




En los meses de diciembre y febrero del 2010-2011 se realizaron estudios en el complejo lacustre de Sardinayacu, localizado en el Parque Nacional Sangay, en los cuales se trabajó 10 días. El objetivo de este estudio fue analizar y describir bioacústicamente las vocalizaciones de los anuros en la laguna Cormorán (una de las lagunas del complejo lacustre de Sardinayacu). Para el estudio se realizaron recorridos diurnos y nocturnos, enfocando el esfuerzo de muestreo a los puntos de mayor actividad vocal, donde se grabó dicha actividad utilizando una grabadora digital, conectada a un micrófono unidireccional. Se analizaron las vocalizaciones de 6 especies de anuros, describiendo los mismos de manera cualitativa y cuantitativa a partir de sus variables espectrales (frecuencias medidas en kilohertzios) y temporales (duración e intervalos de los cantos, notas y pulsos medidos en milisegundos). Las variaciones interespecíficas indican que los cantos de las 6 especies de anuros de la laguna Cormorán presentan características temporales y espectrales propias.
In December and February, 2010-2011, studies were conducted in the Sardinayacu Lake Complex, located in Sangay National Park. Each field study had a duration of 10 days. The objective of this study was to analyze the bioacoustic characteristics of anuran vocalizations in Cormorán Lagoon, one of the lakes of the Complejo Lacustre de Sardinayacu. Day and night rounds were performed for the study, focusing the sampling effort on the areas with the greatest vocal activity. This activity was recorded with a digital recorder connected to a unidirectional microphone. The vocalizations of 6 species of anurans were analyzed, which were described both qualitatively and quantitatively based on spectral variables (frequencies measured in kilohertz) and time variables (duration and interval of the calls, notes and pulses measured in milliseconds). The interspecific variations indicate that the calls of 6 anuran species from the Cormoran Lagoon have their own time and spectral characteristics.
Una de las ciencias encargadas del estudio de la emisión, grabación y análisis de los sistemas comunicativos que presentan los animales es la bioacústica (Angulo, 2006); sus estudios han sido fundamentales para esclarecer y comprender los mecanismos de comunicación que utilizan las especies al emitir sonidos y de igual manera brindar alternativas para su conservación (Tubaro, 1999). En herpetología, el desarrollo de los estudios bioacústicos ha sido relevante, en particular los realizados en anuros, en los cuales uno de los objetivos principales es esclarecer el significado de los cantos de los mismos (Narins, Feng, Fay y Popper, 2007).
Una de las características de mayor distinción en los anuros es su vocalización, misma que se encuentra agrupada en diferentes tipos de cantos y conforman un sistema complejo de comportamiento y comunicación (Cocroft y Ryan, 1995; Dapper, Baugh y Ryan, 2011), definiendo los cantos como una unidad funcional de las vocalizaciones (Toledo et al., 2015). Uno de los cantos de mayor importancia es aquel que emite el macho para atraer a la hembra (selección sexual), ya que esta será quien elija al macho reproductor atraída por su canto (Gerhardt y Huber, 2002), de tal manera que uno de los comportamientos más importantes para la permanencia y perpetuación de las especies de anuros es la vocalización (Duellman y Trueb, 1994).
En un grado de especificidad los anuros llegan a especializar sus cantos, de tal manera que en determinadas estaciones marcan su reproducción con cantos que son únicos para dichas estaciones (Baugh y Ryan, 2010). Pueden responder a un carácter de comportamiento que varía entre las especies, como los de combate (Bolívar, Grant y Osorio, 1999), teniendo así una variedad que difiere de acuerdo con su contexto social (Wells, 2007).
Acústicamente los cantos tienen sus propias características y presentan una gran variación intra- e interespecífica en notas, frecuencias y duraciones (Almendáriz y Batallas, 2012); en cuanto a cómo se escuchan, generan cantos de tipo trino, goteo, silbidos, pulsados y compuestos (Bernal, Montealegre y Páez, 2004).
En Ecuador los estudios que contienen análisis de cantos de anuros se han enfocado al complemento de la información de nuevas especies, estudios ecológicos e historia natural (e.g., Brito, Almendáriz y Batallas, 2014; Coloma et al., 2012; Hutter y Guayasamin, 2012; Páez-Vacas, Coloma y Santos, 2010; Yánez-Muñoz, Bejarano-Muñoz, Brito y Batallas, 2014), habiendo pocos estudios focalizados a las características acústicas del canto de los anuros (e.g., Batallas y Brito, 2014a; Dautel et al., 2011; Guerra y Ron, 2008; Hutter et al., 2013; Ron, 2008; Sinsch y Juraske, 2006). El conocimiento de las vocalizaciones ha permitido identificar especies sin la necesidad de colectar o sacrificar (Angulo, 2006) y realizar comparaciones taxonómicas relacionando sus evidencias evolutivas (Peloso et al., 2014). En el presente estudio analizamos y describimos las vocalizaciones de 6 especies de anuros de la laguna Cormorán, aportando con sus valores espectrales y temporales valores de gran importancia que permitirán en estudios futuros establecer diferencias intra- e interespecíficas entre las especies analizadas.
