



La riqueza de especies de líquenes de México no se conoce con certeza. Se estima, por ejemplo, que sólo en la porción tropical del país puede haber 3 600 especies que es la mitad de las calculadas para todo el neotrópico. Si se toma en cuenta que México es también diverso en climas y tipos de vegetación no tropicales, el número de especies de líquenes quizás alcance las 5 000. De ser así, el país ocuparía el segundo lugar en este rubro entre los países neotropicales, después de Brasil (4 900 especies, sólo en sus partes tropicales) y representaría del 18-37% de las especies de líquenes a nivel mundial, que es estimado entre 13 500 y 20 000. Pero el conocimiento de la flora liquenológica de México está todavía lejos de completarse. Se estima que a la fecha hay registro de alrededor del 50% de las especies de líquenes del país. El objetivo es presentar un conteo actualizado de las especies de líquenes conocidas para México. En este trabajo se contabilizaron 2 722 especies y 111 categorías infraespecíficas. Están agrupadas en 2 divisiones, 2 subdivisiones, 6 clases, 21 órdenes, 84 familias y 364 géneros. El 90% de las especies son clorolíquenes y el resto cianolíquenes. La mayoría de ellas tienen forma de crecimiento costrosa (62%), les siguen las foliosas (28%) y las fruticosas (11%). El sustrato más común donde crecen es la corteza de los árboles (46%), seguido por las rocas (34%), hojas (11%) y suelo (10%). El tipo de vegetación con más riqueza de líquenes es el bosque templado (947especies), seguido por el matorral xerófilo (544), el bosque tropical húmedo (391), el bosque estacionalmente seco (386) y el bosque húmedo de montaña (230). Los estados con más especies de líquenes son Baja California (710), Veracruz (700), Sonora (516), Chihuahua (515) y Baja California Sur (511), y aquellos con menos especies son Campeche (17), Aguascalientes (2) y Tabasco (1), pero esto refleja más la disparidad de los estudios liquenológicos entre estados que sus diferencias reales en diversidad de líquenes.
Mexican lichen species richness is not known with certainty. It is estimated, for example, that 3 600 species could occur only on its tropical parts, which is half of the total estimated for the Neotropics. Considering Mexico’s climate and non-tropical vegetation diversity, the number of lichen species, as some authors estimate, may reach 5 000. Therefore, the country would be only second to Brazil (4 900 lichen species in its tropical parts) among the Neotropical countries and it could harbor between 18-37% of the 13 500 to 20 000 estimated species worldwide. We consider that only near 50% of the Mexican lichen species have been recorded. The goal is to present an updated account of the known lichen species for Mexico. Herein, 2 722 species and 111 infraspecific categories grouped in 2 divisions, 2 subdivisions, 6 classes, 21 orders, 84 families, and 64 genera are recorded. Ninety percent of the species are chlorolichens and the remaining 10% cyanolichens. Crustose lichens are the most frequent growth form (62%), followed by foliose (28%), and fruticose forms (11%). The most common substrate is bark (46%), followed by rocks (34%), leaves (11%) and soil (10%). Temperate forest is the species richest type of vegetation (947 species), followed by xerophytic scrubland (544), tropical humid forest (391), seasonal dry forest (386) and humid mountain forest (230). The states with higher number of lichen species are Baja California (710), Veracruz (700), Sonora (518), Chihuahua (515) and Baja California Sur (511), and those with less species are Campeche (17), Aguascalientes (2) and Tabasco (1). However, these figures reflect more the disparity in the number of studies conducted in each state than real differences in lichen diversity among states.
Los líquenes son formas de vida simbiótica estables formadas por organismos de 2 o 3 diferentes reinos biológicos. Siempre están constituidos por un hongo (micobionte) y 1 o 2 autótrofos algales y/o cianobacteriales (fotobiontes) en una unidad morfofisiológica distinta a sus componentes de vida libre. Entre los componentes del liquen, y entre éste y el ambiente, se mantiene un flujo de agua, carbono, nitrógeno y otros elementos esenciales (Lawrey, 1984). Este tipo de simbiosis es considerada como mutualista; es decir, con beneficio mutuo para los 2 o 3 componentes (Scott, 1973; Hale, 1983; Honegger, 1998, 2008; Nash, 2008a), pero ocasionalmente ha sido calificada como un parasitismo controlado por parte del micobionte, el cual se beneficia de los carbohidratos producidos por el fotobionte, provocando que éste crezca más lentamente que en vida libre (Ahmadjian, 1993). Los líquenes también se han conceptualizado como ecosistemas en miniatura con un productor primario (el fotobionte), un consumidor primario (el micobionte) y, en ciertos casos, con descomponedores, como cuando un liquen es parasitado por otro hongo (Tehler, 1996; Nash, 2008a).
