



INTRODUCCION
La masa ósea, al igual que otras características del organismo, tiene un componente genético. Aunque la importancia relativa de los factores hereditarios y los ambientes en la osteoporosis no ha sido totalmente aclarada, se estima que entre un 40% y un 80% de la variabilidad de la masa ósea puede depender de factores genéticos1-3. Los estrógenos desempeñan un importante papel en la homeostasis esquelética, tanto en mujeres como en varones4. De ahí que se haya explorado la posibilidad de que las diferencias en los receptores estrogénicos pudieran asociarse a diferencias en la masa ósea. En este sentido, varios autores han señalado que algunos alelos de regiones polimórficas del intrón 1 del receptor de estrógenos alfa (RE) se relacionan con diferencias en la densidad mineral ósea (DMO)5-7. Sin embargo, estos resultados no han sido confirmados en otros estudios, incluidos los realizados por nuestro grupo8. Recientemente se ha señalado que el análisis de los haplotipos del gen del RE, determinados conjuntamentet con las enzimas de restricción Pvull y Xbal tendría un poder discriminante superior al del análisis alélico convencional. En particular, se ha sugerido que las mujeres con haplotipo Px presentan una menor masa ósea9, de ahí que nos planteáramos este estudio con el objetivo de analizar si dicho haplotipo determina diferencias en el pico de masa ósea o el riesgo de fractura en nuestro medio.
MATERIAL Y MÉTODOS
Los detalles del estudio se describen en una publicación previa en la que se analizó la relación entre la masa ósea y los alelos del receptor de la vitamina D y de estrógenos8.
SUJETOS
La población estudiada se compuso de los siguientes grupos:
1) Una muestra de convivencia de 53 mujeres jóvenes sanas de 18-34 años (media 26), seleccionadas entre estudiantes de Medicina y personal hospitalario.
2) Sesenta y seis mujeres postmenopáusicas control de 43-85 años (media 61), sin fracturas, bien sanas (seguidas rutinariamente sin tratamiento hormonal en una consulta de menopausia), bien con coxartrosis.
3) Cuarenta y una mujeres ingresadas por fractura de cadera (66-96 años; media 84).
4) Setenta y seis mujeres con osteoporosis postmenopáusica y fractura vertebral, de 43-85 años (media 68), seguidas en nuestra consulta de osteoporosis.
En las mujeres jóvenes se midió la DMO en columna lumbar (L2-L4) y cuello de fémur, mediante un densitómetro de rayos X Hologic QDR 1000.
ANALISIS GENÉTICO
Se obtuvo ADN de sangre periférica mediante un método comercial, siguiendo las intrucciones del fabricante (Quiagen, Hilden, Alemania). Se amplificó mediante reacción en cadena de la polimerasa (PCR), con un termociclador Perkin-Elmer 2400, una región del gen del RE que contiene los sitios de corte de las enzimas Xbal y Pvull. Los reactivos incluían 10 μl de la solución de ADN; 1,5 mM MgCl2, 2U de Taq polimerasa; 0,2 mM dNTPs y los amplímeros siguientes:
5'-CTGCCACCCTATCTGTATCTTTTCCTATTCTCC-3' y
5'-TCTTTCTCTGCCACCCTGGCGTCGATTATCTGA-3'
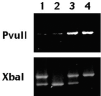
Después de una desnaturalización a 94° C durante 5 minutos, los tubos se sometieron a 30 ciclos de amplificación (94° C durante 15 segundos, 60° C durante 30 segundos, 72° C durante 30 segundos) y una extensión final a 72° C durante 7 minutos. Los productos de la PCR se digirieron en las enzimas durante 2 horas. Los fragmentos resultantes se separaron mediante electroforesis en gel de agarosa y se tiñeron con bromuro de etidio. Los alelos que contenían el sitio de restricción se designaron con letras minúsculas; los que no lo poseían con mayúsculas (fig. 1). En una fracción del 10% de las muestras tomadas al azar se repitió el análisis. La reproducibilidad fue perfecta en todo los casos.