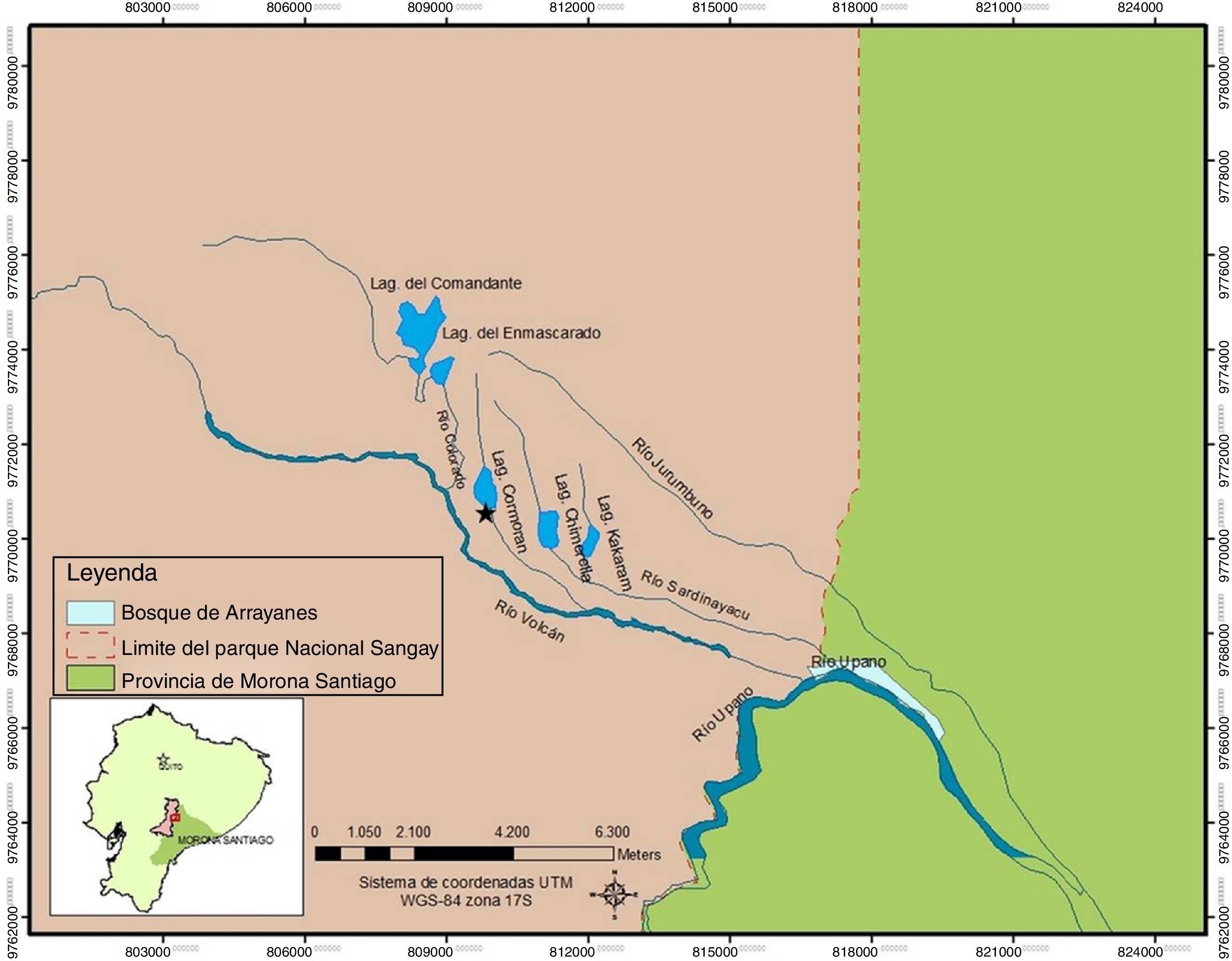
Materiales y métodosEl área de estudio se localiza en la laguna Cormorán, del complejo lacustre de Sardinayacu, al centro oriente del Parque Nacional Sangay, Ecuador (2°04'20.59"S, 78°12'52.40"O; 1,750m snm) (fig. 1). Políticamente corresponde a la provincia de Morona Santiago, cantón Morona, parroquia Sinaí. El área pertenece al piso zoogeográfico Subtropical Oriental (Albuja et al., 2012) y a la formación vegetal Herbazal Lacustre Montano Bajo del Sur de la Cordillera Oriental de los Andes (Cerón, 2013).

La fase de campo se desarrolló en los meses de diciembre 2010-febrero 2011, con 10 días de trabajo por mes. Se utilizó el muestreo ad libitum (Lehner, 1996), estableciendo 3 puntos permanentes de grabación, los mismos que se ubicaron en la zona litoral de la laguna, en el bosque de ribera y en el efluente de la misma (fig. 2). El esfuerzo de muestreo fue de 10:00h a 13:00h y de 19:00h a 22:00h siendo la actividad vocal la que marcó el esfuerzo de muestreo, grabando un mínimo de 2 individuos y colectando un máximo de 3 especímenes de referencia (voucher) por especie (apéndice1); los sitios de muestro se focalizaron en entornos libres de saturación de frecuencia y ruidos excesivos, teniendo muestras con una calidad aceptable para el análisis; en algunos casos la muestra fue baja (aunque representativa), debido a las difíciles condiciones de grabación. Los cantos se registraron con una grabadora digital Olympus WS-802, a una frecuencia de muestreo de 44.1kHz y 16 «bits» de resolución, conectada a un sistema modular Sennheiser K6-C, acoplado a un micrófono de cabezal Sennheiser (ME 66) 40-20,000Hz±2.5dB.
 Zona litoral de la laguna Cormorán; (B) bosque de ribera; (C) efluente de la laguna Cormorán.")
Una vez realizada la grabación se procedió a la colecta manual de los individuos, siendo transportados en recipientes plásticos al campamento base para su registro fotográfico y preservación. Los especímenes fueron sacrificados según las recomendaciones de Chen y Combs (1999) y preservados según los protocolos de Simmons (2002). Los animales colectados y los cantos grabados se encuentran depositados en la sección de herpetología del Museo de Historia Natural «Gustavo Orces» de la Escuela Politécnica Nacional (MEPN).
Para realizar la digitalización, los filtros de ruido y los análisis temporales, se utilizó el programa Adobe Audition CS6 (Almendáriz y Batallas, 2012), a su vez, para las diagramaciones y análisis espectrales el programa Raven 1.4 (Charif, Waack y Strickman, 2010), utilizando la ventana de Hann al 80% de superposición y 512 puntos de resolución de la transformación rápida de Fourier (FFT).
