




La diversidad biológica no está uniformemente distribuida en la tierra, existiendo sitios más relevantes en virtud de sus propiedades intrínsecas o de sus contextos evolutivos y/o biogeográficos. Los Andes tropicales son reconocidos a nivel global por su alta diversidad y endemismos, y al mismo tiempo por los altos niveles de presiones antrópicas. En el presente estudio se evaluó la diversidad de mariposas Pronophilina, dominantes en riqueza de especies y abundancia en las partes altas de los Andes, en el área de influencia del páramo de Belmira, Antioquia, Colombia, al norte de la Cordillera Central. Se evaluó el aporte de esta área a la diversidad de mariposas altoandinas comparada con la reportada para el norte de los Andes tropicales. Se encontró un total de 50 especies de la subtribu Pronophilina, lo que representa cerca de la cuarta parte de la riqueza de especies del grupo conocidas para Colombia y del 10% de la riqueza global de la misma, incluyendo especies únicas y de distribución restringida. Los presentes resultados aportan evidencias sobre la importancia de las formaciones montañosas aisladas en el extremo norte de los Andes, reforzando la idea de que el páramo de Belmira es un componente clave para la conservación de la diversidad de las mariposas altoandinas a nivel regional, que amerita mayores medidas para su conservación y estatus de protección.
Biodiversity is not uniformly distributed on earth, with sites more relevant than others, due to their intrinsic properties or their evolutionary and/or biogeographical context. The tropical Andes are globally acknowledged for their importance for preserving high biodiversity and endemism, and at the same time by the high level of anthropogenic pressure. In this study, we assessed the subtribe Pronophilina butterfly diversity, the dominant group in species richness and abundance in the higher Andes, in the influence area of the Páramo de Belmira, Antioquia, Colombia, in the northern part of the Central Cordillera. The contribution of this area to the overall regional Andean butterfly diversity reported for the northern tropical Andes was assessed. In Belmira, 50 Pronophilina species were found, representing approximately one fourth of the known Colombian species of this subtribe and 10% of the overall richness, including unique and geographically restricted species. These results provide evidence on the importance of isolated mountain formations in the northern end of the Andes and confirm the assumption that the Páramo de Belmira is a key conservation component for the high Andean butterfly diversity at the regional level, deserving stronger conservation actions and protection status.
Los Andes tropicales han sido reconocidos como una de las regiones más relevantes a nivel global en términos de biodiversidad, siendo propuestos como uno de los puntos claves globales («hot-spot»), debido a su alta variabilidad y tasas de endemismo en plantas y vertebrados (Myers, Mittermeier, Mittermeier, da Fonseca y Kent, 2000). Esto mismo aplica para insectos, dentro de los cuales el orden Lepidoptera, con grupos como Geometridae (Brehm, Pitkin, Hilt y Fiedler, 2005), Ithomiini (Chazot et al., 2016) y Arctiidae (Hilt, 2005), presenta una alta diversidad en esta región. Asimismo, para la superfamilia Papilionoidea se ha reportado una gran diversidad y un alto número de endemismos, particularmente en las partes altas de las montañas y en los macizos más aislados (Adams, 1984, 1985; Adams y Bernard, 1977, 1979; Chazot et al., 2016; Espeland et al., 2015; Robbins y Busby, 2008; Viloria, 2003), presentando un amplio número de especies con rangos altitudinales restringidos (Casner y Pyrcz, 2010; Pyrcz, Wojtusiak y Garlacz, 2009). Esto ha llevado al desarrollo de ensambles de especies únicos entre las diferentes cadenas montañosas (Marín, Giraldo, Marín, Álvarez y Pyrcz, 2015; Pyrcz, 2004; Pyrcz y Garlacz, 2012; Pyrcz, Viloria, Lamas y Boyer, 2011), siendo Satyrinae la subfamilia más destacada (Adams, 1973), con un gran número de especies endémicas, particularmente en la subtribu Pronophilina, y en algunos grupos de Lycaenidae, Pieridae y Biblidinae, que también presentan una amplia diversificación en las partes altas de los Andes (Adams, 1985, 1986; Bollino y Costa, 2007; Prieto, 2011; Pyrcz, 2004; Pyrcz y Rodríguez, 2007; Robbins y Busby, 2008).
