


Con base en un análisis exhaustivo de la bibliografía de Lepidoptera relacionada con México y en la revisión de la megabase mariposa (>500 000 registros y ejemplares), se presenta una síntesis sobre la riqueza mundial, sus estimados y sus proporciones con México, así como un examen detallado de la distribución de la riqueza y el endemismo de Papilionoidea s. lat, una de las superfamilias mejor conocidas para México. La distribución va precedida por un resumen de las contribuciones del equipo respecto a estudios estatales, fisiográficos y vegetacionales realizados durante las últimas 3 décadas. En cifras resumidas, se estima que México contiene 23 750 especies de Lepidoptera, con cerca de 14 500 descritas y documentadas. La cifra real y la estimada se acercan al 10% de representación en México. Lo mismo ocurre con el porcentaje de Papilionoidea (s. lat.) de México respecto al mundial. Los porcentajes de endemismo en especies de las familias de Papilionoidea para el país fueron: Hesperiidae 15.83%, Papilionidae 6.3%, Pieridae 12.98%, Lycaenidae 5.27%, Riodinidae 16.29% y Nymphalidae 11.86%. Si se consideran las subespecies, el endemismo en general aumenta a 22.2%, casi la cuarta parte de los taxones de este nivel. Se tabula el número de especies y las endémicas representadas en cada estado y provincia biogeográfica a partir de una lista con cerca de 450 subespecies endémicas. El estado de Oaxaca y la provincia biogeográfica Sierra Madre del Sur contienen la mayor singularidad proporcional por su endemismo, mientras que Chiapas y la provincia Costa del Golfo de México contienen la mayor riqueza.
Based on a thorough analysis of the literature on Lepidoptera associated with Mexico, and a review of the database mariposa (>500 000 records and specimens), a synthesis of the global Lepidoptera diversity is presented, including estimates and proportions of diversity and endemism in Mexico. In addition, a detailed examination of the species richness and endemism of one of the best-known families in Mexico, Papilionoidea s. lat., is provided, preceded by a summary of state-level, habitat-specific and transect-based research on Lepidoptera diversity conducted over the past 3 decades. In summary, Mexico hosts about 14 500 documented species of Lepidoptera, with about 23 750 species estimated. These figures suggest that 10% of the world’s Lepidoptera taxa are represented in Mexico. Among Papilionoidea, similarly, about 10% of the world’s taxa are reported from Mexico. Percentages of endemic species for the various families of Papilionoidea in Mexico are: Hesperiidae 15.83%, Papilionidae 6.3%, Pieridae 12.98%, Lycaenidae 5.27%, Riodinidae 11.86% and Nymphalidae 16.29%. When subspecies are considered, 22.2% of these taxa are endemic to Mexico, almost a quarter of all subspecies in the country. The number of endemic taxa represented in each state and biogeographic province is tabulated from a list of about 450 endemic subspecies. Oaxaca and the Sierra Madre del Sur represent the state and biogeographic province with the highest proportion of endemic taxa, while Chiapas and the Gulf Coast region represent the state and biogeographic province hosting the greatest overall diversity.
Los Arthropoda conforman cerca del 80% del total del reino Animalia; este último registra a la fecha poco más de 1 550 000 especies (Zhang, 2011). Los Lepidoptera, junto con Diptera, Hymenoptera y Coleoptera se reconocen como los 4 órdenes hiperdiversos de la clase Insecta (Martin-Piera et al., 2000), con un número de especies superior a los 650 000. De los Lepidoptera se han descrito y denominado unas 155 000 especies (Apéndice), es decir, constituyen el 10% del total de las especies animales; aunque las estimaciones oscilan entre 255 000 y cerca del medio millón de especies (Kristensen et al., 2007).
Avances importantes sobre la filogenia, la clasificación y análisis de la diversidad del reino Animalia pueden encontrarse en Zhang y Shear (2007) y Zhang (2011), quienes recapitulan escuetamente sobre los Lepidoptera. Sobre este grupo hiperespecioso, Heppner (1991, 1998), Scoble (1992), Kristensen (1998, 2004) y Kristensen et al. (2007) han realizado estudios sintéticos. Tales monografías son referencia obligada para todos los lepidopterólogos interesados en la diversidad y evolución de dicho orden.
PublicacionesDado que diversos subgrupos de Lepidoptera se consideran modelos de estudio biológico general en varias disciplinas, a menudo aparecen textos especializados sobre ello, que reúnen a los principales expertos mundiales. Por ejemplo, en genética, el libro de Goldsmith y Marec (2010); en bioconservación, la compilación de Dover et al. (2011); en embriología endocrinológica, el volumen colectivo de Goldsmith y Wilkins (2006); en ecología evolutiva de mariposas, el de Boggs et al. (2003) o el editado por Vane Wright y Ackery (1989): The biology of the butterflies.
No existe un volumen integral y contemporáneo sobre la historia de la lepidopterología; lo que se conoce está disperso en las introducciones o antecedentes de muchas y variadas monografías, revisiones y atlas regionales o mundiales. Tales contribuciones con frecuencia se reducen a algunos periodos de tiempo, a algunos subgrupos o taxones, o bien, a algunas subregiones o países (v. gr,Michán et al., 2004, 2005).
Los estudios generales más extensos para los Lepidoptera del mundo o de México y Centroamérica continúan siendo Die Gross-Schmetterlinge der Erde (The Macrolepidoptera of the world) editado por Seitz (1906-1954) y la Biologia Centrali Americana de Godman y Salvin (1878-1901), hoy en línea (http://www.sil.si.edu/digitalcollections/bca). Para el país se pueden agregar resúmenes de actualización, como el Catálogo sistemático y zoogeográfico de los lepidópteros mexicanos de Hoffmann (1940, 1941, 1942) y los distintos capítulos que aparecen en Biodiversidad, taxonomía y biogeografía de los artrópodos de México: hacia una síntesis de su conocimiento, obra de Llorente et al. (1996a, 2000, 2004b) y Llorente y Morrone, (2002), editada en 4 volúmenes.
En las últimas 4 décadas se han generado numerosas monografías, revisiones y atlas de gran relevancia para las grandes regiones biogeográficas donde México está comprendido. Por su calidad, participación de expertos e ilustraciones sobresale la importante serie The moths of America, North of Mexico, que desde 1970 se publicó fascicularmente durante más de 2 décadas, y el Atlas of Neotropical Lepidoptera, que John Heppner edita mediante la Association for Tropical Lepidoptera con sede en Gainesville, Florida.
A lo largo del siglo XX, numerosas revistas entomológicas y lepidopterológicas, además de publicaciones especializadas de los órganos de difusión de muchos museos y sociedades de entomología o lepidopterología (locales, nacionales, regionales o mundiales), han publicado miles o tal vez decenas de miles de trabajos sobre el orden Lepidoptera o sus subgrupos. Síntesis de esas publicaciones aparecen, como ejemplos importantes, en Heppner (1991), Kristensen (1998) y Lamas (2000, 2008, 2010).
Sólo para Hesperioidea y Papilionoidea, Lamas et al. (1995) y Lamas (2010) compilaron alrededor de 13 000 trabajos ropalocerológicos en el Neotrópico, básicamente desde Linnaeus (1758) a la fecha. Luis et al. (2000) registraron un número próximo a los 1 800 trabajos sobre el mismo grupo y en el mismo periodo para México. Debe considerarse que los Rhopalocera comprenden cerca del 12% de los Lepidoptera y tal vez sea uno de los grupos mejor conocidos, pero aún no bien conocidos. Entonces, un catálogo completo del número de publicaciones sobre Rhopalocera en el mundo posiblemente supere los 50 000 y para el total de los Lepidoptera quizá sean más de un cuarto de millón las citas bibliográficas.
En la última década han surgido varias fuentes electrónicas profesionales, cada vez mejores y más frecuentemente consultadas (v. gr,Lamas, 2000, 2010; Kristensen et al., 2007; Beccaloni et al. http://www.nhm.ac.uk/research-curation/projects/lepindex/index.html). Finalmente, es relevante citar la obra coordinada por Nye (1975-1991), que en 6 volúmenes compila los más de 15 000 nombres genéricos de las polillas del mundo, una obra fundamental para todo lepidopterólogo.
Instituciones y museosEl sistema de referencia nomenclatural de los lepidopterólogos, al igual que el resto de los animales, subyace en ejemplares tipo que se alojan en colecciones institucionales, museos y universidades. En este orden de insectos, 30 instituciones son las más importantes; algunas de las principales son el Museo Británico de Historia Natural (Londres), el Museo Nacional de Historia Natural dependiente de la Smithsonian Institution (Washington, D.C.,), el McGuire Center para Lepidoptera y Biodiversidad ligado a la Universidad de Florida (Gainesville), el Museo Americano de Historia Natural (Nueva York) y desde luego, para México, la Colección Nacional de Insectos del Instituto de Biología, UNAM (México, D.F). La selección concuerda con los catálogos de Bridges (1983, 1988), que informan acerca del depósito de tipos en los museos, entre otros aspectos de gran interés.
Morfología, posición filogenética y definición de LepidopteraLa anatomía y morfología de los lepidópteros, tanto de estados juveniles como de adultos, ha sido compilada por diversos autores. Las obras de Eaton (1988), Heppner (1998), Dias (2006) y Kristensen et al. (2007) ofrecen resúmenes y guías excelentes para este orden, con numerosas ilustraciones y claves de los grandes grupos (subórdenes, superfamilias o familias) a nivel mundial o del Neotrópico.