Los parámetros que se analizaron fueron: 1) frecuencia dominante: frecuencia de mayor energía medida a lo largo de todo el canto y en todos sus componentes; 2) cantos/minuto: tasa de repetición de llamados en el lapso de un minuto; 3) notas/canto: número de unidades acústicas de un determinado patrón de amplitud reconocido en los cantos; 4) pulsos/nota: número de subunidades acústicas de un llamado, reconocido en las notas; 5) duración del canto: tiempo desde el inicio hasta el final de un llamado, medido con el analizador de forma de onda; 6) duración de las notas; 7) duración de los pulsos; 8) intervalos entre cantos: tiempo transcurrido entre llamado y llamado; 9) intervalos entre notas. Las definiciones y mediciones realizadas en los parámetros acústicos se siguieron según los términos de Batallas y Brito (2014a), Cocroft y Ryan (1995), Díaz y Cádiz (2007) y Duellman y Pyles (1983).
Con las variables analizadas se calcularon medidas de tendencia central y de dispersión (máximo, mínimo, media y desviación estándar), utilizando los diagramas de cajas y bigotes (box-plot) para la comparación de estas medidas.
ResultadosCaracterización de los cantosEn la laguna Cormorán se registraron vocalizaciones de 6 especies de anuros (fig. 3), analizando un total de 225 cantos, distribuidos en 19 grabaciones, teniendo un mínimo de 2 grabaciones específicas por especie.
 Chimerella mariaelenae, MEPN 14519, LRC 20.2mm; (B) Noblella personina, MEPN 14324, LRC 16.3mm; (C) Pristimantis petersi, MEPN 14413, LRC 19.7mm; (D) Hypsiboas almendarizae, MEPN 14699, LRC 35.4mm; (E) Hypsiboas cinerascens, MEPN 14465, LRC 36.8mm; (F) Osteocephalus cf. verruciger, MEPN 14479, LRC 48.8mm. Fotografías: J. Brito.")
Especímenes machos adultos colectados en la laguna Cormorán. (A) Chimerella mariaelenae, MEPN 14519, LRC 20.2mm; (B) Noblella personina, MEPN 14324, LRC 16.3mm; (C) Pristimantis petersi, MEPN 14413, LRC 19.7mm; (D) Hypsiboas almendarizae, MEPN 14699, LRC 35.4mm; (E) Hypsiboas cinerascens, MEPN 14465, LRC 36.8mm; (F) Osteocephalus cf. verruciger, MEPN 14479, LRC 48.8mm. Fotografías: J. Brito.
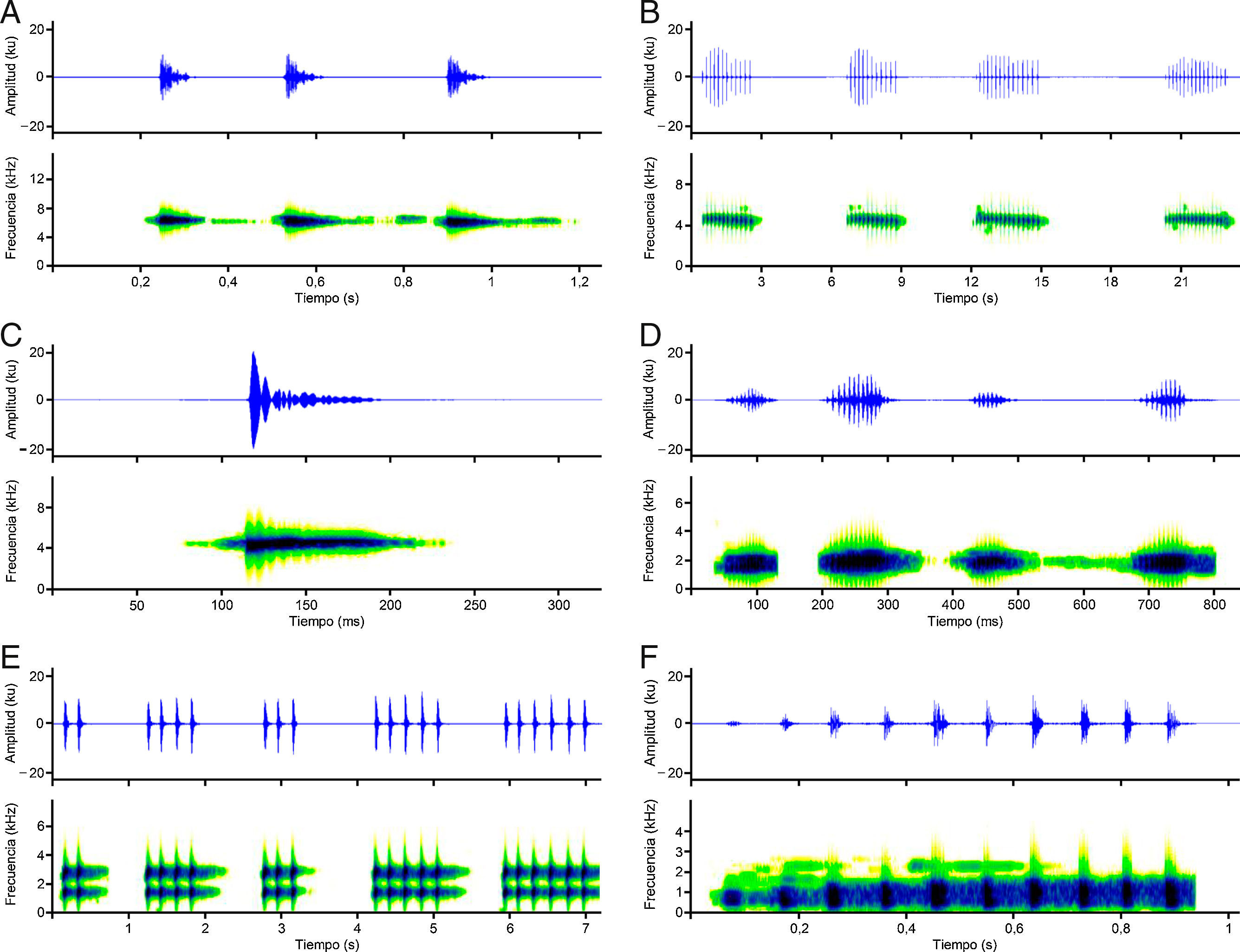
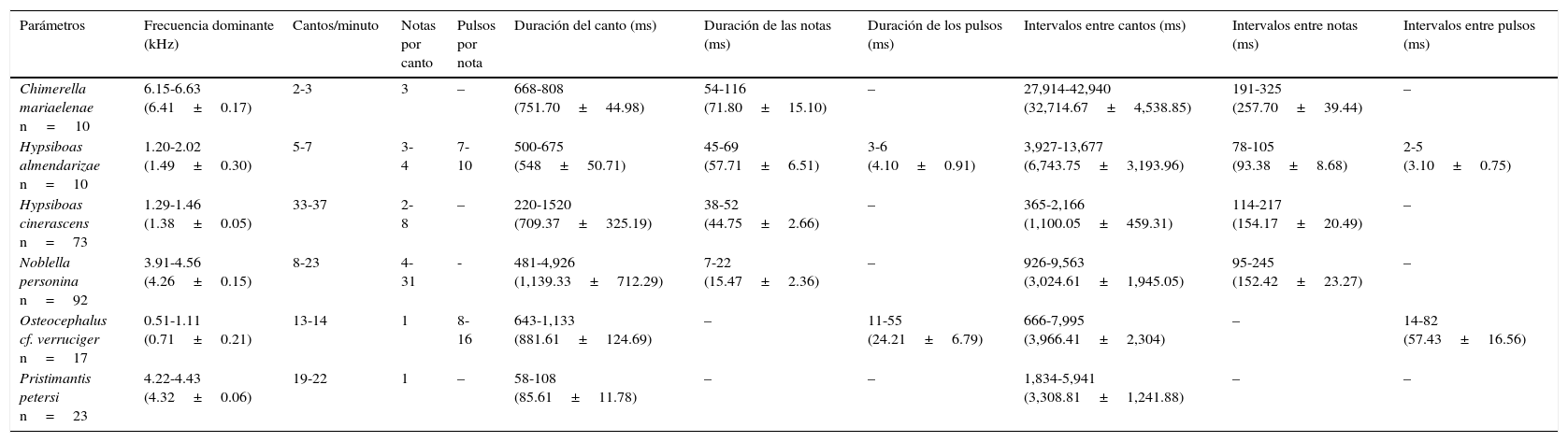
Chimerella mariaelenae (Cisneros-Heredia y McDiarmid, 2006). Presenta cantos de frecuencia modulada, cuya frecuencia dominante es de=6.41±0.17kilohertzios (kHz) (fig. 4A); tienen una duración de=751.70±44.98milisegundos (ms), con intervalos de=32,714.67±4,538.85ms, emitiendo de 2-3 cantos/minuto. Están conformados por 3 notas, las mismas que presentan una duración de=71.80±15.10ms, con intervalos de=257.70±39.44ms (tabla 1).
