




Se estudió la distribución en los forófitos y se detalla la producción de frutos de las 46 plantas de Telipogon helleri, recientemente registradas en México por Solano et al. (2011). Se caracterizó el cafetal, se determinaron las clases de vida de cada planta de orquídea, inmaduras y adultas, distribución vertical; tronco, horquetas, ramas y ramillas; y orientación que tenían en los forófitos. De las plantas de T. helleri, 97.83% son adultas, lo que sugiere limitación en el reclutamiento. Sobre cafetos crecían 10 plantasde T. helleri, 21.74% del total y 36 ocupaban árboles de sombra (78.26%). Incluyendo cafetos y árboles, 73.92% de la población (34 plantas) crecía en el tronco. Entre 2.1-4m de altura estaba 78.26% de las plantas. Se contabilizaron 51 inflorescencias con 181 flores, de las cuales 23.76% formaron frutos (43 frutos). Telipogon helleri aún tiene una población muy incipiente y localizada, siendo imprescindible monitorearla para ver su comportamiento a largo plazo y evaluar su relación con las demás especies de epífitas vasculares presentes en el agroecosistema cafetalero.
The phorophytes distribution was studied and the fruit production of the 46 Telipogon helleri (Orchidaceae) plants, recently registered in Mexico by Solano et al. (2011), was detailed. The coffee plantation was characterized, the life stages of every orchid plant (immature and adult), the vertical distribution (trunk, forks, branches, twigs) and the orientation they had in the phorophytes were determined. From T. helleri plants, the 97.83% are adults, what suggests there is recruiting limitation. A number of 10 T. helleri plants grew up over coffee bushes (21.74% of the total) and 36 of the same species occupied shade trees (78.26%). Considering coffee bushes and shade trees, the 73.92% of the population (34 plants) grew up in the trunk. The 78.26% of the plants was between 2.1-4m of height. A quantity of 51 inflorescences with 181 flowers was assessed, from which the 23.76% formed fruits (43 fruits). Telipogon helleri still has a very incipient and located population, that is why it is essential to monitor it in order to see what its behavior might be in future and to evaluate its relationship with the rest of vascular epiphytes present in the coffe agroecosystem.
México, con su variedad de ecosistemas, posee una gran diversidad de orquídeas, con aproximadamente 1 150 especies registradas actualmente (Espejo et al., 2004), cifra que probablemente aumente a 1 300-1 400 especies, con la continuidad de los estudios de esta familia botánica (Hágsater et al., 2005).
La región del Soconusco en el estado de Chiapas, al suroeste del país, fronteriza con Guatemala, tiene una superficie de 5 475km2 que incluye llanuras costeras y parte de la cordillera de la sierra Madre, con ecosistemas de bosques tropicales y templados (Sánchez y Jarquín, 2008). A pesar de la sistemática destrucción de los ecosistemas de la región, podemos encontrar 293 especies de orquídeas, considerándose el corredor biológico Tacaná-Boquerón como la segunda región con mayor riqueza de orquídeas en México, después de El Mormón-Las Margaritas-Montebello (Damon, 2011).
El bosque mesófilo de montaña o bosque nublado, es el ecosistema más rico en orquídeas del mundo y del país, 60% de la flora de orquídeas de México es originaria de este hábitat. En el Soconusco aún existen fragmentos de bosque nublado, pero la mayor parte ha sido transformado en plantaciones de café (Coffea arabica L.) (Hágsater et al., 2005). No obstante, este tipo de cafetales siempre que sean cultivos de bajo impacto o tradicionales de sombra, es uno de los agroecosistemas perennes que más semejan las condiciones de las selvas y bosques originales, y en ellos logran desarrollar su ciclo de vida numerosas especies de plantas y animales (Moguel y Toledo, 1999; Sherry, 2000; Espejo et al., 2004; Hágsater et al., 2005; Solis-Montero et al., 2005; Manson et al., 2008a, b). Al menos 213 especies de orquídeas, 18.52% del total de México, se pueden encontrar dentro de las plantaciones de café (Espejo et al., 2004).