Las formaciones aisladas al norte de los Andes, como la Sierra Nevada de Santa Marta y la serranía del Perijá (Adams, 1985), se comportan como islas intracontinentales para muchos grupos de lepidópteros que se adaptaron a las partes más elevadas, lo que ocurre también en las cadenas montañosas. Por ejemplo, en la Cordillera Occidental, el nivel de endemismo de las mariposas de la subtribu Pronophilina es de un 25%, habitando principalmente en las zonas de subpáramo y páramo (Pyrcz y Rodríguez, 2007). En este mismo sentido, para el norte de la Cordillera Central es de esperarse que los bosques altoandinos y páramos de las regiones montañosas más aisladas alberguen una considerable riqueza y endemismo de especies, conformando probables centros locales de endemismo para mariposas (Pyrcz y Rodríguez, 2006; Pyrcz et al., 2016). En el presente estudio se analizó la importancia del área de influencia del páramo de Belmira, también conocido como páramo de Santa Inés, última formación montañosa con más de 3,000m de altura en el extremo norte de la Cordillera Central, estableciendo su aporte a la diversidad regional de Pronophilina del norte de los Andes.
Materiales y métodosLa investigación se realizó en el área de influencia del páramo de Belmira, en adelante denominada como Belmira, ubicada en el extremo norte de la Cordillera Central de los Andes en Colombia, (6°35′-6°51′N, 75°47′-75°38′O), a 32km al noroccidente de la ciudad de Medellín, departamento de Antioquia, y distante de otras formaciones montañosas similares en la Cordillera Central, como el páramo de Sonsón a más de 100km, y de la Cordillera Occidental, como del Nudo de Paramillo y el páramo de Frontino-Urrao, a más de 50km. El área está incluida en el Distrito de Manejo Integrado del Sistema de Páramos y Bosques del Noroccidente Medio Antioqueño (DMI-SPBNMA), figura de conservación y de manejo mixto, la cual comprende cerca de 42,600ha, donde el 14.54% de la cobertura original se ha transformado (IAvH, 2013). Incluye zonas de bosque muy húmedo montano bajo (bmh-Mb) y bosque pluvial montano (bp-Mp) (Espinal, 1985), con áreas denominadas como subpáramo (>3,000m), presentando precipitaciones entre los 1,900 y los 2,200mm anuales en un régimen monomodal (CORANTIOQUIA, 1999; Morales et al., 2007).
El inventario se efectuó entre junio del 2011 y abril del 2014, con 133 días de campo desarrollados de forma alternante entre 12 localidades situadas desde 2,650 a 3,300m, con coberturas vegetales de pastizal, de bosque de niebla, de páramo y de mosaico, que incluyen pastos, bosques y diferentes estadíos sucesionales de regeneración natural. De forma similar a Marín, Álvarez, Giraldo, Pyrcz y Uribe (2014), trabajando entre las 8a.m. y 4p.m., con 2-3 equipos de 2 personas por jornada por localidad, se efectuaron capturas con red entomológica y trampas Van Someren-Rydon cebadas con pescado en descomposición, banano fermentado y coprocebo. Se ubicaron en promedio 8 trampas por localidad, con un muestreo mínimo por cobertura de 28 días, alcanzando un esfuerzo total de muestreo cercano a 4,500h.
El procesamiento del material se realizó en el Laboratorio de Biología y Sistemática de Insectos, Universidad Nacional de Colombia sede Medellín, con la colaboración del Zoological Museum of the Jagiellonian University en Cracovia, Polonia. Posteriormente, los ejemplares fueron depositados en la colección del Museo Entomológico Francisco Luis Gallego (MEFLG) de la Universidad Nacional de Colombia sede Medellín, con copias en la colección del Instituto Alexander von Humboldt (Villa de Leyva). Los ejemplares en su mayoría fueron identificados a especie, empleando fuentes de información como De Vries (1987), Lewis (1975), Pulido y Parrales (2009), Valencia, Gil y Constantino (2005), Warren, Davis, Stangeland, Pelham y Grishin (2014), y comparados con los ejemplares depositados en el MEFLG.