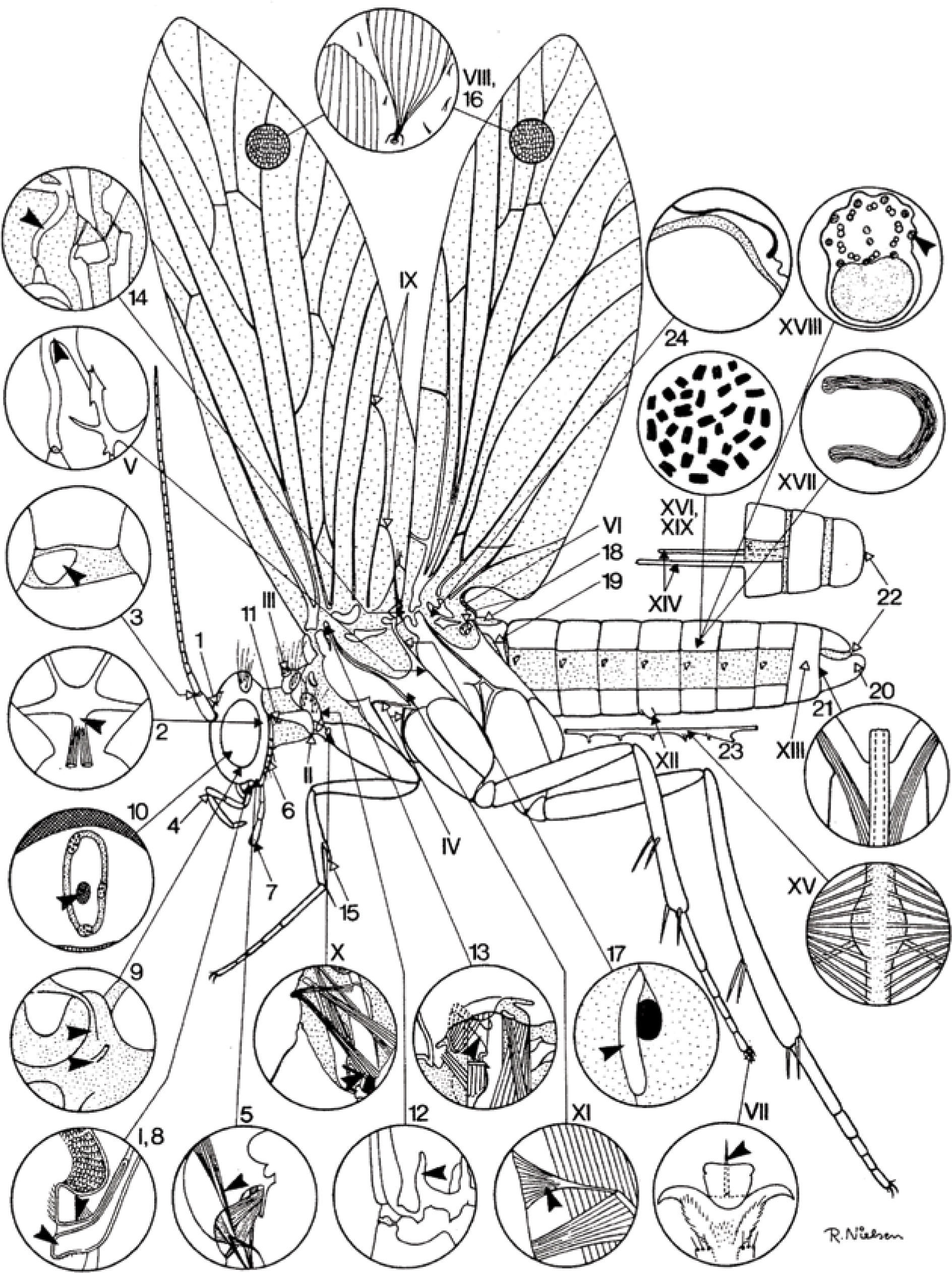
Kristensen (1984) y Kristensen y Skalski (1998) propusieron un esquema de un imago que han seguido Heppner (1998) y Kristensen et al. (2007), y que resume muy bien el plan básico del orden, incluyendo las sinapomorfías de Lepidoptera con su grupo hermano, los Trichoptera. El esquema de Heppner (1988; Fig. 1) destaca por sí solo autapomorfías y sinapomorfías cuyos trazos de detalle están basados en los trabajos originales de Hannemann (1956), Sharplin (1963), Mickoleit (1969), Suomalainen (1969), Bacceti et al. (1970), Matsuda (1970) y Common (1970), a los que se refirió Kristensen (1984).
![Diagrama de una polilla ancestral hipotética (tomado de Heppner, 1998:5, según Kristensen, 1984). Los números romanos y arábigos se refieren, respectivamente, a las autapomorfías anfiesmenopteranas y lepidopteranas enseguida listadas. Los dibujos de detalles se basan parcialmente en figuras de Hannemann (1956), Sharplin (1963), Mickoleit (1969), Suomalainen (1969), Bacceti et al. (1970), Common (1970) y Matsuda (1970). 1. Ocelo mediano revertido: perdido secundariamente. 2. Corporo-tentorium: proceso postero-mediano ausente. 3. Esc lerito intercalar del escapo antenal y membrana del pedicelo: presente. 4. Palpo maxilar: puntos de flexión presentes entre los segmentos l°-2° y 3°-4°. 5. Músculo craneoestipital largo, contiguo al músculo craneocardinal: presente. 6. Esclerito postlabial: presente. 7. Quimiorreceptores (órganos de von Rath) del segmento terminal del palpo labial: presente. 8. Músculo dorsal longitudinal del salvarium: ausente. 9. Nervio labral y ganglio conectivo frontal: separación en tritocerebro. 10. Nervus recurrens anterior al complejo retrocerebral: corre dentro de la aorta cefálica 11. Esclerito laterocerial "placa pubescente" en el ápice anterior: presente. 12. Apéndice libre del puente entre el sternum y el extremo inferior del pleuron del endoesqueleto protorácico: presente. 13. Apodema tergopleural del meso tórax desde la sutura pleural (por un músculo tergopleural): presente. 14. Brazo prescutal del mesotórax: presente. 15. Epífisis de la tibia anterior: presente (perdido secundariamente en algunos grupos). 16. Escamas en las alas: presentes (ausentes en Trichoptera primitivos). 17. Estigma respiratorio metatorácico: una abertura anterior externa única. 18. Tergum abdominal I: desesclerosado y corto, músculos dorso-longitudinales I-II perdidos. 19. Lóbulos laterales pareados del tergum abdominal I: extendidos hacia abajo o hacia atrás y articulados a los extremos anteriores del sternum II. 20. Gonopodio masculino (valva): no dividido. 21. Músculos protractores del phallus masculino: se originan dentro de los gonopodios. 22. Cerci: ausentes en ambos sexos. 23. Cordón nervioso abdominal: sólo 5 masas ganglionares presentes y los conectivos son impares (regresiones secundarias en Hepialoidea [a 6] y Neupseustidae [conectivos separados]). 24. Aorta mesotorácica: Curveada hacia el dorso del tórax (carácter revertido entre algunos Lepidoptera superiores). I. Prelabium fusionado con la hipofaringe. II. Esclerito latero-cervical agudo sobre el ángulo posterior inferior al prosternum. III.](https://static.elsevier.es/multimedia/18703453/00000085000000S1/v1_201501071628/S1870345314707122/v1_201501071628/es/main.assets/gr1.jpeg?xkr=ue/ImdikoIMrsJoerZ+w997EogCnBdOOD93cPFbanNd2Vt2E9KIXSbfPNY5VCUB4jY7b2FCQgmLmZ5CSh68s3mln+MeNOIpFC4sV5aixHWKWm+G45LQz6GIB9cwKtzB0UDtiBZhBGJohQahk78ytZf5O/V5xVsmNYkTOM+2/c7rOeabh4m5DcTxawRekcO5UO0OKIicK2AkVxjvP0+/kLsYQL6YRlnd2F+aF3Jpc5mlf81J+hrN/IlHWwL9xilKlOiSXWtvPyRD1mSWnko0BIqphd3C0S9AdBLaWah2mdKL5Rw56bHlS1VVDOncJ4HbvlUP7GZ13EIFFXmYm4EPEgA== "Diagrama de una polilla ancestral hipotética (tomado de Heppner, 1998:5, según Kristensen, 1984). Los números romanos y arábigos se refieren, respectivamente, a las autapomorfías anfiesmenopteranas y lepidopteranas enseguida listadas. Los dibujos de detalles se basan parcialmente en figuras de Hannemann (1956), Sharplin (1963), Mickoleit (1969), Suomalainen (1969), Bacceti et al. (1970), Common (1970) y Matsuda (1970). 1. Ocelo mediano revertido: perdido secundariamente. 2. Corporo-tentorium: proceso postero-mediano ausente. 3. Esc lerito intercalar del escapo antenal y membrana del pedicelo: presente. 4. Palpo maxilar: puntos de flexión presentes entre los segmentos l°-2° y 3°-4°. 5. Músculo craneoestipital largo, contiguo al músculo craneocardinal: presente. 6. Esclerito postlabial: presente. 7. Quimiorreceptores (órganos de von Rath) del segmento terminal del palpo labial: presente. 8. Músculo dorsal longitudinal del salvarium: ausente. 9. Nervio labral y ganglio conectivo frontal: separación en tritocerebro. 10. Nervus recurrens anterior al complejo retrocerebral: corre dentro de la aorta cefálica 11. Esclerito laterocerial \"placa pubescente\" en el ápice anterior: presente. 12. Apéndice libre del puente entre el sternum y el extremo inferior del pleuron del endoesqueleto protorácico: presente. 13. Apodema tergopleural del meso tórax desde la sutura pleural (por un músculo tergopleural): presente. 14. Brazo prescutal del mesotórax: presente. 15. Epífisis de la tibia anterior: presente (perdido secundariamente en algunos grupos). 16. Escamas en las alas: presentes (ausentes en Trichoptera primitivos). 17. Estigma respiratorio metatorácico: una abertura anterior externa única. 18. Tergum abdominal I: desesclerosado y corto, músculos dorso-longitudinales I-II perdidos. 19. Lóbulos laterales pareados del tergum abdominal I: extendidos hacia abajo o hacia atrás y articulados a los extremos anteriores del sternum II. 20. Gonopodio masculino (valva): no dividido. 21. Músculos protractores del phallus masculino: se originan dentro de los gonopodios. 22. Cerci: ausentes en ambos sexos. 23. Cordón nervioso abdominal: sólo 5 masas ganglionares presentes y los conectivos son impares (regresiones secundarias en Hepialoidea [a 6] y Neupseustidae [conectivos separados]). 24. Aorta mesotorácica: Curveada hacia el dorso del tórax (carácter revertido entre algunos Lepidoptera superiores). I. Prelabium fusionado con la hipofaringe. II. Esclerito latero-cervical agudo sobre el ángulo posterior inferior al prosternum. III.")