Telipogon helleri (L. O. Williams) N. H. Williams y Dressler fue registrada por primera vez para México en 2009, con una población de 46 plantas creciendo en Benito Juárez El Plan, en la zona de amortiguamiento de la Reserva de la Biosfera Volcán Tacaná de la región del Soconusco, Chiapas, en un cafetal tradicional de sombra, derivado de bosque mesófilo de montaña con elementos de la selva mediana perennifolia (Solano et al., 2011). Previamente la especie fue reportada para Nicaragua (Williams, 1962) y posiblemente también esté presente en Costa Rica y Guatemala (Solano et al., 2011).
Telipogon helleri es una planta epífita con una forma poco usual dentro de la familia Orchidaceae. Es una planta pequeña de hasta 12cm de alto, incluyendo la inflorescencia, puede tener de 1-3 hojas, de 6-10cmx1-2mm, basales, erectas, dísticas, membranosas, linear-lanceoladas, acuminadas, dentadas a lo largo de los márgenes y deciduas. Tiene raíces gruesas, carnosas, ligeramente aplanadas de 2-4mm de grosor. Florece entre octubre y enero y la inflorescencia es una panícula erecta de 7-8.5cm de largo, saliendo de las axilas de las hojas. Las flores son diminutas de 4.5-5mm de diámetro, con apertura escalonada, sépalos y pétalos verde púrpura, labelo púrpura oscuro (Solano et al., 2011).
Al ser T. helleri una especie recientemente registrada para México, con una población relativamente pequeña y muy localizada, se decidió estudiar su distribución en los forófitos que ocupa su estructura poblacional y su producción de frutos, para conocer más de las características y comportamiento de esta orquídea en el agroecosistema cafetalero donde se localiza.
Materiales y métodosCaracterización del sitio. El estudio se desarrolló durante el año 2009, en una plantación de café (C. arabica) de sombra de 20 años de edad, en la comunidad rural Benito Juárez El Plan (BJ), en la región del Soconusco, municipio de Cacahoatán, Soconusco, Chiapas, México. En este cafetal no se aplican agroquímicos y anualmente sólo se realizan 2 limpiezas de maleza y 1 poda ligera. La plantación se localiza aproximadamente a 1 400m snm, en los 15°5'15” N y 92°8'55” O. El promedio anual de precipitación es de 3 730mm y la temperatura promedio es de 20.7° C (García, 1988). En este cafetal se encuentra la única población conocida en México de T. helleri (Solano et al., 2011).
Toma de datos. Se realizaron transectos exploratorios, de longitud variable en dependencia de la dificultad del terreno, por todo el cafetal y varios parches de bosque cercanos, intentando localizar plantas de T. helleri; se recorrieron aproximadamente 4 ha. La búsqueda se realizó a simple vista y con binoculares cuando fue necesario. Se localizaron e identificaron todos los forófitos donde crecía T. helleri y se extendió la unidad de muestreo a 1 875m2 (0.1875 ha) de cafetal alrededor de la población de la orquídea, por considerarse esta área como la de mayor influencia sobre la población. En este trabajo el término forófito sólo se utiliza para los árboles y cafetos donde crecía T. helleri (García-González y Pérez, 2011).
Altura y DAP de los forófitos y los forófitos potenciales. Además de los forófitos identificados, se midió la altura y DAP (diámetro a la altura del pecho, a 1.3m del suelo), de todos los árboles de sombra y cafetos dentro del área de muestreo (1 875m2). La altura (m) se estimó empleando una vara recta de madera, de 4m de largo, graduada a intervalos de 50cm (García-González et al., 2011). El DAP (cm) se determinó midiendo la circunferencia (C) del tronco y luego aplicando a este valor la fórmula DAP= C / π. De forma similar se determinó el diámetro y la altura de los forófitos.