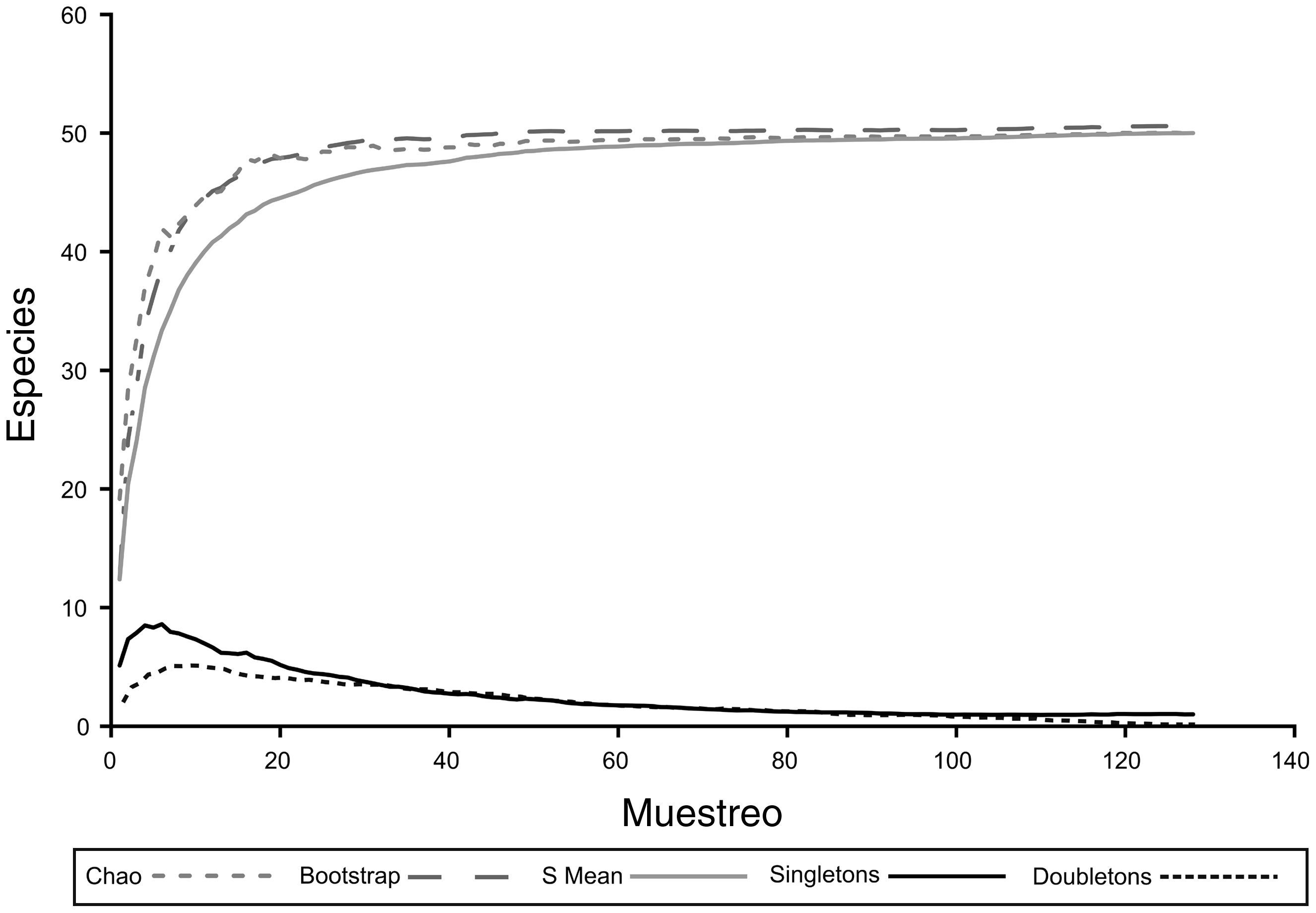
Para estimar la representatividad y eficiencia del muestreo, se construyeron curvas de acumulación de especies y estimadores de diversidad (Colwell y Coddington, 1994) con el programa EstimateS 8.2 (Colwell, 2009), lo que se complementó con el cálculo de la completitud de muestreo empleando la plataforma en línea iNEXT (http://chao.stat.nthu.edu.tw/blog/software-download/inext-online/) (Hsieh, Ma y Chao, 2013).