Los líquenes producen sustancias orgánicas a partir del metabolismo primario y del metabolismo secundario. Los primeros son metabolitos intracelulares encontrados también en hongos no liquenizados, algas y plantas vasculares como aminoácidos, proteínas, polioles, polisacáridos, carotenoides y vitaminas (Hawksworth y Hill, 1984; Elix y Stocker-Wörgöter, 2008). Los segundos representan el mayor número de compuestos orgánicos liquénicos. Son producidos por el micobionte y depositados en el exterior de las paredes hifales. Se conocen más de 1 000, de los cuales sólo una mínima cantidad (~ 50) ha sido identificada también en hongos no liquenizados o en plantas superiores. Incluyen ácidos alifáticos, para y meta-depsidas, depsidonas, bencil ésteres, dibenzofuranos, ácidos úsnicos, xantonas, antraquinonas, terpenoides y ácido pulvínico y sus derivados (Hawksworth y Hill, 1984; Elix y Stocker-Wörgöter, 2008; Huneck y Yoshimura, 1996). La importancia farmecéutica de estas sustancias radica en sus propiedades antibióticas, antitumorales y/o anti-VIH (Richardson, 1988; Hirabayashi et al., 1989; Pereira, 1998).
La utilidad de los metabolitos secundarios en la taxonomía de los líquenes está bien establecida (Culberson, 1969, 1970; Hawksworth y Hill, 1984; Hafellner, 1988; Elix, 1996). Su estudio ha permitido, en conjunción con la morfología, anatomía, los análisis de la distribución geográfica y filogenéticos, la delimitación e interpretación de relaciones a nivel de especie, género, suborden y familia como en Letharia, Parmeliaceae y Pertusariaceae (Culberson, 1969, 1970, 1986; Hawksworth y Hill, 1984; Hafellner, 1988; Elix, 1996; Schmitt y Lumbsch, 2004).
La liquenización es una estrategia de vida muy antigua y exitosa, como lo demuestran su existencia desde por lo menos el Devónico Temprano (≈ 400 millones de años) y su diversidad actual estimada entre 13 500 y 20 000 especies a nivel mundial, que podría alcanzar 30 000 al estudiarse regiones inexploradas, principalmente del Neotrópico (Tehler, 1983; Ahmadjian y Paracer, 1986; Ahmadjian, 1987; Poelt, 1994; Yuan et al., 2005; Feuerer y Hawksworth, 2007; Lücking et al., 2009; Lumbsch et al., 2011). El nombre de las especies de los líquenes se asigna al hongo y, por lo tanto, son clasificados dentro del Reino Fungi.
La diversidad de los líquenes en México es un tema todavía abierto. El avance en esta materia ha sido muy lento y esporádico desde tiempos coloniales hasta fechas recientes, dependiendo más del interés de exploradores, colectores e investigadores de instituciones extranjeras que de los mexicanos. La cronología del desarrollo de la liquenología en México desde la colonia hasta la penúltima década del siglo pasado está relatada en Godínez y Ortega (1989). El primer conteo de las especies conocidas de líquenes de México fue publicado por Imshaug (1956), quien dio la cifra de 1 000 especies. Un segundo recuento lo hicieron Ryan et al. (1996), quienes detectaron varios nombres no válidos en Imshaug (1956) y contabilizaron más de1 300 especies conocidas para México.
En este capítulo se presenta un tercer conteo actualizado y desglosado de las especies y categorías supraespecíficas de los hongos liquenizados de México. Se partió del catálogo de Ryan et al. (1996) añadiendo datos de la literatura aparecida entre 1996 y 2012. Adicionalmente se consultaron las bases de datos de la colección de líquenes de MEXU, del Consorcio de Herbarios de Líquenes de Norteamérica (CNALH, por sus siglas en inglés, http//:lichenportal.org/portal/index.php), las listas del herbario de líquenes de la Universidad Estatal de Arizona (http://nhc.asu.edu/lherbarium) y la información de proyectos en curso del Laboratorio de Líquenes del Instituto de Biología, UNAM. La clasificación taxonómica de los nombres recopilados fue revisada en línea durante el año 2012 en Mycobank (http://www.mycobank.org) y el Index Fungorum (http://www.indexfungorum.org). Es preciso señalar que estas bases de datos no tienen actualización inmediata, por lo que pueden no reflejar los avances más recientes, por ejemplo la sinonimización de Thelotremathaceae y Graphidaceae (Mangold et al., 2008), o la inclusión en esta última de Gomphillaceae (Rivas Plata et al., 2012).