Diagrama de una polilla ancestral hipotética (tomado de Heppner, 1998:5, según Kristensen, 1984). Los números romanos y arábigos se refieren, respectivamente, a las autapomorfías anfiesmenopteranas y lepidopteranas enseguida listadas. Los dibujos de detalles se basan parcialmente en figuras de Hannemann (1956), Sharplin (1963), Mickoleit (1969), Suomalainen (1969), Bacceti et al. (1970), Common (1970) y Matsuda (1970).
1. Ocelo mediano revertido: perdido secundariamente.
2. Corporo-tentorium: proceso postero-mediano ausente.
3. Esc lerito intercalar del escapo antenal y membrana del pedicelo: presente.
4. Palpo maxilar: puntos de flexión presentes entre los segmentos l°-2° y 3°-4°.
5. Músculo craneoestipital largo, contiguo al músculo craneocardinal: presente.
6. Esclerito postlabial: presente.
7. Quimiorreceptores (órganos de von Rath) del segmento terminal del palpo labial: presente.
8. Músculo dorsal longitudinal del salvarium: ausente.
9. Nervio labral y ganglio conectivo frontal: separación en tritocerebro.
10. Nervus recurrens anterior al complejo retrocerebral: corre dentro de la aorta cefálica
11. Esclerito laterocerial "placa pubescente" en el ápice anterior: presente.
12. Apéndice libre del puente entre el sternum y el extremo inferior del pleuron del endoesqueleto protorácico: presente.
13. Apodema tergopleural del meso tórax desde la sutura pleural (por un músculo tergopleural): presente.
14. Brazo prescutal del mesotórax: presente.
15. Epífisis de la tibia anterior: presente (perdido secundariamente en algunos grupos).
16. Escamas en las alas: presentes (ausentes en Trichoptera primitivos).
17. Estigma respiratorio metatorácico: una abertura anterior externa única.
18. Tergum abdominal I: desesclerosado y corto, músculos dorso-longitudinales I-II perdidos.
19. Lóbulos laterales pareados del tergum abdominal I: extendidos hacia abajo o hacia atrás y articulados a los extremos anteriores del sternum II.
20. Gonopodio masculino (valva): no dividido.
21. Músculos protractores del phallus masculino: se originan dentro de los gonopodios.
22. Cerci: ausentes en ambos sexos.
23. Cordón nervioso abdominal: sólo 5 masas ganglionares presentes y los conectivos son impares (regresiones secundarias en Hepialoidea [a 6] y Neupseustidae [conectivos separados]).
24. Aorta mesotorácica: Curveada hacia el dorso del tórax (carácter revertido entre algunos Lepidoptera superiores).
I. Prelabium fusionado con la hipofaringe.
II. Esclerito latero-cervical agudo sobre el ángulo posterior inferior al prosternum.
III. 'Verrugas' pareadas pubescentes: presentes (supuestos precursores de los patagia en los Lepidoptera más avanzados).
IV. Episternum pterotorácico con suturas pareadas desde el origen de la sutura pleural.
V. Brazos furcales pterotorácicos secundarios fusionados con el margen posterior del epimeron adyacente.
VI. Metatórax con esclerito pubescente en la base membranosa alar debajo de la subalar.
VII. Ápice pretarsal con un pseudempodium.
VIII Alas con recubrimiento de sedas (modificadas en los Trichoptera en escamas, véase 16)
IX. Venas anales de las alas anteriores: una configuración de lazo doble-Y.
X. Músculo ventral del cuello solitario desde la coxa anterior (también manifiesto en Trichoptera primitivos).
XI. Músculo furcopleural cónico del mesotórax con extremo amplio sobre la cresta pleural.
XII. Glándulas pareadas sobre el sternum abdominal V: presente (puede estar reducido secundariamente).
XIII. Segmento genital masculino (IX) como una fusión del tergum y el sternum, formando un anillo (fusión ausente en algunos Lepidoptera como una reversión de carácter).
XIV. Márgenes anteriores de los segmentos abdominales femeninos VIII y IX con apodemas a modo de varillas: presentes (pueden estar perdidos secundariamente).
XV. Músculos ventrales del diafragma desde el cordón nervioso.
XVI. Heterogametia femenina en Lepidoptera: presente.
XVII. Esperma apyreno: presente (también en Trichoptera).
VIII. Espermatozoides con gruesos filamentos accesorios externos.
XIX. Número cromosómico alto (cromosomas probablemente holocéntricos) y oogénesis aquiasmática (véase Suomalainen, 1969).
No obstante, cabe destacar que el nombre lepidóptero (lepidos–escama, pteron–ala) que significa alas con escamas, es una generalización del taxón. No es éste el único carácter, aunque sí el más conspicuo; hay otros caracteres morfológicos (Fig. 1) que singularizan el orden. Las sedas de su grupo hermano (Trichoptera) en Lepidoptera evolucionan a escamas. Kristensen et al. (2007) destaca que la monofilia de Lepidoptera está bien apoyada en una impresionante cantidad de sinapomorfías y autapomorfías. Los Trichoptera con los Lepidoptera constituyen los Amphiesmenoptera que, a su vez, se relacionan cercanamente con los Antliophora (mecopteroideos, Siphonaptera y Diptera) para formar los Mecopterida o clado “Panorpoide”, incluido en los Endopterygota, un grupo monofilético supraordinal de los Holometabola.
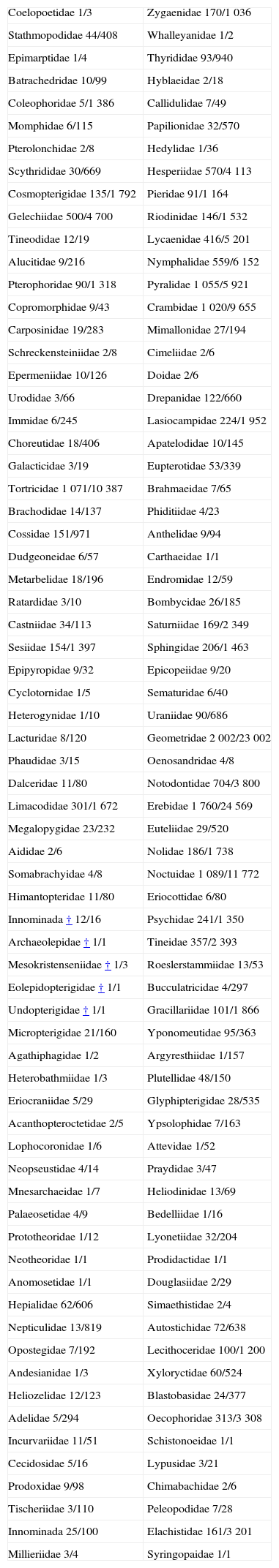
Clasificación, diversidad alfa y filogeniaUna historia general de las clasificaciones supragenéricas en los Lepidoptera se resume en los trabajos de Heppner (1998) y Kristensen et al. (2007). Las clasificaciones de suborden son relativamente estables y de consenso desde el trabajo de Common (1970), con 4 subórdenes, pero en el de super-familia (40 a 49) y familia (124 a 139), aunque similares entre diversos autores, varían ligeramente. Kristensen et al. (2007) ofrecen una discusión reciente con referencia a la clasificación alternativa de Heppner (1998). En el Apéndice se presenta la lista de familias de Kristensen et al. (2007), con los números de géneros y especies conocidas para el mundo hasta ese año; según estos autores, 45 superfamilias, 139 familias, 15 578 géneros y 157 424 especies. En 1991, 17 años antes, Heppner reconocía 146 565 especies.
Las diferencias en las clasificaciones actuales se presentan por criterios filogenéticos distintos aplicados a las diversas jerarquías entre los subgrupos de Lepidoptera (Kristensen et al., 2007), aunque todavía es considerable el desconocimiento de caracteres en estados juveniles (Heppner, 1998) y de otros caracteres para resolver y alcanzar un consenso mayor. No obstante, la incorporación reciente de análisis con bases genéticas y moleculares están ayudando a resolver algunos conflictos (Regier et al., 2009; Mutamen et al., 2010; Nieukerken et al., 2011).
Conceptos de especie y listas sistemáticasLas cifras de especies que se han propuesto, además de que varían por la concepción en la clasificación supragenérica (Heppner, 1998; Kristensen, 1998; Kristensen et al., 2007), también dependen del concepto de especie aplicado. Por ejemplo, Lamas (2000) reconoce las diferencias notables para Castniidae según la lista de Miller (1995) y de su propio trabajo del mismo año (Lamas, 1995), resultados que pueden explicarse por una comparación inadecuada de 2 listas distintas, una que se basa en el uso del concepto de especie politípica, versus otra basada en la práctica del concepto de especie biológica; tal lista sería más extensa si el concepto utilizado fuera filogenético o evolutivo. Mientras que la especie politípica subestima, la especie filogenética puede sobreestimar, en especial si en ambas hay prácticas injustificadas al valorar superficialmente las similitudes y las diferencias entre la variedad de caracteres y sus posibles interpretaciones geográfico-politípicas, biológicas o cladísticas.
A menudo el concepto de especie politípica subvalora discontinuidades morfológicas acentuadas, o bien, en ausencia de estudios relativamente detallados, conduce a reunir gran variedad de razas geográficas, aún las más disyuntas y divergentes. Una acción lumper en el sentido de Mayr et al. (1953), por defecto del concepto aplicado y la ausencia de información que permita interpretaciones más precisas. De cualquier modo su aplicación general resulta injustificada.