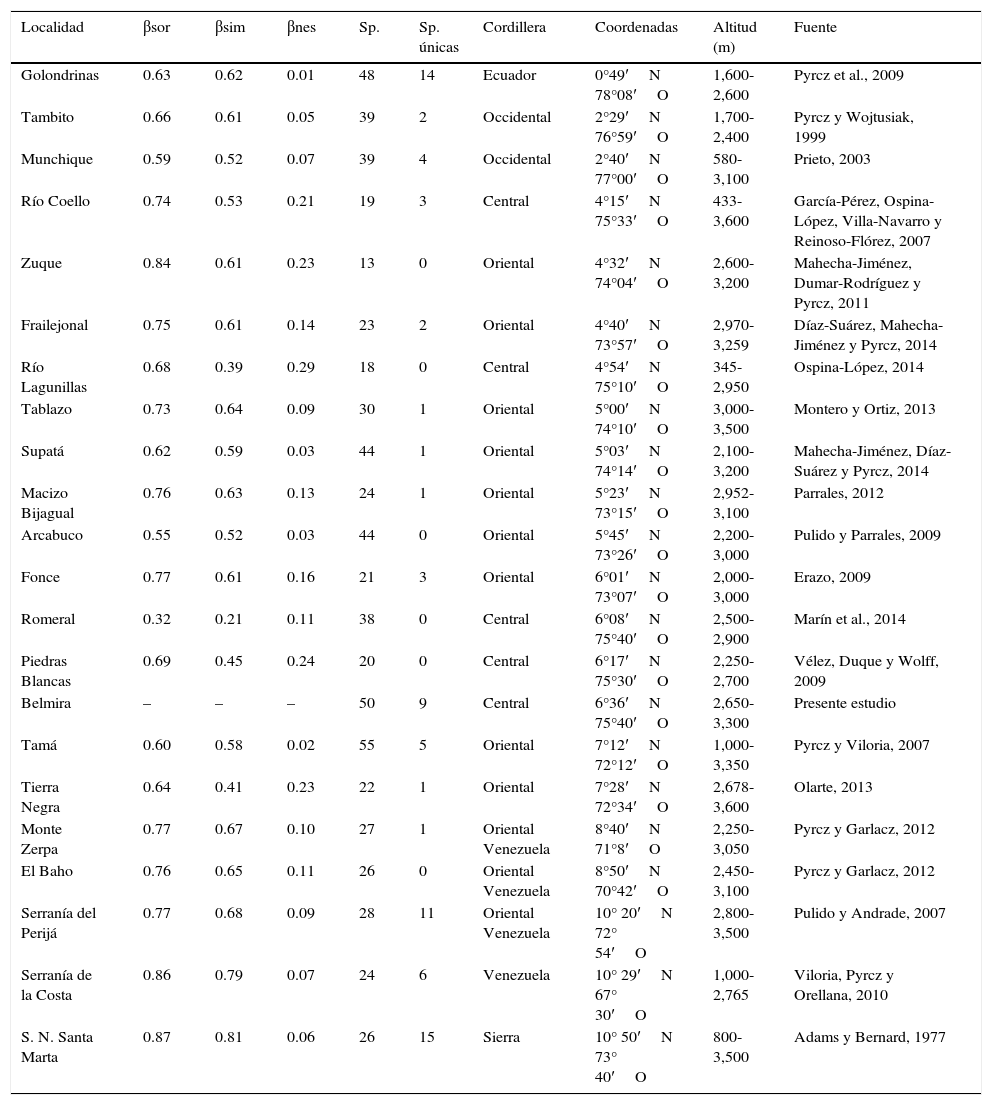
Para comparar el ensamble de especies de Belmira con otras localidades ubicadas al norte de los Andes, se consideró a la subtribu Pronophilina a partir de los inventarios presentados en referencias bibliográficas, en zonas que incluyeran muestreos sobre los 2,000m (fig. 1). La comparación, dadas las diferencias metodológicas y la disponibilidad de datos de las fuentes de información secundarias, se basó en la riqueza (presencia-ausencia) de especies, contabilizando las especies por localidad y registrando sus aportes por especies únicas.

Se exploraron las diferencias entre los ensambles de mariposas del área de Belmira con los demás sitios, efectuando una comparación de la diferenciación pareada entre localidades a través de la partición aditiva en 3 diferentes componentes de la diversidad beta (β), βsor=βsim+βnes (Baselga, 2010). βsor es la diferencia general en la composición entre sitios y es un resultado de la suma del reemplazo (βsim) y la diferencia en riqueza de especies (βnes). Igualmente se calculó el índice de Jaccard, generando un dendrograma de similitud y un escalamiento multidimensional no métrico (nMDS), explorando el agrupamiento por similitud entre las localidades. Estos análisis fueron realizados con el software R empleando la librería «vegan» (Oksanen et al., 2016).