Descripción morfológica del grupoEn los líquenes hay una gran variedad de estructuras, tamaños y colores que evidencian el papel de esta simbiosis en la generación de novedades evolutivas, apreciables particularmente en la radiación morfológica de los ascomicetos (Büdel y Scheidegger, 2008). La diversidad morfológica de los líquenes se explica parcialmente por adaptación a los muy variados microambientes donde habitan (Hawksworth, 1988b). Los líquenes varían considerablemente en tamaño desde apenas milímetros de diámetro hasta varios metros de longitud y colorean llamativamente los sustratos donde crecen. Los colores de los líquenes pueden ser verdes de tonos muy variados, blanquecinos, grises, cafés, negros y una gran gama de brillantes colores amarillos, anaranjados o rojos, debidos ya sea a los pigmentos fotosintéticos del fotobionte o a las propiedades ópticas de los metabolitos secundarios producidos por el micobionte.
A los líquenes con fotobionte algal se les conoce como clorolíquenes y a los que tienen fotobionte cianobacteriano como cianolíquenes. El grado de liquenización o integración de los hongos con su fotobionte se refiere al tipo de contacto entre ellos (Nash, 2008a). Este contacto se da a través de las paredes celulares con o sin distintos tipos de haustorios o apresorios y una capa fúngica hidrofóbica que se extiende sobre la pared celular del fotobionte (Honegger, 2008). El fotobionte estimula al hongo para formar un talo diferenciado, pero la apariencia de la mayoría de los líquenes está determinada por los tejidos fúngicos (Jahns, 1988; Büdel y Scheidegger, 2008). Algunos autores consideran al micobionte como el “habitante externo” (exhabitant) y el fotobionte el “habitante interno” (inhabitant) de esta simbiosis (Hawksworth, 1988b).
En los líquenes más sencillos, las hifas se entrelazan laxamente envolviendo a las células del fotobionte (Caliciales), o forman un talo algal gelatinoso surcado por hifas o un talo con apariencia capilar como en Coenogonium, Cystocoleus y Ephebe (Jahns, 1988; Hawksworth y Hill, 1984; Büdel y Scheidegger, 2008). Los talos más complejos son estratificados, con el fotobionte localizado en una capa interna por debajo de una cubierta fúngica, estos talos son más frecuentes en los macrolíquenes.
Los talos liquénicos anatómicamente se subdividen en homómeros o heterómeros, de acuerdo con su organización y su diferenciación “tisular”. Los primeros son homogéneos, sin estratificación alguna y con los fotobiontes dispersos al azar en el tapete fúngico. En los segundos hay capas fúngicas definidas: una corteza superior, una médula -que incluye a la capa algal- y una corteza inferior también fúngica. Los pseudotejidos o plencténquimas de estas capas son de 2 tipos generales: a) pseudoparénquima (paraplecténquima), caracterizado por hifas más o menos isodiamétricas con paredes soldadas, indistinguibles individualmente e irregularmente orientadas, y b) prosénquima (prosoplecténquima) cuyas hifas son alargadas, distinguibles entre sí, con paredes gruesas y orientadas paralela y/o perpendicularmente al eje del talo.
Por la forma de crecimiento del talo de los líquenes, se distinguen 3 tipos morfológicos generales (Fig. 1): costrosos (microlíquenes), foliosos y fruticosos (ambos macrolíquenes) aunque hay también algunas formas con características intermedias o que combinan 2 de los tipos generales, por ejemplo, una porción costrosa o foliosa y otra fruticosa. Las formas de crecimiento no tienen valor taxonómico. Por ejemplo, algunos taxa incluyen tanto formas costrosas como foliosas y/o fruticosas, como el género Caloplaca y las familias Roccellaceae y Teloschistaceae.
 Hoffm., foto J. Aguilar. C, fruticoso, Teloschistes exilis (Michx.) Vain., foto J. Aguilar. D, fruticoso compuesto, Cladonia macilenta Hoffm., foto J. Aguilar.")