Distribución vertical. Para establecer la zonación vertical de T. helleri, los cafetos fueron divididos en tronco, horquetas, ramas, ramillas, y los árboles de sombra en tronco, horquetas, ramas, según la zonación propuesta por García-González et al. (2011), específicamente para este tipos de forófitos.
Orientación en el forófito. Mediante 4 tramos de cuerda de 2m, que fueron colocados en el suelo en forma de cruz, teniendo como punto central el tronco de los forófitos y empleando una brújula, se estimó la orientación en que se encontraba ubicada cada orquídea en el forófito: norte (N), sur (S), este (E), oeste (O), noroeste (NO), noreste (NE), sureste (SE) y suroeste (SO).
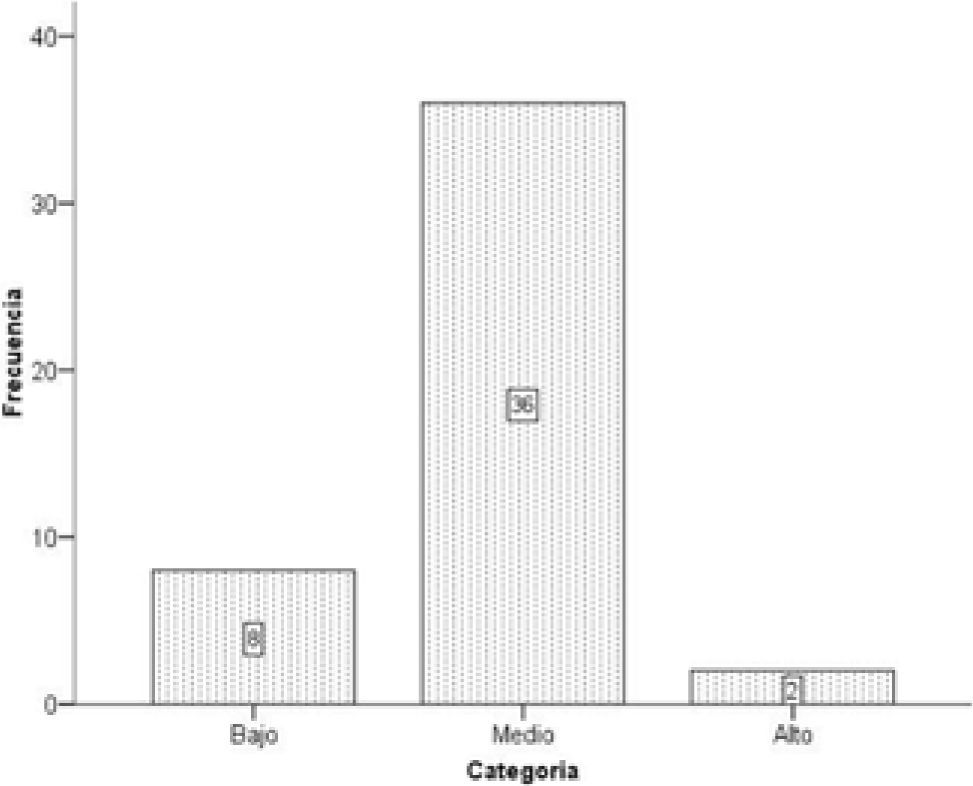
Parámetros poblacionales. Todas las plantas de orquídea se incluyeron en 2 clases de vida, las que presentaban indicios de floración actual o de años anteriores fueron consideradas como adultas y las restantes como plantas inmaduras. Se contaron y midieron las inflorescencias producidas por cada planta adulta de T. helleri. En cada inflorescencia se contó el número de flores y de frutos. Análisis estadísticos. Por el reducido número de plantas sólo se aplicó estadística descriptiva. Se utilizó la media y la desviación estándar. El programa SPSS 15.0 se empleó para hacer un gráfico de frecuencias con las categorías de altura a las que se encontraron las plantas de T. helleri en cada forófito (bajo= 0-2m, medio= 2.1-4m, alto= 4.1-6m). Los datos de altura se dividieron equitativamente en 3 categorías, teniendo en cuenta los mínimos y máximos donde se localizaron orquídeas en cada forófito.