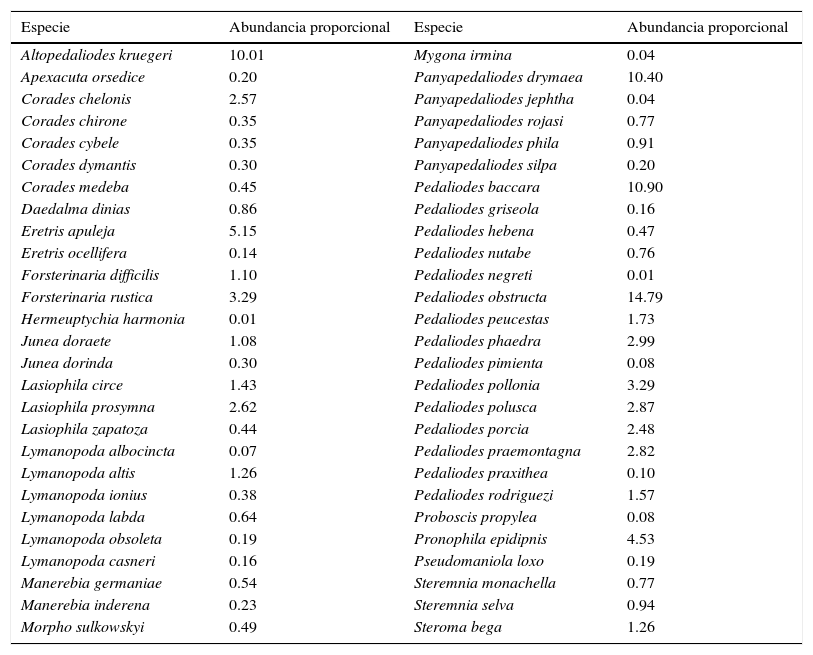
ResultadosUn total de 50 especies de Pronophilina fueron registradas en Belmira (tabla 1). Cinco especies componen más del 50% de los individuos registrados: Pedaliodes obstructa (14.79%), Pedaliodes baccara (10.9%), Panyapedaliodes drymaea (10.40%), Altopedaliodes kruegeri (10.01%) y Eretris apuleja (5.16%). Las curvas de acumulación de especies y estimativos de diversidad presentaron una tendencia asintótica con un decrecimiento gradual de «singletons» y «doubletons» (fig. 2), mientras que la estimación de la cobertura de muestreo fue del 0.99.
Diversidad y abundancias relativas de mariposas Satyrinae en el área de Belmira.
| Especie | Abundancia proporcional | Especie | Abundancia proporcional |
|---|---|---|---|
| Altopedaliodes kruegeri | 10.01 | Mygona irmina | 0.04 |
| Apexacuta orsedice | 0.20 | Panyapedaliodes drymaea | 10.40 |
| Corades chelonis | 2.57 | Panyapedaliodes jephtha | 0.04 |
| Corades chirone | 0.35 | Panyapedaliodes rojasi | 0.77 |
| Corades cybele | 0.35 | Panyapedaliodes phila | 0.91 |
| Corades dymantis | 0.30 | Panyapedaliodes silpa | 0.20 |
| Corades medeba | 0.45 | Pedaliodes baccara | 10.90 |
| Daedalma dinias | 0.86 | Pedaliodes griseola | 0.16 |
| Eretris apuleja | 5.15 | Pedaliodes hebena | 0.47 |
| Eretris ocellifera | 0.14 | Pedaliodes nutabe | 0.76 |
| Forsterinaria difficilis | 1.10 | Pedaliodes negreti | 0.01 |
| Forsterinaria rustica | 3.29 | Pedaliodes obstructa | 14.79 |
| Hermeuptychia harmonia | 0.01 | Pedaliodes peucestas | 1.73 |
| Junea doraete | 1.08 | Pedaliodes phaedra | 2.99 |
| Junea dorinda | 0.30 | Pedaliodes pimienta | 0.08 |
| Lasiophila circe | 1.43 | Pedaliodes pollonia | 3.29 |
| Lasiophila prosymna | 2.62 | Pedaliodes polusca | 2.87 |
| Lasiophila zapatoza | 0.44 | Pedaliodes porcia | 2.48 |
| Lymanopoda albocincta | 0.07 | Pedaliodes praemontagna | 2.82 |
| Lymanopoda altis | 1.26 | Pedaliodes praxithea | 0.10 |
| Lymanopoda ionius | 0.38 | Pedaliodes rodriguezi | 1.57 |
| Lymanopoda labda | 0.64 | Proboscis propylea | 0.08 |
| Lymanopoda obsoleta | 0.19 | Pronophila epidipnis | 4.53 |
| Lymanopoda casneri | 0.16 | Pseudomaniola loxo | 0.19 |
| Manerebia germaniae | 0.54 | Steremnia monachella | 0.77 |
| Manerebia inderena | 0.23 | Steremnia selva | 0.94 |
| Morpho sulkowskyi | 0.49 | Steroma bega | 1.26 |

El área de Belmira presentó el segundo mayor número de especies al norte de los Andes después del Tamá, ubicada en la Cordillera Oriental, con 55 especies. Igualmente fue una de las localidades con mayor número de especies únicas y reducida presencia regional (tabla 2). Del análisis de diversidad beta se puede observar una amplia disimilitud de Belmira con otras localidades, con un valor de βsor>0.50, a excepción de Romeral que fue de 0.32. Los valores de βsim y βnes indican que hay un alto reemplazo de especies entre Belmira y las otras localidades (βsim>0.41), con una baja diferencia en la proporción de la riqueza entre localidades (βnes<0.29) con respecto al área de estudio.