Formas de crecimiento de los talos liquénicos. A, costroso, Arthonia redingeri Grube, foto R. Miranda. B, folioso, Lobaria pulmonaria (L.) Hoffm., foto J. Aguilar. C, fruticoso, Teloschistes exilis (Michx.) Vain., foto J. Aguilar. D, fruticoso compuesto, Cladonia macilenta Hoffm., foto J. Aguilar.
Los microlíquenes forman talos con apariencia de costras más o menos circulares y adheridas totalmente al sustrato. Pueden ser homómeros o heterómeros y miden de unos milímetros a unos cuantos centímetros cuadrados de superficie. Algunas especies costrosas desarrollan un margen (protalo) de color variable, como Rhizocarpon y Cryptothecia.
Existen diversas variantes de líquenes costrosos: pulverulentos o leprosos (Lepraria), escuamulosos (Psora, Catapyrenium, Toninia), peltados (Peltula), pulvinados (Mobergia), lobulados (Caloplaca, Lecanora), efigurados (Dimelaena, Acarospora, Pleopsidium) y subfruticosos (algunas especies de Caloplaca) (Büdel y Scheidegger, 2008). Algunos líquenes costrosos crecen dentro de las rocas (endolíticos, Verrucaria), o son endofleódicos (Graphis), cuya corteza superior presenta fragmentos del sustrato donde crecen, como cristales de roca (litocorteza) o células epidérmicas de hojas (filocorteza, Strigula) o de corteza (fleocorteza).
Los líquenes foliosos se caracterizan por tener talos laminares adheridos total o parcialmente al sustrato mediante rizinas u ombligos. Miden desde unos cuantos centímetros de diámetro hasta 30cm. La mayoría de estos talos son de clorolíquenes y son heterómeros, con excepción de los cianolíquenes, que son homómeros y de consistencia gelatinosa (Collema, Leptogium). Se distinguen 2 subtipos de talos foliosos: a) los laciniados o lobulados, cuyos lóbulos pueden ser aplanados y sólidos o cilíndricos y huecos como en Hypogymnia y Menegazia; con patrones de ramificación variable y superficie inferior con rizinas, cilios o tomento (Parmelia, Heterodermia, Parmotrema, Peltigera, Sticta, Lobaria, etc.) y b) los umbilicados, que son circulares o multilobulados con ramificación escasa y que se reconocen porque se fijan al sustrato en un punto central que por la cara superior del talo tiene un aspecto de ombligo (Dermatocarpon, Umbilicaria). Hay un tipo peculiar de líquenes foliosos conocidos como vagrantes, que nunca se fijan al sustrato y son desplazados por el viento (Xanthoparmelia convoluta, X. vagrans). Estos líquenes son más comunes en zonas áridas y semiáridas, son higroscópicos y su fuente de humedad es el rocío. En estado seco se enrollan exponiendo la superficie inferior del talo, y al hidratarse se extienden exponiendo la superficie superior (Büdel y Sheidegger, 2008).
Los talos fruticosos tienen apariencia arbustiva o filamentosa, pueden ser erectos o péndulos, su tamaño oscila desde unos cuantos centímetros a varios metros de longitud (Usnea longissima). Sus patrones de ramificación son variables y las ramas pueden ser aplanadas (Evernia) o cilíndricas (Usnea). Todos estos talos son siempre heterómeros, con tejidos de soporte externos o internos (Cladonia, Alectoria, Usnea). Algunos líquenes fruticosos tienen talos compuestos o bifásicos; es decir, una porción horizontal vegetativa foliosa o escuamulosa y una porción vertical reproductiva, los llamados podecios (Cladonia) o pseudopodecios (Stereocaulon) que portan las estructuras de reproducción sexual.
Ciclo de vidaLos hongos liquenizados tienen una fase sexual (teleomorfo) y una asexual (anamorfo). Las divisiones taxonómicas de los líquenes están definidas por sus estructuras de reproducción sexual: Ascomycota o Basidiomycota. En la primera división se agrupan los líquenes que producen ascas y ascosporas en 2 tipos de cuerpos fructíferos: peritecios y apotecios. Los peritecios son fructificaciones cerradas, globosas o en forma de frasco, sin el himenio expuesto y con un tubo que desemboca en el ostiolo, por donde liberan las ascosporas. Los apotecios, en cambio, son estructuras abiertas con forma de disco o copa, tienen el himenio expuesto y distintos mecanismos de dehiscencia de las ascas para liberar las ascosporas. Por su parte, los basidiomicetos liquenizados forman basidiomas que son típicos de los Aphyllophorales y los Agaricales.