El criterio splitter se sigue al considerar exageradamente diferencias fenotípicas o genotípicas menores de múltiples poblaciones alopátricas, lo cual resulta en la sobrevaloración del número de especies que realmente existen. Cualquier cifra de inventario y estimación de especies, entonces, también depende de las unidades biológicas basadas en los conceptos y su aplicación. Más ejemplos y una valoración sobre ello la ofrecen Kristensen et al. (2007).
De cualquier modo, por lo general, en la gran mayoría de los grupos los criterios reproductivos están subrogados a criterios morfológicos y apenas es reciente el uso de caracteres moleculares, que lamentablemente valoran más la divergencia que la discontinuidad cualitativa. Divergencia no implica especiación, ya que ambos procesos pueden estar parcial o totalmente desacoplados en la evolución.
Con todo, es indispensable efectuar comparaciones; al provenir éstas de un mismo autor, hace posible que las diferencias conceptuales puedan atenuarse y quizá entonces se alcancen resultados más robustos, aunque siempre limitados por los conceptos de autoridad, que los alejan del fundamento de la ciencia.
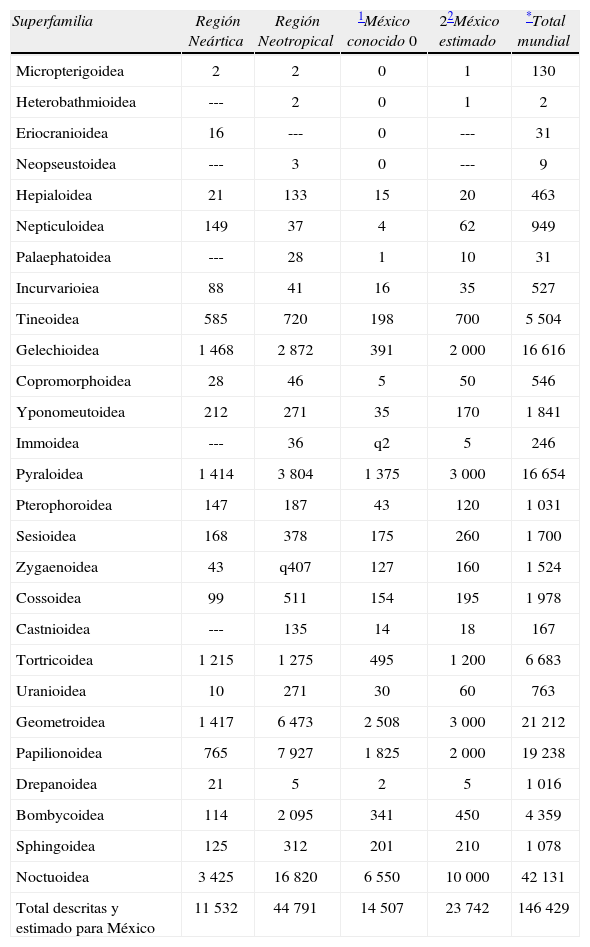
Conteos y cálculos de Lepidoptera en las regiones Neártica y Neotropical, y en MéxicoLa síntesis numérica de todos los Lepidoptera para las regiones biogeográficas del mundo por Heppner (1991, 1998), es una aproximación satisfactoria en comparaciones de diversidad. En el Cuadro 1 se resume la última de ellas (Heppner, 1998: 2). También se dan cifras para México muy similares a los números de Heppner (2002). En nuestras valoraciones se consideran hábitats, estado de desconocimiento de las superfamilias y distribuciones regionales-mundiales. No obstante, la cifra total estimada para México por Heppner (2002) es equivalente a la que aquí se alcanza. Las cifras de Heppner se formaron con base en publicaciones recientes y conocimiento de especialistas o expertos.
Riqueza mundial de las regiones Neártica y Neotropical, y de México con estimados
| Superfamilia | Región Neártica | Región Neotropical | 1México conocido 0 | 22México estimado | *Total mundial |
| Micropterigoidea | 2 | 2 | 0 | 1 | 130 |
| Heterobathmioidea | --- | 2 | 0 | 1 | 2 |
| Eriocranioidea | 16 | --- | 0 | --- | 31 |
| Neopseustoidea | --- | 3 | 0 | --- | 9 |
| Hepialoidea | 21 | 133 | 15 | 20 | 463 |
| Nepticuloidea | 149 | 37 | 4 | 62 | 949 |
| Palaephatoidea | --- | 28 | 1 | 10 | 31 |
| Incurvarioiea | 88 | 41 | 16 | 35 | 527 |
| Tineoidea | 585 | 720 | 198 | 700 | 5 504 |
| Gelechioidea | 1 468 | 2 872 | 391 | 2 000 | 16 616 |
| Copromorphoidea | 28 | 46 | 5 | 50 | 546 |
| Yponomeutoidea | 212 | 271 | 35 | 170 | 1 841 |
| Immoidea | --- | 36 | q2 | 5 | 246 |
| Pyraloidea | 1 414 | 3 804 | 1 375 | 3 000 | 16 654 |
| Pterophoroidea | 147 | 187 | 43 | 120 | 1 031 |
| Sesioidea | 168 | 378 | 175 | 260 | 1 700 |
| Zygaenoidea | 43 | q407 | 127 | 160 | 1 524 |
| Cossoidea | 99 | 511 | 154 | 195 | 1 978 |
| Castnioidea | --- | 135 | 14 | 18 | 167 |
| Tortricoidea | 1 215 | 1 275 | 495 | 1 200 | 6 683 |
| Uranioidea | 10 | 271 | 30 | 60 | 763 |
| Geometroidea | 1 417 | 6 473 | 2 508 | 3 000 | 21 212 |
| Papilionoidea | 765 | 7 927 | 1 825 | 2 000 | 19 238 |
| Drepanoidea | 21 | 5 | 2 | 5 | 1 016 |
| Bombycoidea | 114 | 2 095 | 341 | 450 | 4 359 |
| Sphingoidea | 125 | 312 | 201 | 210 | 1 078 |
| Noctuoidea | 3 425 | 16 820 | 6 550 | 10 000 | 42 131 |
| Total descritas y estimado para México | 11 532 | 44 791 | 14 507 | 23 742 | 146 429 |
En lo general se sigue a Heppner (2002), aunque las cifras de México están levemente modificadas en algunas superfamilias. Los estimados consideran hábitat, distribución mundial-regional, estado de conocimiento del grupo, y generalmente sólo de 10 a 12% de la representación de la riqueza mundial.
En este total no se consideran algunos grupos ausentes en las regiones Neártica y Neotropical. Si se considerase el total mundial sería 146 565 para el año 1990. Para el año 2007 (Apéndice) se referían 157 424 especies (Nieukerken et al., 2011). Casi 11 000 spp. 17 años después.
En México se encuentra representada la mayor parte de las superfamilias (25), cuando menos las más especiosas. En 2002, el conteo indicaba 14 507 especies y un cálculo moderado (las 255 000 especies de Heppner, 1998) alcanzó la cifra de 23 742, esto es, 9.3% de la estimación total citada. Las estimaciones de Kristensen et al. (2007) parecen sobrevaloradas (420 000 a 500 000 especies de Lepidoptera). Los Papilionoidea, una superfamilia de las mejor conocidas, alcanza un porcentaje equivalente al total de los Lepidoptera en el mundo, cuando se comparan con los Animalia (±10%).
De lo anterior y con el Cuadro 1 se pueden hacer las siguientes comparaciones y deducciones: 1) faltan por describir y denominar aproximadamente 9 300 especies de Lepidoptera para México, lo cual significa cerca de un 38% adicional a las ya conocidas; 2) según las cifras de Heppner (1998), México tiene una lepidopterofauna superior a la de toda la Región Neártica (11 532 descritas). Más aún, si se comparan las cifras estimadas de ambas áreas (México=23 742, Región Neártica=14 000). En relación con la Región Neotropical (44 791 spp.), México alcanza el 32.4% respecto a las cifras de conteo; pero sólo 26.4% de acuerdo con lo estimado (Región Neotropical 90 000 spp); 3) hacia la década de 1990, México comprendía el 9.8% de las especies de Lepidoptera registradas para el mundo y 4) hay pocas superfamilias especiosas conocidas en un 90% de sus especies, v. gr., Papilionoidea, Sphingoidea y Bombycoidea; pero de otras aún se debe hacer un mayor esfuerzo taxonómico en los próximos 15 a 30 años: Tineoidea, Gelechioidea, Yponomeutoidea, Pyraloidea, Tortricoidea y Geometroidea, y muy especialmente en Noctuoidea.
Podría decirse que algunos macroheteróceros y los ropalóceros son muy conocidos, aunque la mayor parte se han descrito con base en los imagos, y muy poco se conoce corológicamente y de los estados juveniles. Por otra parte, de las especies de distribución amplia se conocen algunos aspectos geográficos y ecológicos, pero en un país betadiverso como México (Soberón et al., 2005), se conoce poco de las especies de distribución restringida o media, que son la mayoría.
Se puede decir poco respecto a muchos tópicos de todos los Lepidoptera, pues su conocimiento es muy desigual. El simple hecho de que se hayan descrito menos del 50% de especies de algunas superfamilias hace imposible cualquier aproximación, y mucho menos generalización alguna respecto a ecología y distribución más o menos detallada. Por ello, en el siguiente apartado sólo se referirá el caso de los Papilionoidea (s. lat.), un grupo relativamente bien conocido.