 Chimerella mariaelenae; (B) Noblella personina; (C) Pristimantis petersi; (D) Hypsiboas almendarizae; (E) Hypsiboas cinerascens; (F) Osteocephalus cf. verruciger.")
Estadística descriptiva utilizada en las vocalizaciones de los anuros de la laguna Cormorán. De los valores calculados se detalla la mínima, máxima; promedio y desviación estándar entre paréntesis. 512 puntos FFT.
| Parámetros | Frecuencia dominante (kHz) | Cantos/minuto | Notas por canto | Pulsos por nota | Duración del canto (ms) | Duración de las notas (ms) | Duración de los pulsos (ms) | Intervalos entre cantos (ms) | Intervalos entre notas (ms) | Intervalos entre pulsos (ms) |
|---|---|---|---|---|---|---|---|---|---|---|
| Chimerella mariaelenae n=10 | 6.15-6.63 (6.41±0.17) | 2-3 | 3 | – | 668-808 (751.70±44.98) | 54-116 (71.80±15.10) | – | 27,914-42,940 (32,714.67±4,538.85) | 191-325 (257.70±39.44) | – |
| Hypsiboas almendarizae n=10 | 1.20-2.02 (1.49±0.30) | 5-7 | 3-4 | 7-10 | 500-675 (548±50.71) | 45-69 (57.71±6.51) | 3-6 (4.10±0.91) | 3,927-13,677 (6,743.75±3,193.96) | 78-105 (93.38±8.68) | 2-5 (3.10±0.75) |
| Hypsiboas cinerascens n=73 | 1.29-1.46 (1.38±0.05) | 33-37 | 2-8 | – | 220-1520 (709.37±325.19) | 38-52 (44.75±2.66) | – | 365-2,166 (1,100.05±459.31) | 114-217 (154.17±20.49) | – |
| Noblella personina n=92 | 3.91-4.56 (4.26±0.15) | 8-23 | 4-31 | - | 481-4,926 (1,139.33±712.29) | 7-22 (15.47±2.36) | – | 926-9,563 (3,024.61±1,945.05) | 95-245 (152.42±23.27) | – |
| Osteocephalus cf. verruciger n=17 | 0.51-1.11 (0.71±0.21) | 13-14 | 1 | 8-16 | 643-1,133 (881.61±124.69) | – | 11-55 (24.21±6.79) | 666-7,995 (3,966.41±2,304) | – | 14-82 (57.43±16.56) |
| Pristimantis petersi n=23 | 4.22-4.43 (4.32±0.06) | 19-22 | 1 | – | 58-108 (85.61±11.78) | – | – | 1,834-5,941 (3,308.81±1,241.88) | – | – |
kHz: kilohertzios; ms: milisegundos; n: número de cantos analizados.
Especie de actividades nocturnas; sus vocalizaciones son muy parecidas a trinos, emitiendo desde vegetación arbustiva y arbórea. En la laguna Cormorán forman grandes agrupaciones alrededor del perfil litoral de la laguna, vocalizando constantemente coros ordenados, sin enmascaramiento o sobreposición; una de las características peculiares del coro es la de ser activados a partir de uno o varios machos dominantes, donde cada individuo emite las notas características de la especie.