Las estructuras asexuales de los líquenes comprenden distintos conidiomas que forman conidiosporas o esporas asexuales y una enorme variedad de propágulos vegetativos. Los conidiomas más comunes de los hongos liquenizados son picnidios, campilidios e hifóforos, estos 2 últimos característicos de los líquenes foliícolas. Los propágulos vegetativos liquénicos son muy diversos, incluyen fragmentos de talo, isidios, soredios, filidios, esquizidios, entre otros. El grado de diferenciación de estos propágulos es muy variable, pueden consistir sólo de células del fotobionte envueltas por el micobionte (soredios) o presentar las distintas capas del talo, como en el caso de los isidios, esquizidios y filidios.
A diferencia de la reproducción sexual del hongo mediante esporas, que requiere de la re-liquenización o encuentro en el ambiente del componente fúngico con su fotobionte compatible, la reproducción por propágulos vegetativos compuestos por ambos simbiontes permite la dispersión y el establecimiento de los talos sin necesidad de un proceso de re-liquenización.
HábitatsEn condiciones naturales, los hongos liquenizados sólo se encuentran como biotrofos simbióticos ecológicamente obligados (Honegger, 2008). Los líquenes crecen en sustratos naturales o sobre materiales antropogénicos. Sus sustratos naturales más comunes son: el suelo (edafícolas), las rocas (saxícolas), las cortezas vegetales (corticícolas, epi o endofleódicos) y las hojas (foliícolas), pero también pueden crecer sobre animales. La gama de sustratos artificiales donde crecen es muy amplia, se les observa creciendo sobre vidrio, lámina, plástico y asbesto, entre muchos otros materiales.
Los líquenes pueden ser considerados como extremistas ambientales por su presencia en ambientes que son adversos para muchas otras formas de vida, como los desiertos más secos y cálidos o los fríos. No obstante, algunas especies de líquenes son muy sensibles a presiones ambientales como la radiación solar ultravioleta, la sequía, el calor, el congelamiento y los cambios abruptos producidos por la deforestación (Hawksworth y Hill, 1984; Kappen, 1994; Lücking, 1998, 2001). La sensibilidad de muchas especies de líquenes a contaminantes ambientales como el dióxido de azufre, metales y diversos compuestos nitrogenados, entre otros, ha sido ampliamente documentada (Nash, 2008d). Más recientemente se ha estudiado la respuesta de los líquenes al calentamiento global (Herk et al., 2002; Hollister et al., 2005; Sancho et al., 2007; Cezanne et al., 2008).
Los líquenes son componentes indispensables en los ecosistemas templados y tropicales. Contribuyen a los procesos ecosistémicos como productores primarios, descomponedores, formadores de suelo y en los ciclos del agua y los nutrientes (Jones, 1988; Lange et al., 2000; Nadkarni, 2000; Nash, 2008a, b, c; Seaward, 2008). Los nutrientes esenciales para los seres vivos, como el N, K, P y S, son obtenidos por los líquenes directamente del aire y los incorporan al ecosistema. Se ha estimado que algunos ecosistemas forestales consiguen vía los líquenes hasta 25% del N (Fritz-Sheridan, 1988; Knops y Nash, 1996; Seaward, 2008). Los líquenes son también importantes captadores de agua desde fuentes atmosféricas, la cual pueden absorber en cantidades de hasta 3 000% su peso seco. Estos organismos forman parte sustancial o complementaria de la dieta de diversos herbívoros vertebrados e invertebrados y constituyen un hábitat para microorganismos y artrópodos (Seaward, 1988; Longton, 1992). Algunas aves los utilizan como material de construcción para sus nidos.