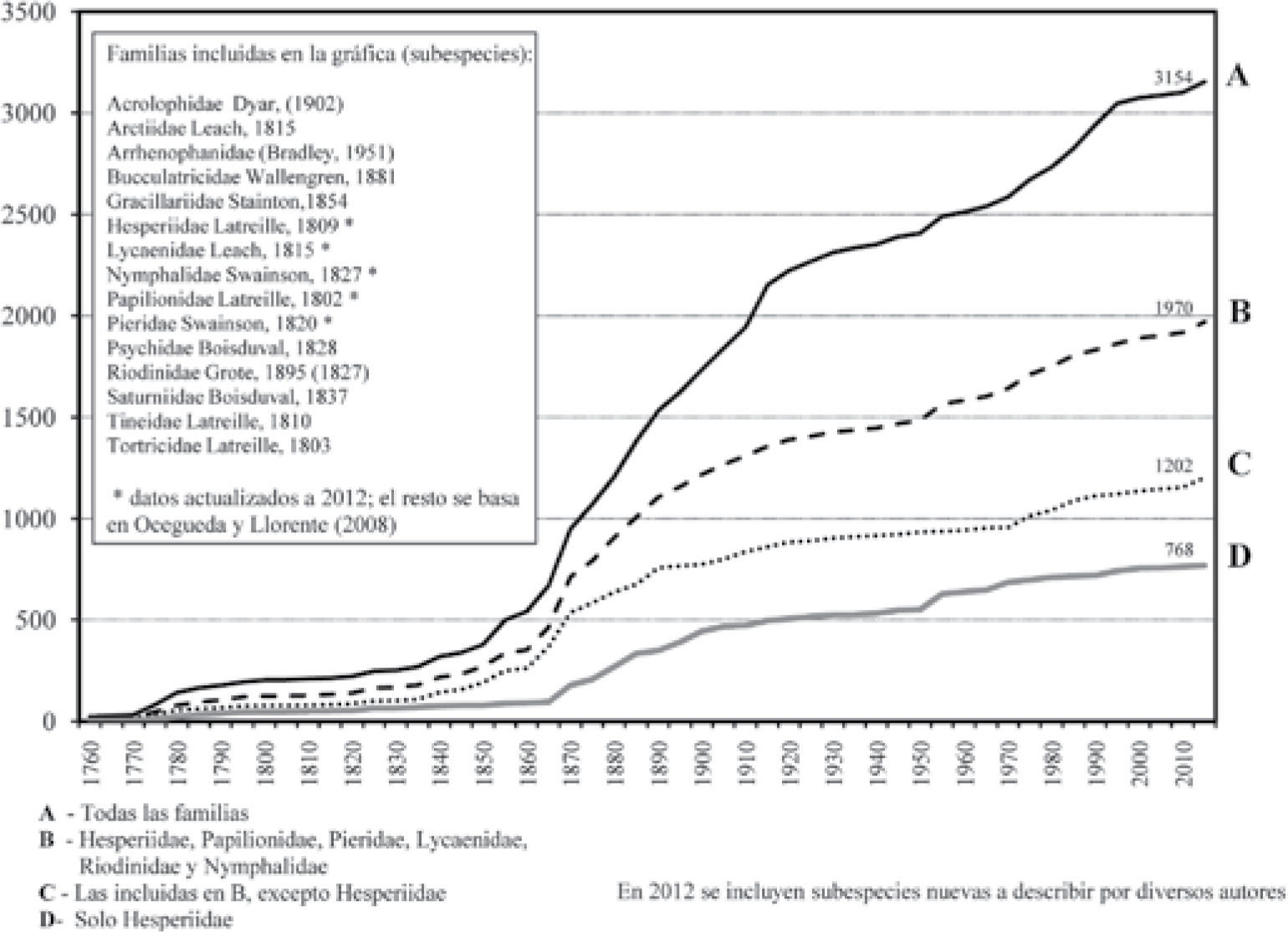
En la figura 2, se muestra el comportamiento en el tiempo de la descripción-denominación de especies de 14 familias de Lepidoptera (de bien a mal conocidas); la línea límite de la gráfica debiera estar por 4 500 especies. Por otra parte, debe considerarse que en los últimos 20 años la curva se ha desacelerado respecto a la descripción de especies, posiblemente debido a la deficiencia de recursos humanos y materiales (Lamas, 2000), pero también por el estado de inseguridad para efectuar recolecciones científicas.
Los Papilionoidea, un grupo modelo
Los antecedentes fundamentales de Papilionoidea (s. str.), el grupo mejor conocido para México en los Lepidoptera, se han citado en varios trabajos de Llorente y Luis (1993, 1998), Llorente et al. (1993, 1996b, 2006a), Luis et al. (2000, 2003, 2005) y Michán et al. (2004). Si se incluyese Hesperiidae como familia de Papilionoidea (s. lat.) se sumarían los trabajos de Llorente et al. (1990), Luis et al. (2003) y Warren (2000). Las publicaciones en la Revista de la Sociedad Mexicana de Lepidopterología (Maza et al., 1989, 1991) en el contexto nacional y los trabajos de Lamas (1981, 1995, 2000, 2004, 2008), Heppner (1998) y Mielke (2005) en el contexto regional y mundial, son referentes necesarios si se quieren comparar y contrastar conceptos o criterios en diversidad de las mariposas de este país en relación con las regiones Neártica y Neotropical o con el mundo. Desde luego, los números citados por Shields (1989) con base en los catálogos de Bridges y otras publicaciones más antiguas, así como los citados por Brown (1982) y Robbins (1982), también basados en publicaciones anteriores a la década de 1980, han quedado obsoletos. Una discusión interesante sobre el número de especies en el mundo y por regiones es la de Lamas (2008), quien contrasta estimaciones de Shields (1989), Robbins (1982) y Heppner (1991) para cada familia de Papilionoidea (s. lat). En su análisis, Lamas (2008) valora la opinión de Descimon y Mallet (2009) sobre la aplicación de conceptos de especie a las mariposas de Europa.
En algunos escritos sobre la historia de la entomología y la lepidopterología de México (Michán y Llorente, 2002; Michán et al., 2004, 2005) y en diversos libros por grupo o por familia, o en las síntesis de Llorente et al. (1997), Luis et al. (2003, 2010) y Vargas et al. (2008) se han tratado varios aspectos de interés relacionados con los periodos históricos, los recolectores, los descriptores de especies, las colecciones o museos, las principales sociedades científicas e instituciones en México y en el extranjero. Se citan las publicaciones relevantes (revistas y libros), así como los itinerarios, las localidades o áreas de mayor riqueza o endemismo y desde luego, la base de datos (mariposa) que ha permitido efectuar diversas síntesis de conocimiento del grupo (Luis et al., 2003, 2005; Llorente et al., 2006a) de naturaleza taxonómica y biogeográfica, durante los últimos 10 años.
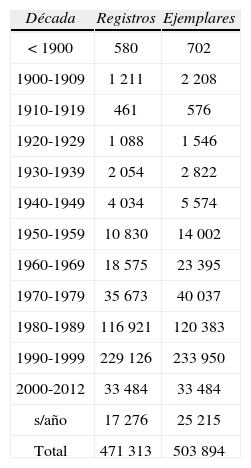
Ritmo cronológico de incremento de las especies-sub-especies y ejemplares en coleccionesLa figura 2 ilustra abreviadamente el comportamiento temporal de la sistemática en el nivel de subespecie, a partir de la lista taxonómica de Llorente et al. (2006a), actualizada a 2012. Para Papilionoidea (s. str.), Michán et al. (2004, 2005) construyeron curvas parciales (por periodo o subtaxones). Las tabulaciones para ejemplares en función del tiempo (megabase mariposa, ca. 500 000 ejemplares) las efectuaron Luis et al. (2005); el Cuadro 2 muestra una actualización a este año (abril 2012).
Registros y ejemplares por década contenidos en la megabase MARIPOSA, actualizados a 2011 (Luis et al., 2005)*
| Década | Registros | Ejemplares |
| <1900 | 580 | 702 |
| 1900-1909 | 1 211 | 2 208 |
| 1910-1919 | 461 | 576 |
| 1920-1929 | 1 088 | 1 546 |
| 1930-1939 | 2 054 | 2 822 |
| 1940-1949 | 4 034 | 5 574 |
| 1950-1959 | 10 830 | 14 002 |
| 1960-1969 | 18 575 | 23 395 |
| 1970-1979 | 35 673 | 40 037 |
| 1980-1989 | 116 921 | 120 383 |
| 1990-1999 | 229 126 | 233 950 |
| 2000-2012 | 33 484 | 33 484 |
| s/año | 17 276 | 25 215 |
| Total | 471 313 | 503 894 |
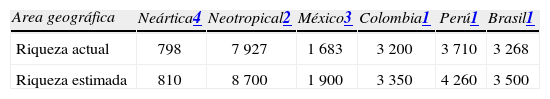
Los trabajos previos de Llorente y Luis (1993, 1998) estimaban 2 200 especies de Papilionoidea (s. lat.) de México. Un recuento detallado, al considerar la base de datos mariposa, que contiene el examen de las colecciones más importantes, además de todas las publicaciones y las opiniones de varios expertos, alcanza una cifra cercana a las 1 700 especies y un estimación de 1 900. Si se considerase la aplicación del concepto de especie evolutiva a los Rhopalocera de México, el número estimado superaría las 2 000 especies. El descubrimiento de endémicos nuevos de distribución restringida en México (estenoendémicos), así como el análisis detallado de complejos de especies crípticas, serán la fuente principal para completar la lista estimada. El Cuadro 3 muestra una tabulación del número de especies de México vs. otras regiones o países.
Riqueza de Papilionoidea (s. lat.) en algunas áreas geográficas
| Area geográfica | Neártica4 | Neotropical2 | México3 | Colombia1 | Perú1 | Brasil1 |
| Riqueza actual | 798 | 7 927 | 1 683 | 3 200 | 3 710 | 3 268 |
| Riqueza estimada | 810 | 8 700 | 1 900 | 3 350 | 4 260 | 3 500 |
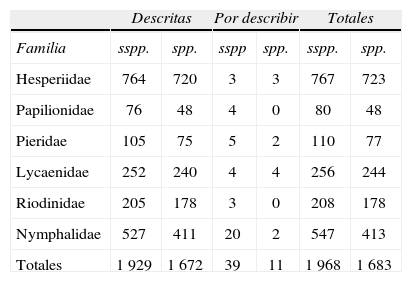
En el Cuadro 4 se tabulan las subespecies y especies de todas las familias de Papilionoidea (s. lat.) de México, descritas o por describir, ya con ejemplares como evidencia. En la figura 2, se utilizan los números: 1 968 sspp. de 1 683 spp. La mayoría de las especies y subespecies por describir se citan en el catálogo de Lamas (2004); algunas están en la colección del Museo de Zoología de la Facultad de Ciencias y otras más se citan en diversas publicaciones o están en prensa.