Noblella personina (Harvey, Almendáriz, Brito y Batallas, 2013). Presenta cantos de frecuencia modulada, cuya frecuencia dominante es de=4.26±0.15kHz (fig. 4B); tienen una duración de=1,139.33±712.29ms, con intervalos de=3,024.61±1,945.05ms, emitiendo de 8-23 cantos/minuto. Están conformados por 4-31 notas, las mismas que presentan una duración de=15.47±2.36ms, con intervalos de=152.42±23.27ms (tabla 1).
Especie de actividad diurna y nocturna; sus vocalizaciones son parecidas a golpeteos metálicos, emitiendo desde la hojarasca e interior de las raíces aéreas de algunos árboles; no forma un coro permanente y sus patrones de vocalización suelen verse modificados, generando distintos tipos de cantos en respuestas etológicas específicas.
Pristimantis petersi (Lynch y Duellman, 1980). Presenta cantos con frecuencia modulada, cuya frecuencia dominante es de=4.32±0.06kHz (fig. 4C); tienen una duración de=85.61±11.78ms con intervalos de=3,308.81±1,241.88ms, emitiendo de 19-22 cantos/minuto. Están conformados por una nota (tabla 1).
Especie de actividad nocturna; sus vocalizaciones son parecidas a un «tic» metálico muy característico de ciertos Pristimantis, como los del grupo P. lacrimosus (Batallas y Brito, 2014b), emitiendo desde arbustos y árboles; sus coros carecen de orden específico, intensificando su actividad en lluvias ligeras, generando silencios prolongados en particular cuando canta otra especie.
Hypsiboas almendarizae (Caminer y Ron, 2014). Presenta cantos de frecuencia modulada, cuya frecuencia dominante es de=1.49±0.30kHz (fig. 4D); tienen una duración de=548±50.71ms, con intervalos de=6,743.75±3,193.96ms, emitiendo de 5-7 cantos/minuto. Están conformados por 3-4 notas, con una duración de=57.71±6.51ms, con intervalos de=93.38±8.68ms. Las notas presentan 7-10 pulsos; con una duración de=4.10±0.91ms, con intervalos de=3.10±0.75ms (tabla 1).
Especie de actividad nocturna; sus vocalizaciones son parecidas a graznidos de patos, emitiendo desde la vegetación arbustiva, vegetación emergente en la zona litoral de la laguna, o en las cercanías de cuerpos de agua. No conforman un coro y pueden presentar gran actividad vocal, como también prolongados lapsos de silencio con la emisión esporádica de cantos.
