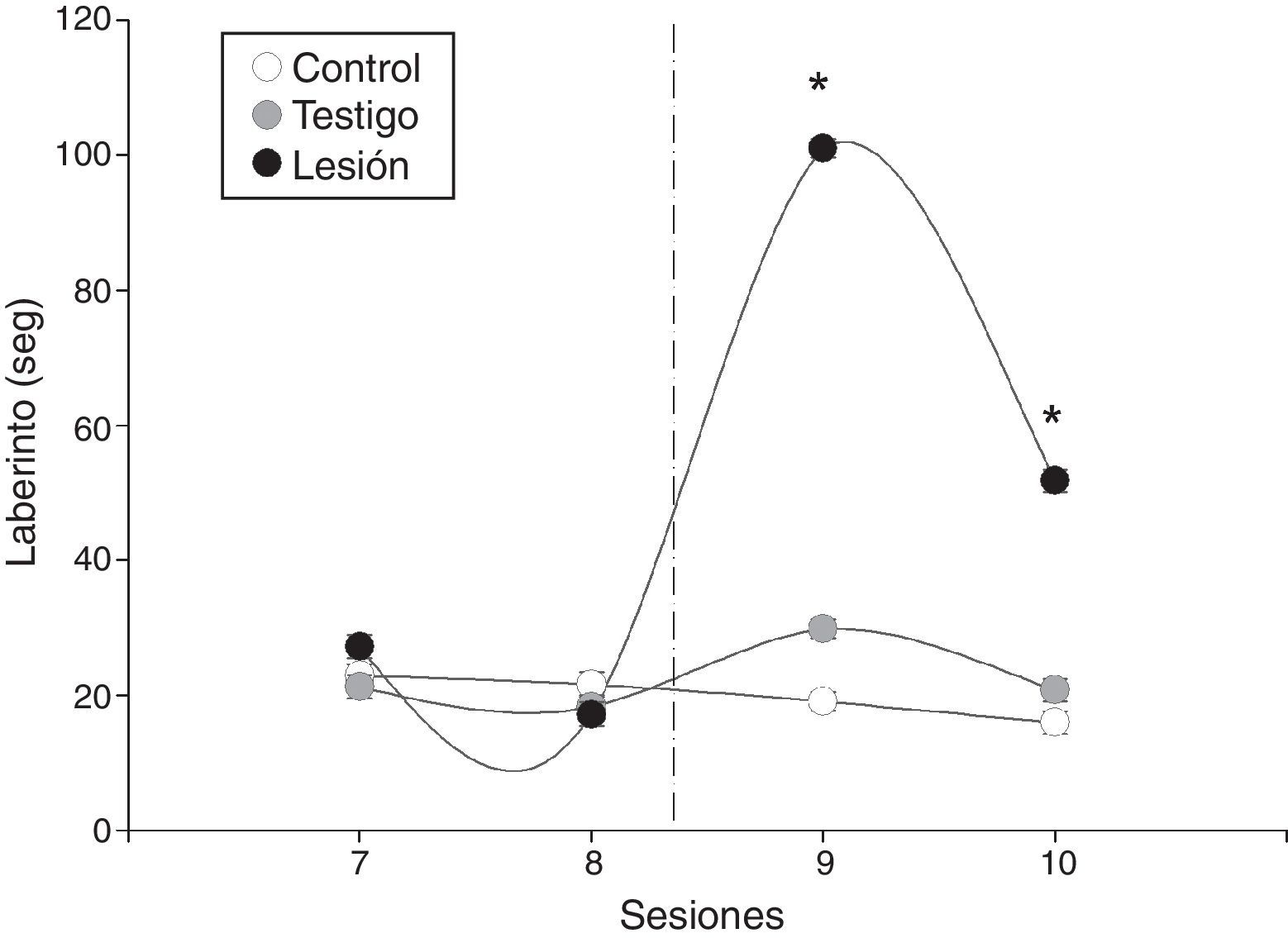
En la prueba de laberinto, las ratas también tuvieron distinto valor promedio en la latencia al contrastar entre los grupos de ratas (F=9,2, p = 0,001). También se registró variación al comparar entre sesiones (F=9,1, p = 0,0006), y la interacción sesión por grupo indicó también diferencias entre los valores promedio antes y después de la lesión en las ratas y no varía en los grupos control y testigo (fig. 3). En este sentido, las ratas del grupo que fue lesionado aumentó de forma amplia su valor promedio de latencia posterior al daño en el EDM, pero en la última la latencia permaneció con un valor alto con respecto a los otros grupos (fig. 3).

Latencia promedio ± DE de los individuos durante la prueba de laberinto. En las pruebas previas a la lesión electrolítica la latencia que los machos requieren para encontrar a la hembra receptiva va disminuyendo, posterior a la lesión electrolítica aumentan significativamente en comparación con los grupos control y testigo.