Subespecies y especies descritas y por describir de los Rhopalocera de México
| Descritas | Por describir | Totales | ||||
| Familia | sspp. | spp. | sspp | spp. | sspp. | spp. |
| Hesperiidae | 764 | 720 | 3 | 3 | 767 | 723 |
| Papilionidae | 76 | 48 | 4 | 0 | 80 | 48 |
| Pieridae | 105 | 75 | 5 | 2 | 110 | 77 |
| Lycaenidae | 252 | 240 | 4 | 4 | 256 | 244 |
| Riodinidae | 205 | 178 | 3 | 0 | 208 | 178 |
| Nymphalidae | 527 | 411 | 20 | 2 | 547 | 413 |
| Totales | 1 929 | 1 672 | 39 | 11 | 1 968 | 1 683 |
Diversos temas faunísticos con marcos geográficos variados se han desarrollado con Papilionoidea. Entre los principales están Península de Baja California: Brown et al. (1992); Nayarit: Llorente et al. (2004a); Jalisco: Vargas et al. (1996), Warren et al. (1996) y Llorente et al. (1996c); Colima: Llorente et al. (1996d) y Warren et al. (1998); Estado de México: Hernández-Mejía et al. (2008a); Morelos: Luna et al. (2012); Veracruz: Luis et al. (1996, 2011a) y Hernández-Baz et al. (2010); Oaxaca: Luis et al. (2004); Quintana Roo: Maza y Gutiérrez (1994); Chiapas: Maza y Maza (1993) y Luis et al. (2011b). Así también, temas por transectos en áreas complejas (v. gr,Luis y Llorente, 1990, 1993; Luis et al., 1991, 1999; Vargas et al., 1991, 1994, 1999; Monteagudo et al., 2001); por unidades vegetacionales (Salinas et al., 2004; Maya et al., 2005); por áreas protegidas (v. gr.,Beutelspacher, 1982a; Raguso y Llorente, 1991, 1997; Llorente y Luis, 1993; Bizuet et al., 2001; Díaz-Batres et al., 2001; Pozo et al., 2003; Luna y Llorente, 2004; Luna et al., 2008), y por sectores fisiográficos o unidades geomorfológicas (Hoffmann, 1933; Ross, 1964-1967; Beutelspacher, 1975; Maza y Maza, 1985, 1988; Llorente et al., 1986; Hernández-Mejía et al., 2008b; Pozo et al., 2008; Hernández-Baz et al., 2010; Luis et al., 2010; Luna et al., 2010), entre los principales temas geográficos. Posiblemente se han desarrollado más de un centenar de tesis de licenciatura o maestría (biología), sobre el tema de faunística de mariposas de diversos lugares o áreas de México, y pocas de ellas han sido publicadas.
El aspecto geográfico es un tema inaugurado por Godman y Salvin (1878-1901), pero sistematizado y sintetizado por Hoffmann (1940, 1941), y fue continuado más tarde por Vázquez (1942) y Beutelspacher (1982b) en el Instituto de Biología (UNAM). Con la formación de la Sociedad Mexicana de Lepidopterología se dio nuevo impulso a los trabajos ropalocerofaunísticos o taxonómicos en las zonas de mayor riqueza y endemismo, en especial de la mitad sur de México. Se pueden consultar varios trabajos en el Boletín y la Revista de dicha Sociedad entre 1975 y 1996 (más de 16 volúmenes).
Aparte del ensayo de Hoffmann (1936), los análisis biogeográficos son más recientes y han servido para caracterizar patrones de distribución, provincias bióticas o regiones ecológicas (Oñate et al., 2006 para Papilionidae; Llorente et al., 2006b, para Pieridae; Luis et al., 2006 y Vargas et al., 2006, para 7 subfamilias de Nymphalidae y para Charaxinae Maya et al., 2009, 2011). Una visión resumida puede consultarse en los trabajos de Luis et al. (2003, 2005), Pozo et al. (2005), Llorente et al. (2006a) y Morrone y Llorente (2006). Sobre áreas de refugios pleistocénicos están los trabajos de Maza y Maza (1993) y Maza et al. (1995a, b). También en técnicas de interés ecogeográfico los Papilionoidea han sido un modelo importante: Soberón y Llorente (1993), Soberón et al. (2000, 2005) y Pozo et al. (2008). Luis et al. (2005) ofrecen una visión sucinta del uso de una megabase de datos (500 000 ejemplares de Papilionoidea) en aspectos de regionalización biogeográfica (provincias, ecorregiones, distribución estatal, densidad de representación biogeográfica de ejemplares y especies), así como de relaciones “cladísticas” entre unidades bióticas: interrelación de endemismos.
Al considerar por familias el endemismo específico, se obtienen los siguientes porcentajes: Hesperiidae 15.83% (Eudaminae 12 spp., Pyrginae 26 spp., Heteropterinae 23 spp. y Hesperiinae 74 spp.), Papilionidae 6.3% (Baroniinae 1 sp. y Papilioninae 3 spp.), Pieridae 12.98% (Dismorphiinae 2 spp.), Lycaenidae 5.27% (Theclinae 8 spp.), Riodinidae 16.29% (Euselasiinae 3 spp. y Riodininae 26 spp.) y Nymphalidae 11.86% (Ithomiinae 1 sp., Morphinae 2 spp., Satyrinae 25 spp., Charaxinae 2 spp., Biblidinae 4 spp., Nymphalinae 12 spp. y Limenitidinae 3 spp.).
El porcentaje para Papilionoidea (s. lat.) en México aumenta a 22.2% si se considera el nivel subespecífico. En este nivel, se alcanzan las siguientes cifras de endémicas: Hesperiidae 155 sspp., Papilionidae 32 sspp., Pieridae 37 sspp., Lycaenidae 10 sspp., Riodinidae 44 sspp. y Nymphalidae 159 sspp. Compárense estas cifras con el Cuadro 4, en el que se ofrecen las cifras totales.
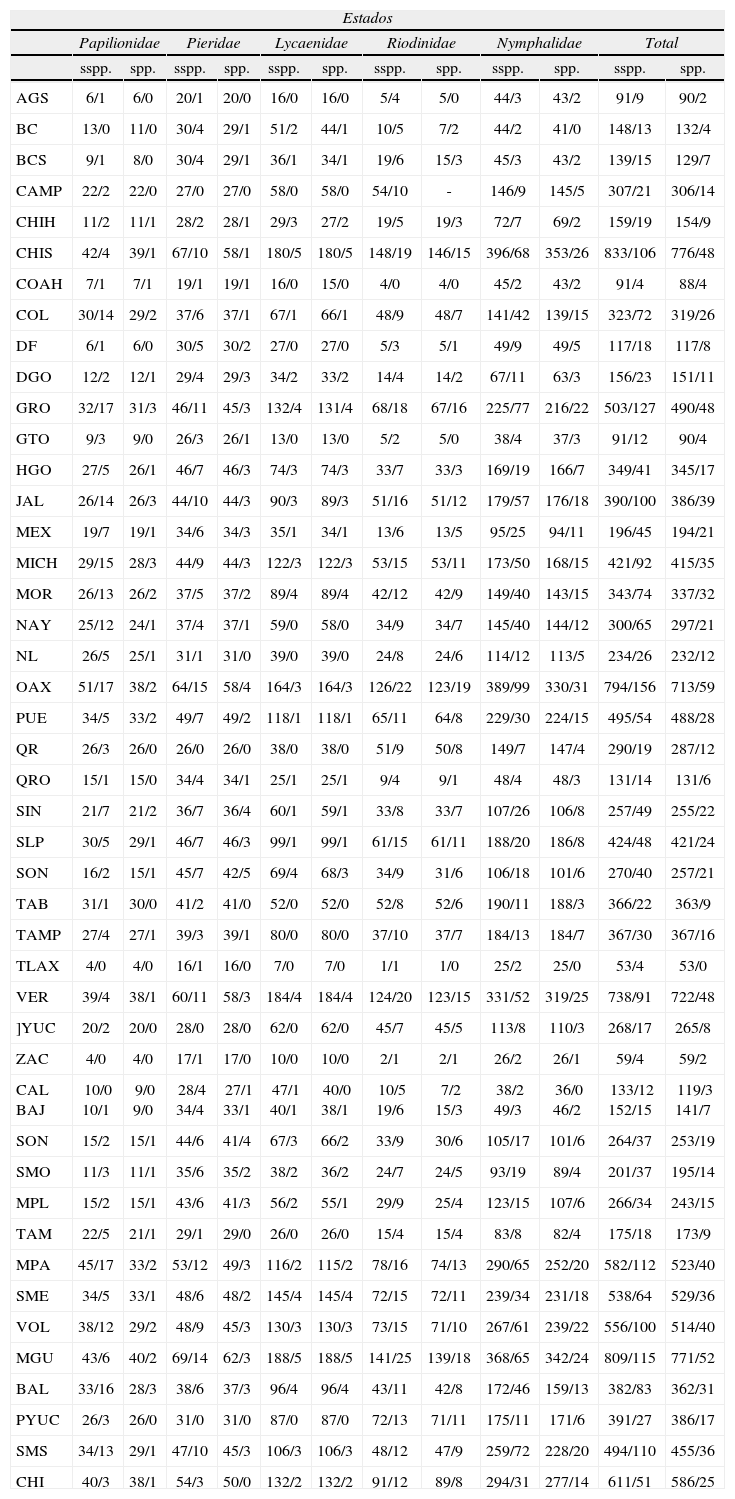
Riqueza, endemismo y su distribuciónDiversos autores han descrito decenas de endémicas para distintas regiones de México durante los últimos 40 años. Listas actualizadas y recientes de las Papilionoidea endémicas de México las ofrecen explícitamente Llorente y Luis (1993, 1998), Luis et al. (2000, 2003) y Michán et al. (2004), e implícitamente en el trabajo de Llorente et al. (2006a). La tabulación de este último trabajo, con actualización a 2012, es la base principal para el cuadro resumido de riqueza y endemismo que aquí se presenta para los estados y provincias bióticas de México (Cuadro 5). Una lista revisada y actualizada de los endemitas de Papilionoidea (s. lat.) de este país, actualmente en preparación, se presentará próximamente; aquí sólo se considera para las cifras actualizadas.