Hypsiboas cinerascens (Spix, 1824). Presenta cantos de frecuencia modulada, cuya frecuencia dominante es de=1.38±0.05kHz, emitiendo un armónico (fig. 4E): tienen una duración de=709.37±325.19ms, con intervalos de=1,100.05±459.31ms, emitiendo de 33-37 cantos/minuto. Están conformados por 2-8 notas, las mismas que presentan una duración de=44.75±2.66ms, con intervalos de=154.17±20.49ms (tabla 1).
Especie de actividad nocturna, emitiendo sus vocalizaciones desde la vegetación cercana a cuerpos de agua. Presenta coros de gran intensidad, sin un orden específico y con sobreposición de cantos.
Osteocephalus cf. verruciger. Presenta cantos de frecuencia modulada, cuya frecuencia dominante es de=0.71±0.21kHz (fig. 4F); tienen una duración de=881.61±124.69ms, con intervalos de=3,966.41±2,304ms, emitiendo de 13-14 cantos/minuto. Están conformados por una nota. Las notas presentan 8-16 pulsos; con una duración de=24.21±6.79ms, con intervalos de=57.43±16.56ms (tabla 1).
Especie de actividad nocturna; sus vocalizaciones las emite desde vegetación arbustiva cercana a cuerpos de agua y sumergidos en el interior de pozas temporales; es una especie de poca actividad vocal a excepción de cuando lo hace en coros alrededor de una hembra; los mismos que son de gran intensidad, sin un orden específico y con sobre posición de cantos.
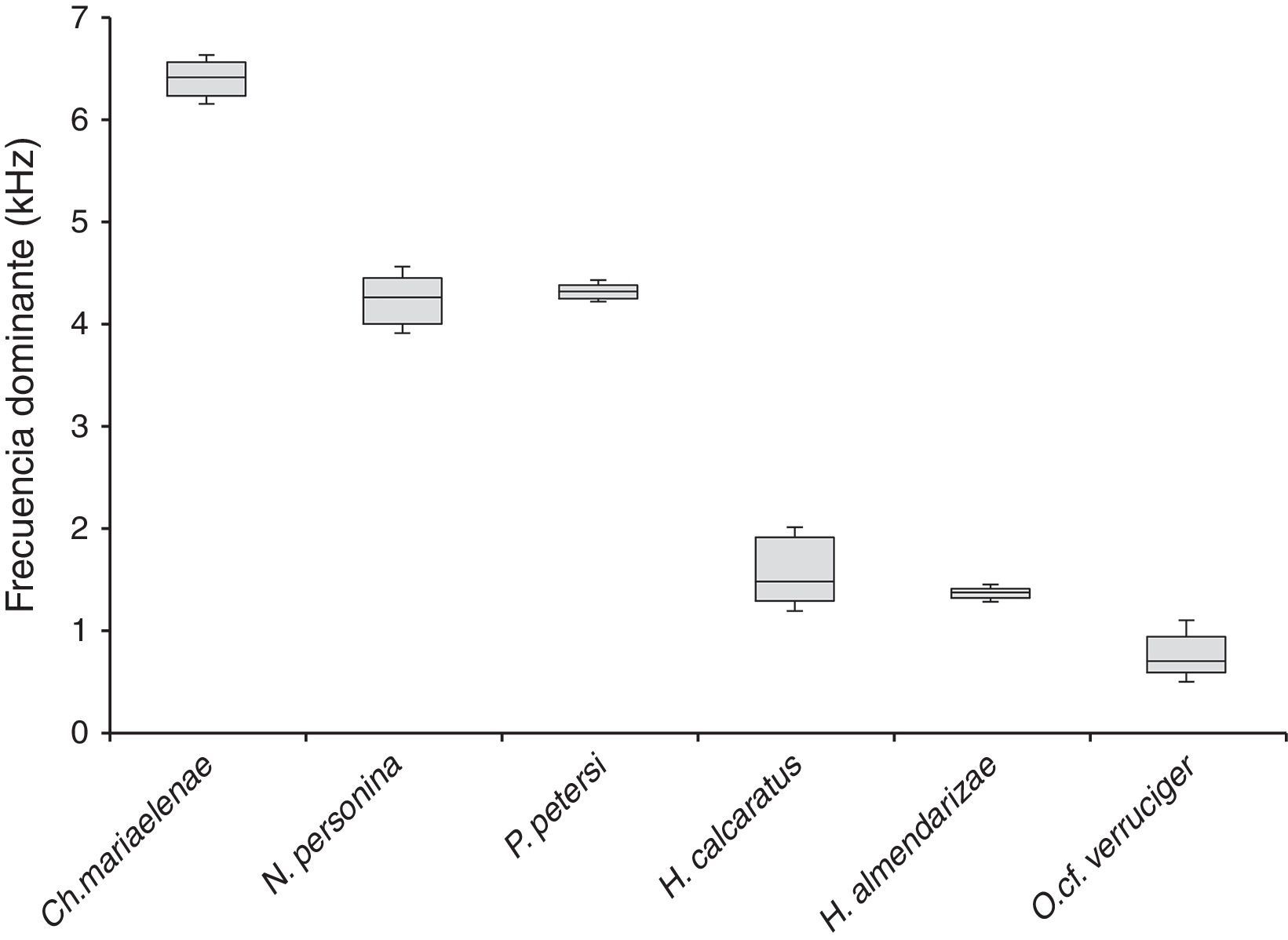
Comparación intraespecíficaChimerella mariaelenae presentó los valores más altos para la frecuencia dominante (=6.41kHz); en valores intermedios se encuentra P. petersi (=4.32kHz) y N. personina (=4.26kHz) y con frecuencias más bajas H. almendarizae (=1.49kHz), H. cinerascens (=1.38kHz) y O. cf. verruciger (=0.71kHz) (fig. 5).

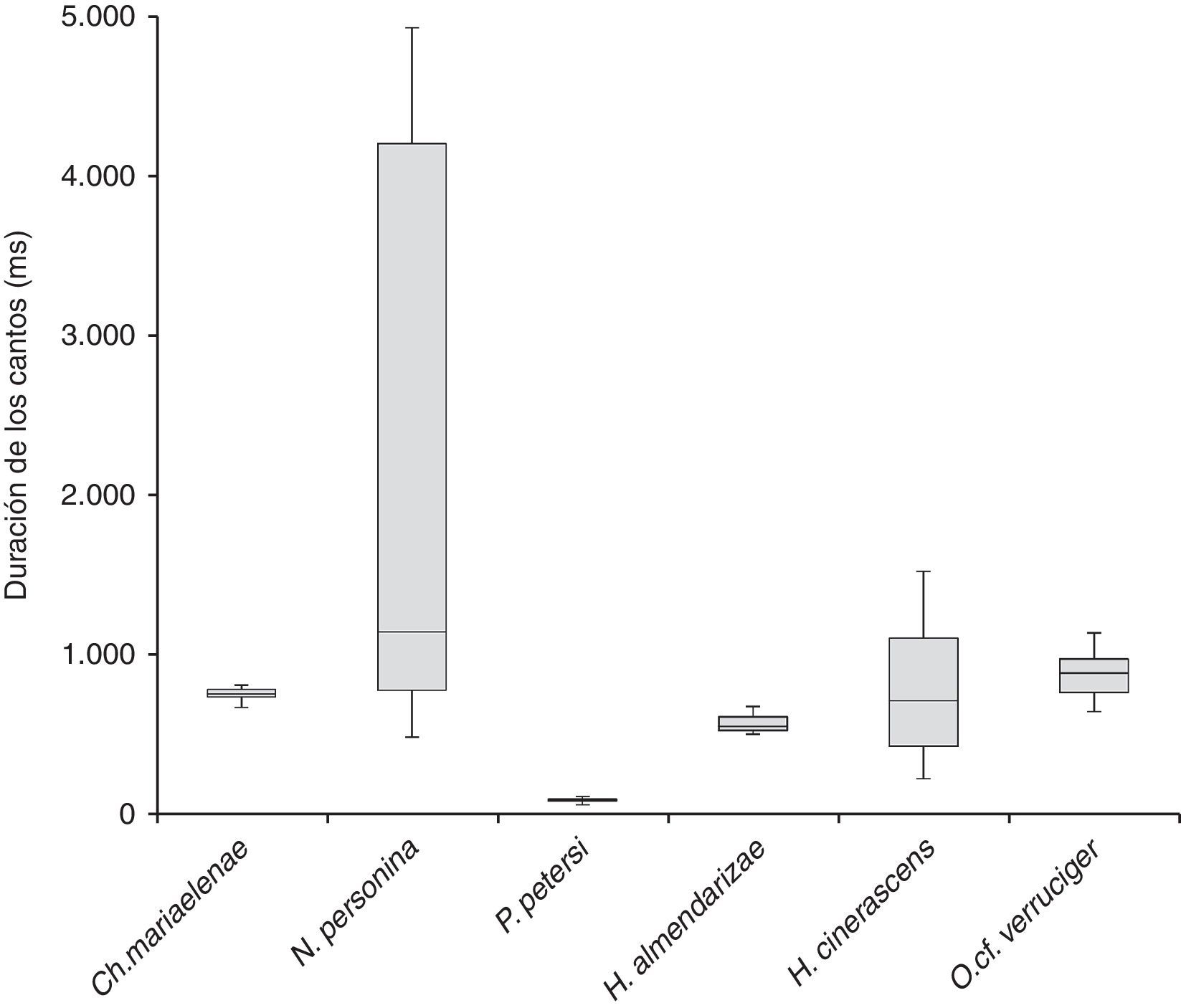
En las variables temporales N. personina presentó cantos de mayor duración de hasta 4,926ms, seguido de H. cinerascens con cantos de hasta 1,520ms y O. cf. verruciger de hasta 1,133ms; Ch. mariaelenae y H. almendarizae no alcanzan los 1,000ms de duración. Los cantos de menor duración se registraron en P. petersi de hasta 108ms (fig. 6).