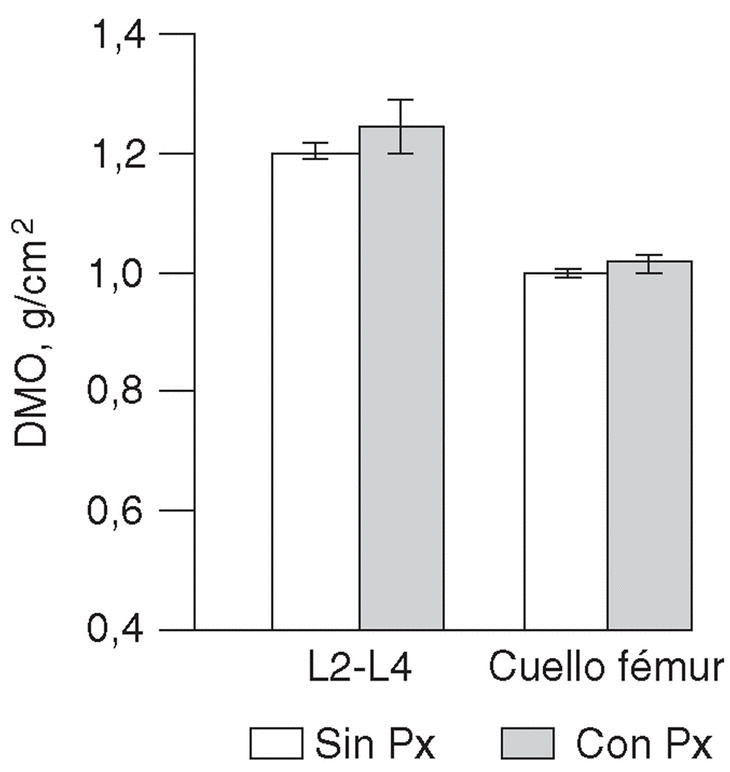
Fig. 1. Fragmentos resultantes de la digestión con Pvull y Xbal. Las muestras 1 a 4 corresponden a genotipos PpXx, Ppxx, PPXx y PPXX, respectivamente.
ANALISIS DE LOS RESULTADOS
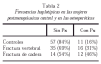
Se dividieron las mujeres según presentaran o no el haplotipo Px. Se excluyeron del análisis las que tenían un genotipo ambiguo que no permitía identificar el haplotipo con seguridad (tabla 1).

Se analizaron las diferencias en la DMO mediante la «t» de Student. Se analizaron las diferencias en las frecuencias de los haplotipos entre las mujeres con fracturas osteoporóticas (tabla 2) y las controles pre y postmenopáusicas mediante la prueba de Chi cuadrado. Puesto que no había diferencias significativas entre ambos subgrupos control se consideraron conjuntamente, a fin de aumentar la potencia estadística. Se calcularon las odds ratios y su intervalo de confianza con el programa estadístico Epilnfo 6.

RESULTADOS
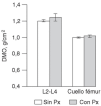
No se encontraron diferencias entre las mujeres jóvenes sin haplotipo Px y con él en cuanto al índice de masa corporal (21 ± 2 kg/m2 en ambos subgrupos) o la DMO (fig. 2). Los índices T fueron 0,4 ± 1,0 y 0,2 ± 1,1, respectivamente.
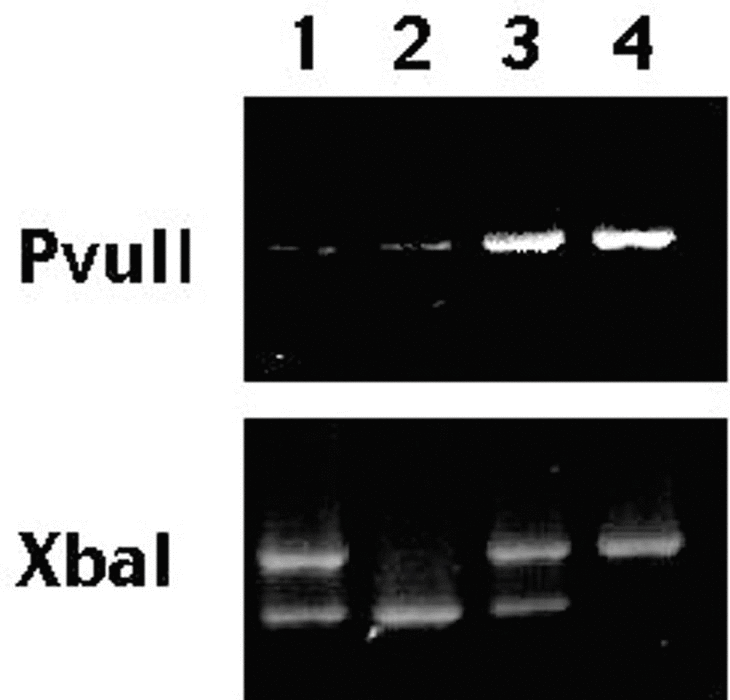
Fig. 2. Densidad mineral ósea en columna lumbar y cuello de fémur en mujeres jóvenes, dependiendo de que presentaran o no el haplotipo Px. DMO: densidad mineral ósea.