Riqueza y representación del endemismo (subespecies-especies) por familias en los estados y las provincias biogeográficas de México*
| Estados | ||||||||||||
| Papilionidae | Pieridae | Lycaenidae | Riodinidae | Nymphalidae | Total | |||||||
| sspp. | spp. | sspp. | spp. | sspp. | spp. | sspp. | spp. | sspp. | spp. | sspp. | spp. | |
| AGS | 6/1 | 6/0 | 20/1 | 20/0 | 16/0 | 16/0 | 5/4 | 5/0 | 44/3 | 43/2 | 91/9 | 90/2 |
| BC | 13/0 | 11/0 | 30/4 | 29/1 | 51/2 | 44/1 | 10/5 | 7/2 | 44/2 | 41/0 | 148/13 | 132/4 |
| BCS | 9/1 | 8/0 | 30/4 | 29/1 | 36/1 | 34/1 | 19/6 | 15/3 | 45/3 | 43/2 | 139/15 | 129/7 |
| CAMP | 22/2 | 22/0 | 27/0 | 27/0 | 58/0 | 58/0 | 54/10 | - | 146/9 | 145/5 | 307/21 | 306/14 |
| CHIH | 11/2 | 11/1 | 28/2 | 28/1 | 29/3 | 27/2 | 19/5 | 19/3 | 72/7 | 69/2 | 159/19 | 154/9 |
| CHIS | 42/4 | 39/1 | 67/10 | 58/1 | 180/5 | 180/5 | 148/19 | 146/15 | 396/68 | 353/26 | 833/106 | 776/48 |
| COAH | 7/1 | 7/1 | 19/1 | 19/1 | 16/0 | 15/0 | 4/0 | 4/0 | 45/2 | 43/2 | 91/4 | 88/4 |
| COL | 30/14 | 29/2 | 37/6 | 37/1 | 67/1 | 66/1 | 48/9 | 48/7 | 141/42 | 139/15 | 323/72 | 319/26 |
| DF | 6/1 | 6/0 | 30/5 | 30/2 | 27/0 | 27/0 | 5/3 | 5/1 | 49/9 | 49/5 | 117/18 | 117/8 |
| DGO | 12/2 | 12/1 | 29/4 | 29/3 | 34/2 | 33/2 | 14/4 | 14/2 | 67/11 | 63/3 | 156/23 | 151/11 |
| GRO | 32/17 | 31/3 | 46/11 | 45/3 | 132/4 | 131/4 | 68/18 | 67/16 | 225/77 | 216/22 | 503/127 | 490/48 |
| GTO | 9/3 | 9/0 | 26/3 | 26/1 | 13/0 | 13/0 | 5/2 | 5/0 | 38/4 | 37/3 | 91/12 | 90/4 |
| HGO | 27/5 | 26/1 | 46/7 | 46/3 | 74/3 | 74/3 | 33/7 | 33/3 | 169/19 | 166/7 | 349/41 | 345/17 |
| JAL | 26/14 | 26/3 | 44/10 | 44/3 | 90/3 | 89/3 | 51/16 | 51/12 | 179/57 | 176/18 | 390/100 | 386/39 |
| MEX | 19/7 | 19/1 | 34/6 | 34/3 | 35/1 | 34/1 | 13/6 | 13/5 | 95/25 | 94/11 | 196/45 | 194/21 |
| MICH | 29/15 | 28/3 | 44/9 | 44/3 | 122/3 | 122/3 | 53/15 | 53/11 | 173/50 | 168/15 | 421/92 | 415/35 |
| MOR | 26/13 | 26/2 | 37/5 | 37/2 | 89/4 | 89/4 | 42/12 | 42/9 | 149/40 | 143/15 | 343/74 | 337/32 |
| NAY | 25/12 | 24/1 | 37/4 | 37/1 | 59/0 | 58/0 | 34/9 | 34/7 | 145/40 | 144/12 | 300/65 | 297/21 |
| NL | 26/5 | 25/1 | 31/1 | 31/0 | 39/0 | 39/0 | 24/8 | 24/6 | 114/12 | 113/5 | 234/26 | 232/12 |
| OAX | 51/17 | 38/2 | 64/15 | 58/4 | 164/3 | 164/3 | 126/22 | 123/19 | 389/99 | 330/31 | 794/156 | 713/59 |
| PUE | 34/5 | 33/2 | 49/7 | 49/2 | 118/1 | 118/1 | 65/11 | 64/8 | 229/30 | 224/15 | 495/54 | 488/28 |
| QR | 26/3 | 26/0 | 26/0 | 26/0 | 38/0 | 38/0 | 51/9 | 50/8 | 149/7 | 147/4 | 290/19 | 287/12 |
| QRO | 15/1 | 15/0 | 34/4 | 34/1 | 25/1 | 25/1 | 9/4 | 9/1 | 48/4 | 48/3 | 131/14 | 131/6 |
| SIN | 21/7 | 21/2 | 36/7 | 36/4 | 60/1 | 59/1 | 33/8 | 33/7 | 107/26 | 106/8 | 257/49 | 255/22 |
| SLP | 30/5 | 29/1 | 46/7 | 46/3 | 99/1 | 99/1 | 61/15 | 61/11 | 188/20 | 186/8 | 424/48 | 421/24 |
| SON | 16/2 | 15/1 | 45/7 | 42/5 | 69/4 | 68/3 | 34/9 | 31/6 | 106/18 | 101/6 | 270/40 | 257/21 |
| TAB | 31/1 | 30/0 | 41/2 | 41/0 | 52/0 | 52/0 | 52/8 | 52/6 | 190/11 | 188/3 | 366/22 | 363/9 |
| TAMP | 27/4 | 27/1 | 39/3 | 39/1 | 80/0 | 80/0 | 37/10 | 37/7 | 184/13 | 184/7 | 367/30 | 367/16 |
| TLAX | 4/0 | 4/0 | 16/1 | 16/0 | 7/0 | 7/0 | 1/1 | 1/0 | 25/2 | 25/0 | 53/4 | 53/0 |
| VER | 39/4 | 38/1 | 60/11 | 58/3 | 184/4 | 184/4 | 124/20 | 123/15 | 331/52 | 319/25 | 738/91 | 722/48 |
| ]YUC | 20/2 | 20/0 | 28/0 | 28/0 | 62/0 | 62/0 | 45/7 | 45/5 | 113/8 | 110/3 | 268/17 | 265/8 |
| ZAC | 4/0 | 4/0 | 17/1 | 17/0 | 10/0 | 10/0 | 2/1 | 2/1 | 26/2 | 26/1 | 59/4 | 59/2 |
| CAL BAJ | 10/0 10/1 | 9/0 9/0 | 28/4 34/4 | 27/1 33/1 | 47/1 40/1 | 40/0 38/1 | 10/5 19/6 | 7/2 15/3 | 38/2 49/3 | 36/0 46/2 | 133/12 152/15 | 119/3 141/7 |
| SON | 15/2 | 15/1 | 44/6 | 41/4 | 67/3 | 66/2 | 33/9 | 30/6 | 105/17 | 101/6 | 264/37 | 253/19 |
| SMO | 11/3 | 11/1 | 35/6 | 35/2 | 38/2 | 36/2 | 24/7 | 24/5 | 93/19 | 89/4 | 201/37 | 195/14 |
| MPL | 15/2 | 15/1 | 43/6 | 41/3 | 56/2 | 55/1 | 29/9 | 25/4 | 123/15 | 107/6 | 266/34 | 243/15 |
| TAM | 22/5 | 21/1 | 29/1 | 29/0 | 26/0 | 26/0 | 15/4 | 15/4 | 83/8 | 82/4 | 175/18 | 173/9 |
| MPA | 45/17 | 33/2 | 53/12 | 49/3 | 116/2 | 115/2 | 78/16 | 74/13 | 290/65 | 252/20 | 582/112 | 523/40 |
| SME | 34/5 | 33/1 | 48/6 | 48/2 | 145/4 | 145/4 | 72/15 | 72/11 | 239/34 | 231/18 | 538/64 | 529/36 |
| VOL | 38/12 | 29/2 | 48/9 | 45/3 | 130/3 | 130/3 | 73/15 | 71/10 | 267/61 | 239/22 | 556/100 | 514/40 |
| MGU | 43/6 | 40/2 | 69/14 | 62/3 | 188/5 | 188/5 | 141/25 | 139/18 | 368/65 | 342/24 | 809/115 | 771/52 |
| BAL | 33/16 | 28/3 | 38/6 | 37/3 | 96/4 | 96/4 | 43/11 | 42/8 | 172/46 | 159/13 | 382/83 | 362/31 |
| PYUC | 26/3 | 26/0 | 31/0 | 31/0 | 87/0 | 87/0 | 72/13 | 71/11 | 175/11 | 171/6 | 391/27 | 386/17 |
| SMS | 34/13 | 29/1 | 47/10 | 45/3 | 106/3 | 106/3 | 48/12 | 47/9 | 259/72 | 228/20 | 494/110 | 455/36 |
| CHI | 40/3 | 38/1 | 54/3 | 50/0 | 132/2 | 132/2 | 91/12 | 89/8 | 294/31 | 277/14 | 611/51 | 586/25 |
El orden de los estados es alfabético; el de las provincias, de N a S y de O a E.Abreviaturas de los estados: AGS, Aguascalientes; BC, Baja California; BCS, Baja California Sur; CAMP, Campeche; CHIH, Chihuahua; CHIS, Chiapas; COAH, Coahuila; COL, Colima; DF, Distrito Federal; DGO, Durango; GRO, Guerrero; GTO, Guanajuato; HGO, Hidalgo; JAL, Jalisco; MEX, Estado de México; MICH, Mchoacán; MOR, Morelos; NAY, Nayarit; NL, Nuevo León; OAX, Oaxaca; PUE, Puebla; QR, Quintana Roo; QRO, Querétaro; SIN, Sinaloa; SLP, San Luis Potosí; SON, Sonora; TAB, Tabasco; TAMP, Tamaulipas; TLAX, Tlaxcala; VER, Veracruz; YUC, Yucatán; ZAC, Zacatecas.Abreviaturas de las provincias: CAL, California; BAJ, Baja California; SON, Sonora; SMO, Sierra Madre Occidental; MPL, Altiplano Mexicano; TAM, Tamaulipas; MPA, Costa del Pacífico; SME, Sierra Madre Oriental; VOL, Eje Neovolcánico; MGU, Costa del Golfo de México; BAL, Depresión del Balsas; PYUC, Península de Yucatán; SMS, Sierra Madre del Sur; CHI, Chiapas(sensuMorrone et al., 2002).