ClasificaciónLos hongos liquenizados son clasificados en el reino Fungi en las divisiones Ascomycota y Basidiomycota. Representan más del 20% de las especies conocidas de hongos. Su diversidad mundial se estima entre 13 500 hasta 20 000 especies; pero cálculos predictivos más recientes establecen que se pueden alcanzar hasta 28 000 especies (Lücking et al., 2009). La gran mayoría de los líquenes son ascomicetos y representan el 46% del total de especies de este grupo de hongos, mientras que los basidiolíquenes constituyen sólo el 0.3% de las especies conocidas de basidiomicetos (Hawksworth, 1988a, 2001; Hawksworth et al., 1995; Sipman y Aptroot, 2001; Feuerer y Hawksworth, 2007). Los fotobiontes de los líquenes son mucho menos diversos, pues sólo se conocen alrededor de 100 especies de 50 géneros, siendo los más comunes Trebouxia y Trentepohlia, entre los clorolíquenes, y Nostoc y Scytonema en los cianolíquenes (Lücking et al., 2009). Los nombres de los líquenes son asignados al micobionte. Los fotobiontes se nombran y clasifican por separado.
La clasificación y el estudio de la evolución de los líquenes es abordado con las mismas técnicas de análisis cladístico y molecular que los hongos no liquenizados, cuyo avance progresivo ha generado cambios continuos en el sistema de clasificación y en la forma de entender las relaciones filogenéticas de los hongos (Tehler, 1988; Eriksson et al., 2002; Lutzoni et al., 2004; James et al., 2006; Miądlikowska et al., 2006; Hibbett et al., 2007).
La alta diversidad resultante y la presencia de hongos liquenizados y no liquenizados en grupos como los Ostropales llevaron a considerar a los primeros como polifiléticos (Gargas et al., 1995; Lutzoni y Vilgalys, 1995a, b; Tehler, 1996). Sin embargo, hallazgos recientes sugieren que la mayoría de los líquenes forman un grupo monofilético del cual se derivaron líneas no liquenizadas como los Eurotiomycetes (que incluye los géneros Aspergillus y Penicillium) y los Chaetothyriomycetes (Lutzoni et al., 2001; James et al., 2006; Miądlikowska et al., 2006).
Estudios de líquenes en MéxicoHasta el año 1930, el conocimiento de los líquenes de México se debió casi exclusivamente a recolectas e identificaciones de estudiosos europeos, con la participación creciente de los norteamericanos desde la segunda mitad del siglo XIX. Esto se aprecia en la literatura referida por Imshaug (1956), quien hizo el primer catálogo de los líquenes mexicanos. El desarrollo de la florística de los líquenes de México desde tiempos de la colonia hasta la década de 1980 se encuentra descrito en el trabajo de Godínez y Ortega (1989).
En un resumen breve, el inicio del conocimiento formal de los líquenes de nuestro país se puede fechar en la segunda década del siglo XIX, cuando el botánico alemán Carl Segismund Kunth identificó los ejemplares mexicanos recolectados por los exploradores Alexander von Humboldt y Aimé Bonpland en el período 1803-1806 (Godínez y Ortega, 1989). Durante todo el siglo XIX, los principales promotores del conocimiento de los líquenes mexicanos siguieron siendo europeos, ya sea como colectores o como taxónomos. Entre ellos se puede citar a liquenólogos famosos como Nylander, Krempelhuber, Vainio, Liebman y Müller Argoviensis. En la segunda mitad de ese siglo, empiezan a figurar autores norteamericanos como Schott, Smith, Brunner y Pringle (Godínez y Ortega, 1989).
La recolecta sistemática de líquenes mexicanos, no obstante, comenzó durante las 3 primeras décadas del siglo XX. Los primeros estudios en México con características de floras locales o regionales son los trabajos de Bouly de Lesdain (1914, 1929, 1933). En la década de 1930 se hizo el primer estudio de los líquenes del valle de México como trabajo de tesis de maestría (Gibert, 1935) y los del valle Actopan y valle del Mezquital en Hidalgo por Ruiz-Oronoz (1936, 1937). Para la siguiente década se publicó un trabajo de Ruiz-Oronoz y Herrera (1948) que incluyó líquenes, entre otros organismos, de Cuicatlán, Oaxaca.
Hacia la segunda mitad del siglo XX y hasta la actualidad, la participación extranjera en el conocimiento de la flora liquenológica de México no ha decaído. Entre los trabajos más conocidos sobre este tema se encuentran los de Hale (1965, 1967, 1977), Wirth y Hale (1963), Hale y Kurokawa (1964) y Hale y Wirth (1971). El trabajo hecho en este campo por mexicanos recibió un considerable impulso a partir de 1970 con los aportes de Gastón Guzmán y colaboradores (Brizuela y Guzmán, 1971; Dávalos de Guzmán et al., 1972; Guzmán y González de la Rosa, 1976; González de la Rosa y Guzmán, 1976; Welden et al., 1979; Coutiño y Mojica, 1982, 1985; Guzmán-Dávalos y Álvarez, 1987; Álvarez y Guzmán-Dávalos, 1988; Patrón et al., 1985).