ResultadosÁrboles de sombra y cafetos, densidad, altura y diámetro. En los 1 875m2 (0.1875 ha) estudiados se encontraron 38 árboles de sombra de 5 especies (Inga micheliana Harms., Inga lauriana (Sw.) Willd., Trema micrantha Blume, Citrus sp., Vernonia deppeana Less.). Inga micheliana fue la especie más abundante con 31 árboles. Se contabilizaron 410 cafetos (C. arabica). En el área muestreada, la densidad de árboles de sombra fue de 0.02 árboles/m2 (202.66 árboles/ha) y la de cafetos de 0.218 cafetos/m2 (2 187 cafetos/ha) en el área muestreada. La densidad general fue de 0.238 plantas/m2 (2 380 plantas/ha) teniendo en cuenta árboles y cafetos. La altura promedio de los cafetos fue 3.04±1.14m y la de los árboles de sombra 6.24±2.03m. El valor promedio de DAP de los cafetos fue 2.99±1.88cm y el de los árboles de sombra 20±12.59cm. Forófitos, altura y diámetro donde crece T. helleri. Sólo se localizó a T. helleri en 2 árboles de I. micheliana y en 1 cafeto (Cuadro 1), todos dentro de un área aproximada de 50m2. Sobre I. micheliana estuvo 78.26% de la población de T. helleri y 21.74% sobre C. arabica.
Forófitos encontrados, altura y diámetro a la altura del pecho (DAP) de cada uno, número de plantas de Telipogon helleri en cada forófito, distribución vertical de las orquídeas en cada forófito (tronco, horquetas, ramas, ramillas) y clase de vida de cada planta de orquídea (I= inmaduras; A= adultas)
| Forófito | Especie | Altura (m) | DAP (cm) | Núm. de Orquídeas | Tronco | Horquetas | Ramas | Ramillas | Orquídeas por clase de vida | |
|---|---|---|---|---|---|---|---|---|---|---|
| I | A | |||||||||
| 1 | Inga micheliana | 7.9 | 26.73 | .1 | 1 | 0 | 0 | — | 0 | 1 |
| 2 | Inga micheliana | 8 | 30.87 | 35 | 32 | 0 | 3 | — | 0 | 35 |
| 3 | Coffea arabica | 4.2 | 4.13 | 10 | 1 | 0 | 9 | 0 | 1 | 9 |
| Total | — | — | — | 46 | 34 | 0 | 12 | 0 | 1 | 45 |
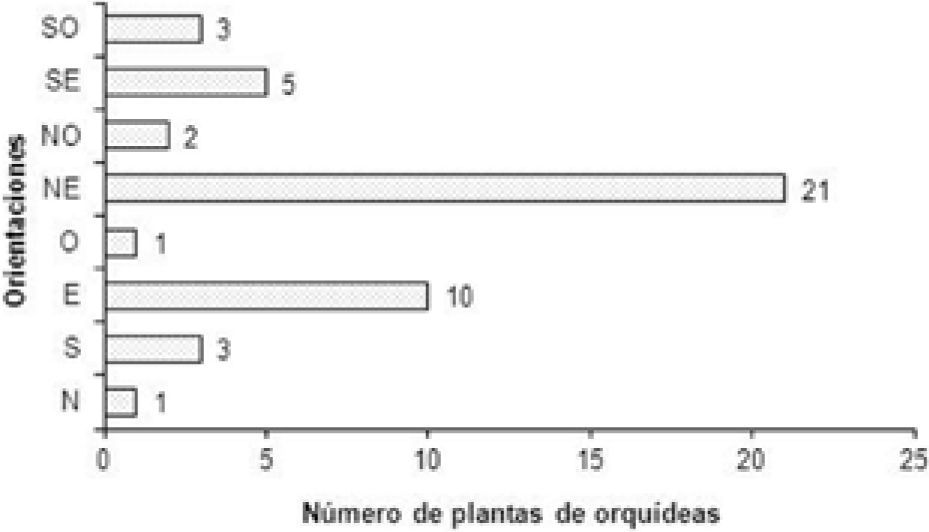
Distribución vertical, orientación en el forófito y parámetrospoblacionales. En el tronco se localizó 73.92% de las orquídeas, 26.08% en las ramas y no se encontraron plantas ocupando las horquetas ni las ramillas (Cuadro 1). El mayor número de orquídeas se encontró en la categoría medio de altura (78.26%) (Fig. 1). Se aprecia una tendencia de T. helleri a crecer en el lado noreste de los forófitos que ocupa, 45.67% de la población, (Fig. 2). La mayor parte de la población (97.83%) fueron plantas adultas (Cuadro 1). Del total de plantas adultas de T. helleri (Cuadros 1 y 2), 97.78% (44 plantas) florecieron. El número promedio de inflorescencias por planta adulta fue de 1.13±0.40, con un largo promedio de 5.79±3.24cm (Cuadro 2). El número promedio de flores por planta fue 4.02±2.79 y el número promedio de frutos por planta 0.95±1.04 (Cuadro 2). De las flores producidas 23.76% formaron frutos.
 y número de plantas en cada categoría.")