Comparación de la composición de especies de Pronophilina del área de estudio (Belmira) con diferentes localidades al norte de los Andes, con disimilitud de Sørensen (βsor), disimilitud de Simpson (βsim) y disimilitud anidada (βnes).
| Localidad | βsor | βsim | βnes | Sp. | Sp. únicas | Cordillera | Coordenadas | Altitud (m) | Fuente |
|---|---|---|---|---|---|---|---|---|---|
| Golondrinas | 0.63 | 0.62 | 0.01 | 48 | 14 | Ecuador | 0°49′N 78°08′O | 1,600-2,600 | Pyrcz et al., 2009 |
| Tambito | 0.66 | 0.61 | 0.05 | 39 | 2 | Occidental | 2°29′N 76°59′O | 1,700-2,400 | Pyrcz y Wojtusiak, 1999 |
| Munchique | 0.59 | 0.52 | 0.07 | 39 | 4 | Occidental | 2°40′N 77°00′O | 580-3,100 | Prieto, 2003 |
| Río Coello | 0.74 | 0.53 | 0.21 | 19 | 3 | Central | 4°15′N 75°33′O | 433- 3,600 | García-Pérez, Ospina-López, Villa-Navarro y Reinoso-Flórez, 2007 |
| Zuque | 0.84 | 0.61 | 0.23 | 13 | 0 | Oriental | 4°32′N 74°04′O | 2,600-3,200 | Mahecha-Jiménez, Dumar-Rodríguez y Pyrcz, 2011 |
| Frailejonal | 0.75 | 0.61 | 0.14 | 23 | 2 | Oriental | 4°40′N 73°57′O | 2,970-3,259 | Díaz-Suárez, Mahecha-Jiménez y Pyrcz, 2014 |
| Río Lagunillas | 0.68 | 0.39 | 0.29 | 18 | 0 | Central | 4°54′N 75°10′O | 345-2,950 | Ospina-López, 2014 |
| Tablazo | 0.73 | 0.64 | 0.09 | 30 | 1 | Oriental | 5°00′N 74°10′O | 3,000-3,500 | Montero y Ortiz, 2013 |
| Supatá | 0.62 | 0.59 | 0.03 | 44 | 1 | Oriental | 5°03′N 74°14′O | 2,100-3,200 | Mahecha-Jiménez, Díaz-Suárez y Pyrcz, 2014 |
| Macizo Bijagual | 0.76 | 0.63 | 0.13 | 24 | 1 | Oriental | 5°23′N 73°15′O | 2,952-3,100 | Parrales, 2012 |
| Arcabuco | 0.55 | 0.52 | 0.03 | 44 | 0 | Oriental | 5°45′N 73°26′O | 2,200-3,000 | Pulido y Parrales, 2009 |
| Fonce | 0.77 | 0.61 | 0.16 | 21 | 3 | Oriental | 6°01′N 73°07′O | 2,000-3,000 | Erazo, 2009 |
| Romeral | 0.32 | 0.21 | 0.11 | 38 | 0 | Central | 6°08′N 75°40′O | 2,500-2,900 | Marín et al., 2014 |
| Piedras Blancas | 0.69 | 0.45 | 0.24 | 20 | 0 | Central | 6°17′N 75°30′O | 2,250-2,700 | Vélez, Duque y Wolff, 2009 |
| Belmira | – | – | – | 50 | 9 | Central | 6°36′N 75°40′O | 2,650-3,300 | Presente estudio |
| Tamá | 0.60 | 0.58 | 0.02 | 55 | 5 | Oriental | 7°12′N 72°12′O | 1,000-3,350 | Pyrcz y Viloria, 2007 |
| Tierra Negra | 0.64 | 0.41 | 0.23 | 22 | 1 | Oriental | 7°28′N 72°34′O | 2,678-3,600 | Olarte, 2013 |
| Monte Zerpa | 0.77 | 0.67 | 0.10 | 27 | 1 | Oriental Venezuela | 8°40′N 71°8′O | 2,250-3,050 | Pyrcz y Garlacz, 2012 |
| El Baho | 0.76 | 0.65 | 0.11 | 26 | 0 | Oriental Venezuela | 8°50′N 70°42′O | 2,450-3,100 | Pyrcz y Garlacz, 2012 |
| Serranía del Perijá | 0.77 | 0.68 | 0.09 | 28 | 11 | Oriental Venezuela | 10° 20′N 72° 54′O | 2,800-3,500 | Pulido y Andrade, 2007 |
| Serranía de la Costa | 0.86 | 0.79 | 0.07 | 24 | 6 | Venezuela | 10° 29′N 67° 30′O | 1,000-2,765 | Viloria, Pyrcz y Orellana, 2010 |
| S. N. Santa Marta | 0.87 | 0.81 | 0.06 | 26 | 15 | Sierra | 10° 50′N 73° 40′O | 800-3,500 | Adams y Bernard, 1977 |
Datos ordenados según la latitud. Las coordenadas y alturas deben considerarse como referencias generales de área, se tomaron a partir de las fuentes, estimándose a partir de conocimiento propio o de información secundaria no citada sobre las zonas.