Sin embargo, el haplotipo Px tendió a ser más frecuente en las mujeres con fracturas que en las controles (tabla 2), aunque sólo alcanzó significación estadística en el caso de las fracturas de cadera (p = 0,03). La odds ratio de la asociación entre la presencia del haplotipo Px y las fracturas de cadera fue de 2,7 (IC 95% 1,1-6,9).
DISCUSION
El gen del RE-alfa está situado en el cromosoma 6. Contiene 8 exones y tiene un tamaño de unas 140 kilobases. Los polimorfismos A-G y T-C aquí analizados se encuentran en el intrón 1, a 350 y 400 pares de bases, respectivamente, del exón 2. La posible relación entre los polimorfismos del RE y la masa ósea es controvertida. Algunos autores han encontrado una menor masa ósea en las mujeres con alelos «P»5,10, pero otros no han confirmado esos hallazgos11. Nosotros mismos no hemos encontrado relación entre los alelos determinados por las enzimas Pvull y Xbal y el riesgo de fractura, cuando se analizan esos loci por separado8. En el presente estudio, al reanalizar los datos según los haplotipos resultantes de considerar conjuntamente ambos loci, encontramos que las mujeres con fracturas de cadera tenían una mayor frecuencia del haplotipo Px.
Así, nuestros resultados son concordantes con los de Albagha et al9. Estos autores estudiaron un grupo de mujeres escocesas con una edad media de 64 años y encontraron que la DMO en columna lumbar y cuello de fémur era significativamente menor en las que presentaban el haplotipo Px (estando la amplitud de la diferencia en torno a 0,5 desviaciones estándar). Es interesante que en ese estudio tampoco se hallaron diferencias significativas en la masa ósea cuando se consideraban por separado ambos loci, aun cuando existe un marcado desequilibrio de unión entre ellos (la presencia de alelos «P» tiende a asociarse con los «X», y los «p» con los «x»). En contra de lo señalado por otros autores6, el hecho de que no encontremos diferencias entre la DMO de las mujeres jóvenes con haplotipo Px y sin él sugiere que en nuestro medio el aumento del riesgo de fracturas en las portadoras del haplotipo Px no estaría en relación con un menor pico de masa ósea, sino con disminuciones posteriores de la misma.
El mecanismo responsable de estas asociaciones es desconocido. Dado que a menudo los intrones incluyen regiones moduladoras de la síntesis de ARN, algunos autores han sugerido que los alelos P representarían secuencias con menor afinidad por ciertos factores de transcripción, pero no está claramente demostrado que el intrón 1 del RE desempeñe un papel regulador de la transcripción. Más bien, la asociación del haplotipo Px con menor masa ósea y fracturas, en ausencia de relación con los alelos respectivos considerados aisladamente, apunta la posibilidad de que exista un desequilibrio de unión entre estos haplotipo y otras secuencias polimórficas, en el propio gen del RE o de otros genes próximos, que sean las realmente influyentes en el balance esquelético. De hecho, se ha descrito la existencia de un desequilibrio de unión entre los alelos del intrón 1 aquí analizados y una secuencia polimórfica repetitiva en tándem TA del promotor del gen del RE. Sin embargo, la relación entre los alelos en esta región repetitiva y la masa ósea es también controvertida9,11.
En conclusión, los polimorfismos del intrón 1 del RE parecen asociarse a diferencias en el riesgo de fracturas osteoporóticas, de manera que las mujeres con haplotipo Px presentan un mayor riesgo de fractura de cadera. No obstante, se necesitan estudios más amplios para confirmar estos resultados.