El endemismo de Papilionoidea de México es aproximadamente de un 14% de las especies (235 endémicas de un total de 1 683), pero una decena de géneros también lo son, algunos de ellos monotípicos y posiblemente paleoendémicos relictos como Aegiale, Baronia, Prestonia y Eucheira. Muchos otros géneros se han diversificado primaria o secundariamente en las montañas, desiertos y bosques tropicales de México, v. gr, Bolla, Dalla, Piruna, Agathymus, Amblyscirtes, Polites, Poanes, Paratrytone, Neposa, Quasimellana, Atrytonopsis en Hesperioidea; Catasticta, Euselasia, Calephelis, Caria, Emesis, Cyllopsis, Paramacera, Memphis, Polygonia, Chlosyne y Adelpha en Papilionoidea.
Dado que el endemismo puede ser compartido con el sur de los Estados Unidos o con el norte de América Central, Llorente y Luis (1993, 1998) propusieron el término cuasiendémico (quasiendemics), pues la mayor parte de la distribución (70-80%) es en México para más del 50% de especies de los taxones. Este término se prefiere a los acuñados por Rzedowski (1978), como Mega México, porque enfatiza el endemismo y los taxones en territorios actuales y reales. No obstante, las áreas naturales son más cercanas a lo propuesto por Rzedowski (1978), aunque los nombres usados por él se rechazan en Centroamérica por su connotación “expansionista” (Cano, com. pers.).
Por lo general, los grupos estenoecos o estenotópicos de las montañas (bosques mesófilos, mixtos de pino-encino y coníferas), los desiertos o semidesiertos y los bosques tropicales (en especial subcaducifolios y caducifolios) de México, han sido escenarios de diversificaciones de géneros, grupos de especies o diferenciación de razas geográficas. En los bosques tropicales perennifolios lo más frecuente es una diferenciación subespecífica, a menudo compartida con áreas del norte de Centroamérica. Algo equivalente ocurre con las áreas desérticas del norte de México y los bosques mesófilos del sur de México, pues son continuas con Estados Unidos y Guatemala, respectivamente.
Un análisis sucinto del Cuadro 5 revela que la representación del endemismo no guarda dependencia absoluta con la riqueza; sea por estados o provincias biogeográficas. Por ejemplo, el orden de mayor riqueza es: Chiapas, Oaxaca y Veracruz, mientras que el del endemismo es Oaxaca, Chiapas y Veracruz. El endemismo está mejor representado en los estados de la vertiente del Pacífico que en los estados de la vertiente del Atlántico, no así la riqueza.
Riqueza y endemismo por familia entre los distintos estados y provincias biogeográficas guardan distintos órdenes de relación. Por ejemplo, Guerrero es el cuarto en riqueza de Papilionidae, pero con la mayor proporción de representación de endémicas. Sin embargo otras familias no cumplen necesariamente esta afirmación.
Cuando se examina las unidades naturales respecto a las unidades políticas, el orden de riqueza y endemismo deja de cumplirse, pues la provincia Costa del Golfo de México supera notablemente en riqueza a la provincia de Chiapas y la de Costa del Pacífico. No obstante, las provincias con mayor riqueza y endemismo son 6, que en orden son: MGU, CHI, MPA, SME, VOL y SMS (véase Cuadro 5). Sin embargo, el muestreo, la extensión y la variedad fisiográfica, climática y vegetacional es distinta para cada provincia.
Este resumen (Cuadro 5) requiere detallarse y complementarse con un análisis comparativo de la distribución del endemismo y la riqueza por pisos altitudinales y ecosistemas o tipos de vegetación; que se encuentra en desarrollo por los autores del presente trabajo. En dicho análisis se comprenderá el tamaño del área de superficie de cada unidad natural.
A la Facultad de Ciencias, UNAM y a la DGAPA, a través de los proyectos PAPIIT IN214212 y PAPIME PE200812; al financiamiento de CONACyT 083237 y a diversos proyectos de CONABIO, además del JF018, que destinaron recursos en varias etapas de la formación de la Megabase mariposa.
Familias de Lepidoptera. El orden de las familias es aproximadamente estratigráfico o filogenético, según Kristensen (1998), Kristensen et al. (2007), Regier et al. (2009) y Mutamen et al. (2010). Nieukerken et al. (2011) recopilaron los números aquí tabulados y sintetizaron la clasificación.
| Coelopoetidae 1/3 | Zygaenidae 170/1 036 |
| Stathmopodidae 44/408 | Whalleyanidae 1/2 |
| Epimarptidae 1/4 | Thyrididae 93/940 |
| Batrachedridae 10/99 | Hyblaeidae 2/18 |
| Coleophoridae 5/1 386 | Callidulidae 7/49 |
| Momphidae 6/115 | Papilionidae 32/570 |
| Pterolonchidae 2/8 | Hedylidae 1/36 |
| Scythrididae 30/669 | Hesperiidae 570/4 113 |
| Cosmopterigidae 135/1 792 | Pieridae 91/1 164 |
| Gelechiidae 500/4 700 | Riodinidae 146/1 532 |
| Tineodidae 12/19 | Lycaenidae 416/5 201 |
| Alucitidae 9/216 | Nymphalidae 559/6 152 |
| Pterophoridae 90/1 318 | Pyralidae 1 055/5 921 |
| Copromorphidae 9/43 | Crambidae 1 020/9 655 |
| Carposinidae 19/283 | Mimallonidae 27/194 |
| Schreckensteiniidae 2/8 | Cimeliidae 2/6 |
| Epermeniidae 10/126 | Doidae 2/6 |
| Urodidae 3/66 | Drepanidae 122/660 |
| Immidae 6/245 | Lasiocampidae 224/1 952 |
| Choreutidae 18/406 | Apatelodidae 10/145 |
| Galacticidae 3/19 | Eupterotidae 53/339 |
| Tortricidae 1 071/10 387 | Brahmaeidae 7/65 |
| Brachodidae 14/137 | Phiditiidae 4/23 |
| Cossidae 151/971 | Anthelidae 9/94 |
| Dudgeoneidae 6/57 | Carthaeidae 1/1 |
| Metarbelidae 18/196 | Endromidae 12/59 |
| Ratardidae 3/10 | Bombycidae 26/185 |
| Castniidae 34/113 | Saturniidae 169/2 349 |
| Sesiidae 154/1 397 | Sphingidae 206/1 463 |
| Epipyropidae 9/32 | Epicopeiidae 9/20 |
| Cyclotornidae 1/5 | Sematuridae 6/40 |
| Heterogynidae 1/10 | Uraniidae 90/686 |
| Lacturidae 8/120 | Geometridae 2 002/23 002 |
| Phaudidae 3/15 | Oenosandridae 4/8 |
| Dalceridae 11/80 | Notodontidae 704/3 800 |
| Limacodidae 301/1 672 | Erebidae 1 760/24 569 |
| Megalopygidae 23/232 | Euteliidae 29/520 |
| Aididae 2/6 | Nolidae 186/1 738 |
| Somabrachyidae 4/8 | Noctuidae 1 089/11 772 |
| Himantopteridae 11/80 | Eriocottidae 6/80 |
| Innominada † 12/16 | Psychidae 241/1 350 |
| Archaeolepidae † 1/1 | Tineidae 357/2 393 |
| Mesokristenseniidae † 1/3 | Roeslerstammiidae 13/53 |
| Eolepidopterigidae † 1/1 | Bucculatricidae 4/297 |
| Undopterigidae † 1/1 | Gracillariidae 101/1 866 |
| Micropterigidae 21/160 | Yponomeutidae 95/363 |
| Agathiphagidae 1/2 | Argyresthiidae 1/157 |
| Heterobathmiidae 1/3 | Plutellidae 48/150 |
| Eriocraniidae 5/29 | Glyphipterigidae 28/535 |
| Acanthopteroctetidae 2/5 | Ypsolophidae 7/163 |
| Lophocoronidae 1/6 | Attevidae 1/52 |
| Neopseustidae 4/14 | Praydidae 3/47 |
| Mnesarchaeidae 1/7 | Heliodinidae 13/69 |
| Palaeosetidae 4/9 | Bedelliidae 1/16 |
| Prototheoridae 1/12 | Lyonetiidae 32/204 |
| Neotheoridae 1/1 | Prodidactidae 1/1 |
| Anomosetidae 1/1 | Douglasiidae 2/29 |
| Hepialidae 62/606 | Simaethistidae 2/4 |
| Nepticulidae 13/819 | Autostichidae 72/638 |
| Opostegidae 7/192 | Lecithoceridae 100/1 200 |
| Andesianidae 1/3 | Xyloryctidae 60/524 |
| Heliozelidae 12/123 | Blastobasidae 24/377 |
| Adelidae 5/294 | Oecophoridae 313/3 308 |
| Incurvariidae 11/51 | Schistonoeidae 1/1 |
| Cecidosidae 5/16 | Lypusidae 3/21 |
| Prodoxidae 9/98 | Chimabachidae 2/6 |
| Tischeriidae 3/110 | Peleopodidae 7/28 |
| Innominada 25/100 | Elachistidae 161/3 201 |
| Millieriidae 3/4 | Syringopaidae 1/1 |