.")
Forófitos encontrados, orquídeas adultas (Telipogon helleri) en cada forófito, número total de espigas, largo promedio de estas espigas, número total de flores y número total de frutos
| Forófito | Especie | Orquídeas adultas | Número de espigas | Número promedio de espigas por planta | Largo promedio de espigas (cm) | Número de flores | Número promedio de flores por planta | Número de frutos | Número promedio de frutos por planta |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Inga micheliana | 1 | 1 | — | 12 | 6 | — | 3 | — |
| 2 | Inga micheliana | 35 | 42 | 1.2±0.44 | 5.34±2.93 | 141 | 4.02±2.91 | 25 | 0.71±0.92 |
| 3 | Coffea arabica | 9 | 8 | 0.88±0 | 7.35±4.04 | 34 | 3.77±2.45 | 15 | 1.66±1.08 |
| Total | — | 45 | 51 | — | — | 181 | — | 43 | — |
Densidad, arquitectura de árboles de sombra y cafetos, y el cafetal como hábitat. La densidad, arquitectura, altura y DAP de los forófitos actuales y potenciales, crea variaciones en las condiciones de temperatura y humedad que a su vez afecta a la germinación y el establecimiento de epífitas (Benzing, 1990). La combinación de estos aspectos puede tener un efecto sustancial sobre la penetración de la luz, la circulación de aire y la superficie disponible para el establecimiento de epífitas. Esas mismas variables también influyen en la abundancia y diversidad de las bacterias, hongos micorrícicos y polinizadores que forman interacciones estrechas con las orquídeas (Bertolini y Damon, 2011; García-González et al., 2011; Pérez-Hernández et al., 2011; Damon et al., 2012; García-González et al., 2013). La densidad de árboles de sombra en BJ contrasta con la densidad de los árboles en el bosque nublado original que es de aproximadamente 638 árboles/ha, con una altura máxima de aproximadamente 22m (Williams-Linera y López-Gómez, 2008).
Aparentemente, según sus requerimientos ecológicos, T. helleri encontró condiciones favorables para establecerse en el cafetal, el cual puede ser un refugio temporal para la población y es de esperarse que las semillas migren en el futuro a los parches cercanos de bosque nublado, con elementos de la selva perennifolia, o es posible que la especie se adapte definitivamente a las condiciones del agroecosistema cafetalero, logrando éxito en la atracción de polinizadores y la dispersión de semillas en esta vegetación menos densa y de dosel menos cerrado que la del bosque nublado original. Será necesario monitorear el comportamiento de esta población en el futuro para dar respuesta a estas interrogantes. Oncidium poikilostalix (Kraenzl.) M. W. Chase y N. H. Williams, otra especie de orquídea recién descubierta en la región del Soconusco en 2008, aparentemente también prefiere el agroecosistema cafetalero y está logrando altas tasas de reproducción en este hábitat (García-González et al., 2011, 2013).