No obstante, el primer tratamiento profundo y moderno de los líquenes mexicanos es indiscutiblemente el de la flora liquénica del Desierto Sonorense, iniciado por Thomas H. Nash III, en la década de 1990, que abarcó un área de 600 000km2 de México y los Estados Unidos. Este proyecto involucró aproximadamente a 100 liquenólogos de 23 países (Bates y Bungartz, 2012), produjo cerca de 200 publicaciones que dan cuenta de más de 1 900 especies y 341 géneros y una obra de 3 volúmenes “Lichen Flora of the Great Sonoran Desert Region” (Nash et al., 2002, 2004, 2007). Estas especies representan alrededor del 40% de las reportadas en la lista de líquenes de Norteamérica (Esslinger, 2012). Por ello la región noroeste de México no sólo es la más estudiada liquenológicamente del país sino una de las mejor conocidas del mundo en este aspecto. El proyecto mencionado ha sido sin duda la más grande contribución al conocimiento de la diversidad de los líquenes mexicanos y un modelo de colaboración científica multinacional.
Desde mediados de la década de 1990 se han realizado distintos estudios sobre biodiversidad y ecología de líquenes de México que han ampliado el conocimiento de la riqueza de estos organismos. Entre ellos se encuentran los trabajos con líquenes de hábito foliícola de Veracruz, Puebla y Oaxaca (Herrera-Campos y Lücking, 2002, 2003, 2004; Herrera-Campos et al., 2004a, b, 2005; Barcenas-Peña, 2004, 2007), los macrolíquenes corticícolas de Oaxaca (Pérez-Pérez y Herrera-Campos, 2004), los líquenes de la Reserva Ecológica del Pedregal de San Ángel (Herrera-Campos y Lücking, 2009), los microlíquenes de Querétaro y Jalisco (Miranda, 2008, 2012), del género Usnea (Clerc y Herrera-Campos, 1997; Herrera-Campos, 1998; Herrera-Campos et al., 1998, 2001) y el primer estudio de delimitación de especies de este género combinando caracteres de la taxonomía clásica y moleculares (Ament, 2012).
Además de los trabajos florístico-taxonómicos, se han realizado algunos estudios ecológicos con diferentes enfoques. Por ejemplo, se ha investigado la sucesión de insectos en comunidades de líquenes y briofitas (Mojica y Johansen, 1990) y la ecoquímica de líquenes sobre rocas mineras (Rodríguez y Rosales, 1995). Los líquenes son ampliamente utilizados en muchos países del mundo como bioindicadores de la calidad del aire, como indicadores de continuidad ecológica o estado de conservación de bosques y para identificar sitios críticos de biodiversidad. Sin embargo, en México y otros países tropicales estos enfoques se han visto limitados por los faltantes de conocimiento taxonómico y ecológico sobre estos organismos en la región. Los únicos trabajos mexicanos con estos enfoques son los de Zambrano et al. (2000, 2002), quienes utilizaron líquenes como indicadores de la calidad del aire en la Cuenca de México, y los de Pérez-Pérez (2005) y Pérez-Pérez et al. (2011) sobre el impacto del manejo forestal en la riqueza de los macrolíquenes de Oaxaca.