El análisis de agrupamiento presentó a la Sierra Nevada de Santa Marta como una unidad aislada. Asimismo, presentó una muy baja similitud de las localidades, entre el 20 y el 40% (fig. 3a), con excepción de El Baho y Monte Zerpa, en Venezuela, que comparten todas sus especies menos una. En el nMDS (fig. 3b) se evidenció una cercanía entre localidades de la Cordillera Oriental y de las zonas aisladas al norte de Colombia y Venezuela, aunque con niveles de similitud bajos entre todas las localidades.
 Análisis de agrupamientos (Índice de Sørensen) de especies de Pronophilina al norte de los Andes. b) Escalamiento multidimensional no métrico (nMDS). Arc: Arcabuco; Bah: El Baho; Bel: Belmira; Bij: Macizo del Bijagual; Coe: Río Coello; Cos: serranía de la Costa; Fon: Fonce; Fra: Frailejonal; Gol: Golondrinas; Lag: Río Lagunillas; Mon: Monte Zerpa; Mun: Munchique; Per: serranía del Perijá; Pie: Piedras Blancas; Rom: Romeral; Sie: Sierra Nevada de Santa Marta; Sup: Supatá; Tab: Tablazo; Tam: Tamá; Tie: Tierra Negra; Tmb: Tambito; Zuq: Zuque.")
a) Análisis de agrupamientos (Índice de Sørensen) de especies de Pronophilina al norte de los Andes. b) Escalamiento multidimensional no métrico (nMDS).
Arc: Arcabuco; Bah: El Baho; Bel: Belmira; Bij: Macizo del Bijagual; Coe: Río Coello; Cos: serranía de la Costa; Fon: Fonce; Fra: Frailejonal; Gol: Golondrinas; Lag: Río Lagunillas; Mon: Monte Zerpa; Mun: Munchique; Per: serranía del Perijá; Pie: Piedras Blancas; Rom: Romeral; Sie: Sierra Nevada de Santa Marta; Sup: Supatá; Tab: Tablazo; Tam: Tamá; Tie: Tierra Negra; Tmb: Tambito; Zuq: Zuque.
En el presente estudio, se logró determinar la relevancia del área de influencia del páramo de Belmira para la conservación de mariposas altoandinas y su aporte a la diversidad regional del norte de los Andes, a nivel de composición de especies y en la diferenciación con otras áreas del norte de los Andes.
Se ha señalado que las zonas montañosas de los Andes presentan una reducida diversidad de especies en comparación con las zonas bajas adyacentes (Shapiro, 1992). Aun así, el número de especies registradas en Belmira aparece como representativo de la zona, según lo indican las curvas de acumulación (fig. 2) y la estimación de la cobertura de muestreo. Se destaca la importancia del muestreo a largo plazo en la zona, habiéndose encontrado especies poco representadas geográficamente, como Pedaliodes hebena, Pedaliodes pimienta o Pedaliodes rodriguezi, así como especies nuevas para la ciencia denominadas Lymanopoda casneri, Panyapedaliodes rojasi y Pedaliodes nutabe (Pyrcz et al., 2016).
Pyrcz, Willmott, Garlacz, Boyer y Gareca (2014) señalan que la edad geológica de los Andes y la estacionalidad climática explican la mayor parte de la variación en la riqueza de especies de mariposas. La heterogeneidad espacial de Belmira, asociada a su ubicación geográfica en la zona más al norte de la Cordillera Central, así como la cercanía al cañón del río Cauca, límite con la Cordillera Occidental y al Altiplano norte Antioqueño en la Cordillera Central, podrían relacionarse con la riqueza encontrada en la zona de investigación. Se ha señalado que la existencia de áreas por encima de 2,000m así como la profundidad de los valles adyacentes a las zonas de montaña pueden tener una alta influencia en la diversidad de especies y los endemismos (Viloria, 2002), elementos presentes en el área de estudio.