En cuanto a las características de los cantos, fueron emitidos en series de notas como en N. personina con un rango de 4-31 notas y H. cinerascens con un rango de 2-8 notas; emitiendo notas constantes, sin variaciones muy grandes en su número tenemos a Ch. mariaelenae con 3 notas y H. almendarizae con 3-4 notas. Osteocephalus cf. verruciger y P. petersi generaron una nota.
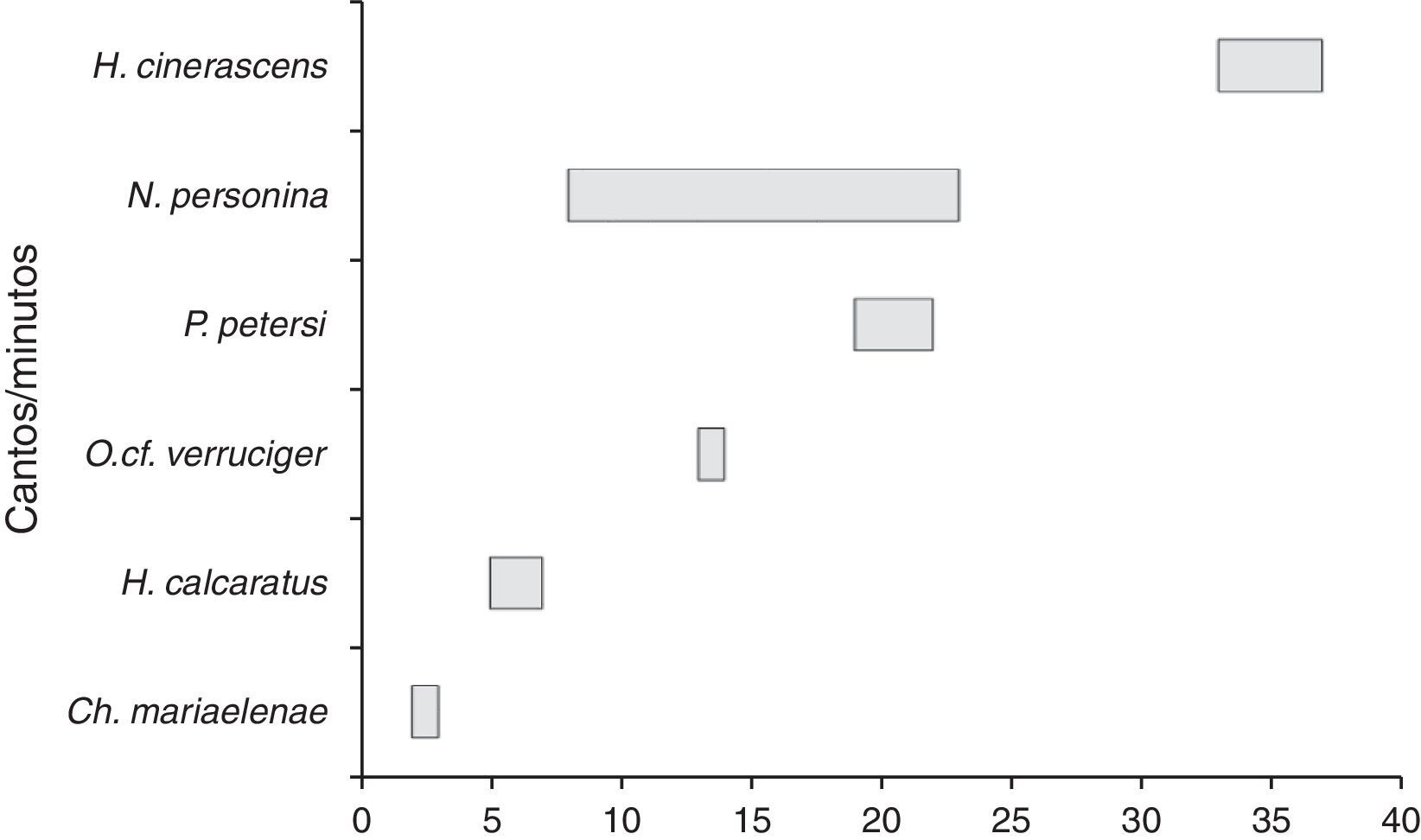
Las especies de mayor actividad vocal fueron H. cinerascens con un rango de 33-37 cantos/minuto, seguida por N. personina de 8-23 y P. petersi de 19-22 cantos/minuto. El resto de especies no sobrepasaron los 15 cantos, por tal motivo son las de menor actividad vocal; hay que notar que la mayor actividad no está ligada a la dominancia vocal, si bien la tasa de emisión es mayor en H. cinerascens, N. personina emite cantos en el día y en la noche y abarca una gran cantidad de microhábitats. En el caso de las especies con menor tasa de emisión tampoco implica que sean inactivas, ya que se registraron casos especiales como el de Ch. mariaelenae, que presentó una gran dominancia vocal formando coros de grandes agrupaciones en torno a la laguna (fig. 7).
DiscusiónCaracterización de los cantos
Las vocalizaciones de los anuros de la laguna Cormorán presentaron un patrón uniforme en sus variables espectrales y temporales; según la clasificación de Bogert (1960) corresponden a cantos de anuncio o advertencia, ya que son los cantos más comunes que emiten las especies, utilizando los mismos para advertir o anunciar receptividad sexual, posición y tamaño (Gerhardt y Huber, 2002).