Teniendo en cuenta que las orquídeas epífitas pueden utilizarse como indicadores de la integridad ecológica de las comunidades vegetales en las que crecen (Williams-Linera et al. 1995), el hallazgo de T. helleri en un cafetal de sombra reafirma el papel de estos hábitats como refugios y corredores para la flora de los bosques originales que sustituyeron (Perfecto y Snelling, 1995; Moguel y Toledo, 1999; Soto-Pinto et al., 2001; Bartra et al., 2002). Probablemente estos cafetales ofrecen nichos no tan frecuentes dentro del bosque conservado y menos aún en los parches de selva circundante, con un marcado efecto de borde y donde la estructura natural de la vegetación y los árboles más altos y viejos, con condiciones más óptimas para el establecimiento de epífitas, se han perdido en muchos casos.
Los cafetales de sombra son refugio para muchas orquídeas raras o ausentes en otros hábitats, como lo plantea Solano et al. (2011) para los casos de T. helleri, O. poikilostalix y Plectrophora alata (Rolfe) Garay. Específicamente en la región Tacaná-Boquerón, para algunos grupos aún es posible encontrar una alta diversidad biológica en las plantaciones de cafetales tradicionales, que son los principales reservorios para importantes componentes de la vegetación original (Solis-Montero et al., 2005; Damon, 2011).
Forófitos, altura y diámetro donde crece T. helleri. La mayor parte de los cafetos y árboles de sombra del área, a pesar de tener similar porte, edad y estar sometidos a las mismas condiciones ambientales y de manejo, no estaban ocupados por T. helleri. Esto puede estar asociado a la presencia en los forófitos del hongo micorrícico con el que se asocia T. helleri para germinar y desarrollarse, aunque esta afirmación requiere estudios más específicos. También puede estar ocasionado por circunstancias aleatorias en la dispersión de las semillas de T. helleri, siendo factible que en el futuro próximo otros cafetos y árboles de sombra sean colonizados.
Distribución vertical, orientación en el forófito y parámetros poblacionales. Diferentes autores plantean que los patrones de estratificación vertical de las epífitas vasculares en un forófito están relacionados con sus adaptaciones ecofisiológicas y los niveles de tolerancia a las condiciones de luz y humedad, debido al gradiente vertical de estos elementos dentro del bosque (Johansson, 1974; Benzing, 1990; Hernández-Rosas, 2000; Shaw, 2004; Kromer et al., 2007; Martínez-Meléndez et al., 2008). En el caso particular de T. helleri, la ausencia de orquídeas en las horquetas y las ramillas y su presencia sólo en el tronco (Cuadro 1), a alturas entre 2.1-4m, categoría medio (Fig. 1), sugiere que esta especie se desarrolla mejor en microhábitats donde no recibe el sol directamente y dispone de luz filtrada, corrientes de aire atenuadas y mayor humedad que en el dosel superior.
En estudios análogos desarrollados en cafetales de sombra en el Soconusco con O. poikilostalix, esta especie demostró preferencia por las ramas, aunque también era abundante en las ramillas (García-González et al., 2011). En este caso, la mayor abundancia en las ramas puede ser consecuencia de las prácticas agroculturales que se le hacen a la plantación de café (C. arabica), donde el extremo de las ramas se poda periódicamente, perdiéndose gran parte de las ramillas, eliminando con ellas las plantas que las ocupaban y limitando la disponibilidad de este microhábitat (García-González et al., 2011).