Los resultados demuestran que Belmira es un importante componente de la diversidad de especies a nivel regional al norte de los Andes. Por ejemplo, al comparar Belmira con Romeral y Piedras Blancas, las localidades más cercanas geográficamente, se obtuvo un valor de disimilitud βsor de 0.32 con Romeral y de 0.69 con Piedras Blancas (tabla 2). Adicionalmente, considerando la diferencia en la proporción de la riqueza entre Piedras Blancas y Belmira de βnes=0.24, se sugiere que en Piedras Blancas se han perdido especies que están presentes en Belmira, resaltando la relevancia del área de estudio para la conservación de las mariposas de montaña al norte de los Andes.
En especies únicas, Belmira solo fue superado en Colombia por lo encontrado en la Sierra Nevada y por la serranía del Perijá, ambos macizos montañosos aislados de los Andes (fig. 1). Los extremos de las cadenas montañosas o cordilleras suelen presentar composiciones bióticas particulares en sus comunidades (Carlos Prieto, comunicación personal), tal como se ha encontrado en Belmira. El análisis de diversidades beta (fig. 3) enfatiza el aporte de diferentes localidades a la riqueza regional de especies, adicional a lo identificado por el número de especies y por la presencia de especies únicas por localidad.
A escala regional, el norte de los Andes no presentó un patrón latitudinal de riqueza de especies de Pronophilina, a diferencia de lo encontrado a escala continental (Pyrcz et al., 2014), lo que podría asociarse a patrones biogeográficos específicos de esta región y a diferencias asociadas a las fuentes de información. En varias localidades se encontraron especies únicas, lo cual es coherente con la reducida similitud evidenciada por el análisis de agrupamiento y concordante con la idea de la singularidad de las comunidades de Pronophilina asociadas a áreas montañosas aisladas (Adams, 1985). Asimismo, se destaca la similitud evidenciada por el análisis de agrupamiento y por el nMDS entre Romeral y Belmira con localidades de la Cordillera Occidental distantes geográficamente cerca de 500km, además del profundo cañón del Cauca. Esto podría asociarse al impacto que tiene el Valle del Magdalena, al ser más amplio y presentar una mayor dificultad para el movimiento de individuos entre la Cordillera Central y la Cordillera Oriental. Los resultados obtenidos concuerdan con Kattan, Franco, Rojas y Morales (2004), en términos de la similitud del occidente de la Cordillera Central con las vertientes del Pacífico de la Cordillera Occidental. En general, la similitud entre comunidades andinas decrece con la distancia y es variable según las capacidades de dispersión y la distribución de las especies de flora y fauna (Anthelme et al., 2014).
El presente estudio resalta la importancia del área de Belmira en la biodiversidad regional de Pronophilina al norte de los Andes tropicales (50 especies), con cerca del 25% de las especies reconocidas para Colombia (208 especies) y del 10% de las reportadas a nivel global (520 especies) (Pyrcz, Prieto, Viloria y Andrade, 2013). La presencia de especies únicas y de limitada aparición en otras localidades, así como de varias especies nuevas y endémicas (Pyrcz y Rodríguez, 2006; Pyrcz et al., 2016), hacen de Belmira una zona clave para la conservación de la lepidopterofauna en los Andes.
Muchas de las especies de mariposas altoandinas están distribuidas en estrechas bandas altitudinales (Casner y Pyrcz, 2010; Pyrcz y Wojtusiak, 2002; Pyrcz et al., 2009), presentándose diferencias en los ensambles de especies en diferentes rangos altitudinales. El presente estudio refleja esta variación, mostrando diferencias del ensamble de especies entre zonas inferiores a 3,000-3,100m (bosque de niebla) con las alturas superiores (páramo), sugiriendo que para el desarrollo de medidas de conservación es necesario incluir el máximo rango altitudinal posible, lo que evidencia la necesidad de fortalecer el estatus de conservación del área del páramo de Belmira incluyendo zonas por debajo de los 2,800m.
A las comunidades locales, al personal del DMI-SPBNMA y entidades del municipio de Belmira, así como a la autoridad ambiental (CORANTIOQUIA) por su colaboración administrativa y en campo. CFAH agradece a la Corporación Universitaria Lasallista, a la Universidad Nacional de Colombia sede Medellín y a Colciencias (Convocatoria 528/2011), por el apoyo a la investigación y a la formación doctoral. MAM agradece a la FAPESP (2014/16481-0) por la beca para la realización de estudios de posgrado. Gracias al GSM, especialmente a Carlos E. Giraldo, Andrés Rojas, Francisco Restrepo Carrasquilla y Leydi Areiza R. por su apoyo en el campo, laboratorio y análisis. Se agradece a Oscar Mahecha, Carlos Prieto y Dumar Parrales por su aporte de información, así como a los colaboradores y revisores anónimos por sus esfuerzos, comentarios y sugerencias.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.