ComparacionesTrueb y Duellman (1970) describieron los cantos de O. verruciger indicando que sus cantos generan notas pulsadas guturales con una frecuencia de alrededor de 1.4kHz; estos valores difieren del canto de O. cf. verruciger, donde la frecuencia dominante es menor (=0.71kHz). En cuanto a los lugares desde donde vocalizaban, señalan que son emitidos desde arbustos y rocas alrededor de charcas, observación similar en la especie de Sardinayacu, lo cual evidencia la dependencia reproductiva de estas especies a microhábitats acuáticos. La sistemática en torno al grupo Osteocephalus buckleyi, donde se encuentra O. verruciger, está en proceso de resolución, encontrando dificultades en las delimitaciones de las especies; Ron et al. (2012) mencionan que la genética es una gran herramienta, pero no suficiente para resolver estas dificultades, teniendo la necesidad de integrar datos morfométricos y descripciones de cantos; en este aspecto es evidente la diferencia de frecuencias entre O. verruciger descrita por Trueb y Duellman (1970) y la presentada en este estudio, existiendo la posibilidad que las poblaciones de Sardinayacu sean una especie no descrita.
Duellman (1978) describe de manera cualitativa y cuantitativa los cantos de H. cinerascens, anteriormente conocida como Hyla granosa, describiéndolos como una serie de 2-4 notas, cuyo sonido es parecido a un «boop–boop–boop» con una frecuencia dominante promedio de 1.38kHz, siendo un valor muy parecido al calculado en este estudio (=1.39kHz).
Caminer y Ron (2014) describen el canto de H. almendarizae, señalando que presentan una serie de 3-4 notas y enfatizando que las mismas suenan como un «quack»; así se tuvo un valor cualitativo y cuantitativo muy parecido al de este estudio. En cuanto a las variables espectrales indican que la frecuencia dominante promedio es de 1.95kHz, siendo ligeramente mayor a la descripción de este estudio (=1.49kHz).
Twomey, Delia y Castroviejo-Fisher (2014) describen el canto de Chimerella corleone, señalando que tiene una duración de 390ms compuesta de 2-3 notas, con periodos de tiempo muy largos entre cantos; Ch. corleone se presenta como una nueva especie de un género monotípico, donde solo se ubicaba a Ch. mariaelenae. El canto de Ch. corleone tiene las frecuencias más altas registradas para la familia Centrolenidae (7.42-7.62kHz), de esta manera se reafirma con las frecuencias de Ch. mariaelenae presentadas en este estudio (6.15-6.63kHz), una posible sinapomorfia sugerida en Twomey et al. (2014) para el género Chimerella a través de este rasgo bioacústico.
Diferencias intra- e interespecíficasEl análisis cualitativo y cuantitativo de las variables espectrales y temporales de los cantos nos indica que presentan frecuencias, duraciones de notas y pulsos diferentes entre las especies (Wells, 2007); estas diferencias interespecíficas permiten que las especies permanezcan aisladas reproductivamente y no existan confusiones por parte de la hembra en la selección del macho para el amplexo (Gerhardt y Huber, 2002). Esta característica evolutiva es notoria al registrar en la laguna Cormorán ensambles vocales, si bien poco estructurados, pero de patrones específicos como en Ch. mariaelenae y N. personina, cuyos cantos guardan una sincronización peculiar ante los de H. cinerascens, evitando de esta manera ser solapados en sus cantos. Littlejohn (1977) y Gerhardt y Huber (2002) mencionan que esta característica etológica se da ante especies más ruidosas, con la finalidad de evitar que los mensajes que contienen los cantos sean confusos y mal interpretados, ya que estas interferencias pueden llegar a disminuir las oportunidades reproductivas.
A nivel intraespecífico no se evidenciaron variaciones considerables en las vocalizaciones; si bien las características de los cantos presentan patrones distintivos para cada especie, varios autores, e.g., Gerhardt (1978); Lüddecke, Amézquita, Bernal, Guzmán y Agudelo (1999) y Valetti, Salas y Martino (2013) mencionan que pueden variar intraespecíficamente de acuerdo con las condiciones ambientales y contexto social.
Las especies aquí analizadas mayoritariamente cantaron en coros, en especial Ch. mariaelenae, donde la distribución espacial abarcó en su totalidad el margen litoral de la laguna, teniendo coros muy ordenados que se activaban bajo el detonante de un posible líder vocal; esta característica se ha registrado en la familia Bufonidae, donde Benson (1966) narra de manera cualitativa el comportamiento de las ranas toro, señalando que los mismos se activan a partir de un líder vocal; este comportamiento ha sido reportado en otras especies de anuros, sin esclarecer si el detonante se da en un mismo macho, o se da con un canto en un macho al azar (Wells, 2007).
Nuestros agradecimientos a los funcionarios del Parque Nacional Sangay zona baja, en especial a Víctor León por las facilidades de transporte y demás logísticas brindadas durante el trabajo de campo. A Ana Almendáriz (MEPN), quien apoyó institucionalmente el desarrollo de esta investigación y nos permitió el acceso a la colección bajo su cargo, brindándonos las facilidades durante el trabajo de laboratorio. A Mario Yánez-Muñoz por sus valiosos aportes y sugerencias en el manuscrito. A Gerardo Cevallos quien elaboró el mapa de ubicación del complejo lacustre de Sardinayacu. A Pablo Ávila, Toyo Álvarez, Cristian Torres y Marcelo Sharup (Churuwia) por su colaboración en el trabajo de campo. A las valiosas y acertadas correcciones y sugerencias de Camila Ferrara y 2 revisores anónimos. Al Ministerio del Ambiente de Morona Santiago quien otorgó el permiso de investigación N.o 04-2010-I-B-DPMS/MAE.
Especímenes grabados: Ecuador. Provincia de Morona Santiago. Complejo lacustre de Sardinayacu, Parque Nacional Sangay. Chimerella mariaelenae: MEPN 14514–15, 14519; Noblella personina: MEPN 14324, 14326–27; Pristimantis petersi: MEPN 14413, 14415, 14417; Hypsiboas almendarizae: MEPN 14541, 14699; Hypsiboas cinerascens: MEPN 14463–65; Osteocephalus cf. verruciger: MEPN 14474, 14479.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.