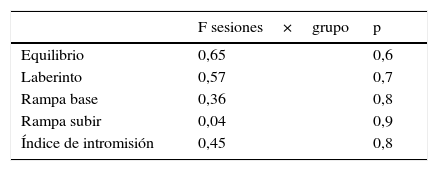
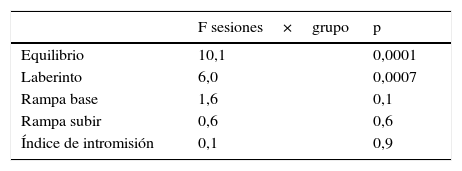
En cambio, los valores promedio de la latencia en los individuos a la prueba de rampa desde la base no aportó efecto contrastante entre los grupos de ratas (F=3,4, p = 0,06). Sin embargo, hubo diferencias entre las sesiones (F=9,7, p = 0,0005) debido al efecto de temporalidad no asociado a los grupos, lo cual se muestra en la homogeneidad en la interacción sesión por grupo que no aportó contraste (tabla 2). El mismo tipo de respuesta se registró en la prueba de la rampa —subir—, que no indicó diferencias entre los grupos (F=1,8, p = 0,19), aunque sí entre sesiones, que refiere al mismo efecto de temporalidad y no de tratamiento (F=3,5, p = 0,04), tal como se muestra en la no interacción de sesión por grupo. Posterior a la lesión del EDM, La interacción entre sesiones por grupo en el índice de intromisión tampoco aportó efecto (tabla 2).
Respuesta de la latencia e índice de intromisión de las ratas a las pruebas locomotoras y de conducta sexual, respectivamente, durante 3 sesiones por los grupos experimental testigo y control. Antes y después de la lesión electrofisiológica en el EDM
| F sesiones×grupo | p | |
|---|---|---|
| Equilibrio | 10,1 | 0,0001 |
| Laberinto | 6,0 | 0,0007 |
| Rampa base | 1,6 | 0,1 |
| Rampa subir | 0,6 | 0,6 |
| Índice de intromisión | 0,1 | 0,9 |
En nuestro estudio el análisis de la lesión del EDM demuestra que esta estructura es requerida para que los sujetos inicien una conducta locomotora, previamente aprendida, pero no afecta la ejecución de esta. El papel del EDM no es claro, sus funciones han sido principalmente consideradas motoras10; sin embargo, diversos estudios indican la participación del EDM en el aprendizaje E-R1,2,16. Experimentos previos han indicado que animales con lesión EDL27,28, pero no del EDM23, presentan un déficit en el aprendizaje de tareas que requieren la formación de asociaciones.
El principal hallazgo de nuestro estudio es que la lesión bilateral del EDM en la rata produce un déficit en la ejecución de 2 tareas locomotoras. Estos déficits consisten en un aumento en la latencia para iniciar las conductas. Todos los sujetos empleados en este estudio fueron sexualmente expertos y también fueron entrenados en las pruebas locomotoras (prueba de laberinto, equilibrio y rampa). Durante las pruebas de entrenamiento las latencias en los instrumentos fueron disminuyendo en todos los grupos conforme fueron adquiriendo habilidad en las pruebas (tabla 1); sin embargo, después de la lesión aumentaron considerablemente en las pruebas de laberinto y equilibrio en comparación con los grupos control y testigo (figs. 2 y 3) (tabla 2).
Nuestros resultados son consistentes con los observados por Rogers et al.15, quienes describen que la lesión del EDM produce aumentos en latencias en pruebas de condicionamiento pavloviano. Estos déficits pueden ser descritos como el resultado no solo de alteraciones en la función atencional, también a partir de las deficiencias en el requisito para organizar tanto el control atencional y la respuesta (p. ej., inhibitoria), procesos necesarios para el desempeño eficaz de una tarea15.
Los aumentos en las latencias, después de la lesión del EDM, en animales que previamente aprendieron una conducta, atribuyen estas deficiencias más a problemas motores que motivacionales. La ausencia de efectos en los patrones de la conducta sexual después de la lesión indica que la motivación de las ratas no se ha afectado. En este sentido, Baunez y Robbins29 encontraron que el agotamiento de dopamina en el estriado dorsal de la rata produce cambios significativos en la velocidad y la probabilidad para responder en pruebas de elección. Por lo tanto, la ejecución efectiva de una tarea (prueba de laberinto o equilibrio) requiere la organización de varios componentes cognitivos y procesos motores, incluyendo la atención visual, y la modulación de respuestas mientras ubica blancos visuales, auditivos u olfativos para la realización de la conducta consumatoria apropiada15. Así, el análisis de la conducta sexual en los animales con lesión del EDM no muestra alteraciones en el índice de intromisión. Estos resultados se ven respaldados por estudios similares en donde no se encuentran cambios en la expresión de algunas conductas aprendidas antes de la lesión27,28.
Nuestros resultados son consistentes con la hipótesis de que diferentes sectores del estriado están involucrados en la ejecución de diferentes tareas visuoespaciales.
Es posible que lesiones previas a la adquisición de la conducta sexual causen retardos en el aprendizaje o ejecución de esta conducta. Existe evidencia de que lesiones del EDM previas a la adquisición de una tarea retardan el aprendizaje de esta23,24.
En nuestro estudio, posterior a la lesión del EDM, solo se realizan 2 pruebas más. En la última prueba, se observa un decremento (recuperación) en las latencias de las pruebas de equilibrio y laberinto. Este decremento no alcanza los promedios de los grupos control y testigo. Esta recuperación podría deberse a un proceso de plasticidad; sin embargo, es necesario realizar más pruebas después de la lesión electrolítica para corroborar dicho fenómeno.
FinanciaciónEl estudio se realizó como parte del trabajo doctoral de ROP con número de becario CONACyT 207997 y tuvo financiamiento parcial del PROMEP-México PTC-195.
Conflicto de interesesLos autores declaramos que no existe conflicto de intereses.
Los autores agradecemos el apoyo a los doctores Grecia Herrera Meza y Armando J. Martínez-Chacón, por los comentarios a las versiones previas del manuscrito y la asistencia con el análisis estadístico.