La aparente tendencia de T. helleri a crecer preferentemente con orientación noreste (Fig. 2) requiere estudios a largo plazo para determinar la estacionalidad y dirección predominante de los vientos en el área, especialmente los que traen las lluvias, y su posible influencia en la dispersión y germinación de las semillas, seguido por el buen desarrollo y persistencia de las plántulas de esta especie de orquídea.
El número de plantas adultas de T. helleri (Cuadros 1, 2) contribuye en gran medida a la reproducción y expansión de la especie, al ser la etapa adulta la más importante en la vida de las orquídeas (Zotz, 1998; Winkler y Hietz, 2001; Mondragón, 2009). Sin embargo, el bajo número de orquídeas inmaduras (Cuadro 1) puede indicar poca viabilidad de semillas, poco éxito de germinación o bajo éxito en la supervivencia de plántulas, lo cual haría vulnerable a la especie.
En comparación con el tamaño general de la planta, T. helleri produce inflorescencias relativamente grandes y con abundantes flores y frutos (Cuadro 2). Damon y Valle-Mora (2008) reportan porcentajes de polinización de 1.23% en Notylia barkeri Lindl. y 1.48% en Erycina crista-galli (Rchb.f.) N. H. Williams y M. W. Chase, orquídeas que crecen en plantaciones de café, también en el Soconusco, mientras Pérez-Hernández et al. (2011) reportaron valores de 4.9 y 4.4%, respectivamente, para las mismas especies de orquídeas y en el mismo sitio. En la polinización de estas 2 especies de orquídea la producción de frutos es densodependiente. No obstante, la producción de frutos en las orquídeas puede estar limitada por una variedad de condiciones que no tienen que ser mutuamente excluyentes, ni inmediatamente evidentes, como pueden ser la ausencia de polinizadores, la herbivoría o la limitación de recursos en la planta madre (Ackerman y Montalvo, 1990).
Actualmente en la región del Soconusco varias especies de orquídeas no producen frutos en todos o la mayoría de los sitios donde se encuentran, sugiriendo poblaciones sin reproducción sexual (Damon, 2011; Damon et al., 2012). De acuerdo con los datos presentados por Neiland y Wilcock (1998), el porcentaje de frutos producidos por T. helleri corresponde a orquídeas no autógamas, que ofrecen recompensa, y es similar al de otras especies de orquídeas que se pueden encontrar en cafetales (Solis-Montero et al., 2005). Esto sugiere que T. helleri en el cafetal estudiado encuentra polinizador(es), que pueden haber emigrado con ella o alguno de los insectos autóctonos logra realizar la polinización.
La mayoría de las plantaciones de café en la región de Soconusco son administradas por pequeños productores, que por razones culturales y económicas realizan un laboreo ligero en sus campos, ya que no se aplican agroquímicos y anualmente sólo se realizan 2 limpiezas de maleza y una poda ligera, lo que favorece la estabilidad y la persistencia de las epífitas. En estas condiciones, T. helleri paulatinamente podría ir ampliando su distribución en el área, aunque debido a los problemas de reclutamiento que aparentemente presenta y al pequeño tamaño y lo localizado de la población, podría ser vulnerable a la transformación de su hábitat por cambios en las prácticas de manejo, extracción de madera o un cambio del uso del suelo. Esto hace imprescindible monitorear la población para ver su comportamiento a largo plazo, y sería importante evaluar su relación con las poblaciones de las demás especies de epífitas vasculares presentes en el agroecosistema cafetalero.
Al Conacyt, por financiar este estudio mediante el proyecto: Diversidad y conservación de las orquídeas del Corredor Biológico Tacana-Boqueron, Conacyt-Fondos Mixtos-Chiapas, CHIS-2006-206-45802. A Nelson Pérez Miguel, de la comunidad Benito Juárez El Plan, por permitirnos el acceso a su plantación. A la organización Idea Wild por el equipamiento proporcionado, fundamental para la elaboración de este trabajo.