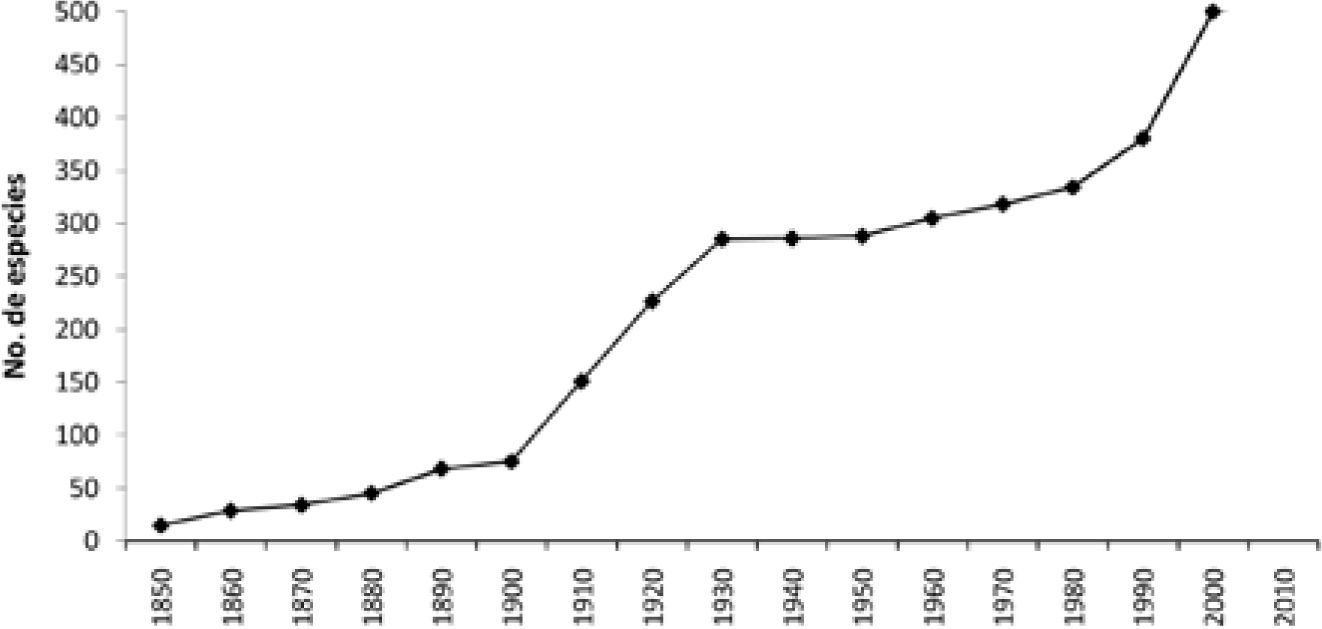
En el presente trabajo se contabilizó un total de 506 especies de líquenes descritas para México entre 1850 y 2012 (Fig. 2). De la segunda mitad del siglo XIX, se encontró registro documental de 69 especies, siendo sus autores principales W. Nylander y J. Müller Argoviensis, con 35 y 16 especies, respectivamente. En el siglo XX se alcanzó un total de 310 especies. La figura 2 muestra que hubo 2 períodos importantes de adición de especies: uno entre los años 1910-1930 y otro en los años noventa. Durante los 30 años del primer período destaca la contribución de B. de Lesdain, quien describió aproximadamente el 42% de las especies. El pico de registros en la década de 1990 se explica por el proyecto de la liquenobiota del Desierto Sonorense. En este lapso, se describieron 46 especies con autorías de 34 liquenólogos, entre los que aparecen los primeros autores mexicanos de especies de líquenes. En la primera década del presente siglo el número de especies descritas para el país alcanzó su máximo histórico, 118. Sus autores suman alrededor de 47, entre los que destacan F. Bungartz, O. Breuss, E. Timdal, T. Esslinger, M. A. Herrera-Campos y R. Lücking. Con este último autor se realizó la descripción del mayor número de especies mexicanas de líquenes, 23%, en particular de hábito foliícola; algunas de ellas en co-autoría por primera vez con las estudiantes mexicanas A. Barcenas-Peña y P. Martínez-Colín. También en este período se estableció en el Instituto de Biología de la UNAM el primer laboratorio mexicano dedicado a los líquenes. En lo que va de la segunda década de este siglo se han descrito 7 nuevas especies, de los autores mencionados y de R. E. Pérez-Pérez, R. Miranda, T. Esslinger y T. H. Nash.

La pendiente ascendente de la curva de acumulación de especies de líquenes descritas originalmente para México (Fig. 2) es un fuerte indicio de que la recolecta y la descripción de nuevas especies están lejos de haber concluido. Las últimas 3 especies mexicanas descritas aparecen en Lumbsch et al. (2011). Estos autores estiman en alrededor de 10 mil las especies liquénicas no descritas a nivel mundial, gran parte de las cuales se espera encontrarlas en áreas y hábitats poco explorados, principalmente en latitudes tropicales.
DiversidadEl número de especies de líquenes en México es todavía muy incierto por la carencia de trabajo taxonómico para la mayor parte del territorio. El catálogo de líquenes mexicanos de Imshaug (1956) contenía cerca de 1 000 nombres específicos, de los cuales sólo la mitad correspondía a nombres válidos aceptados (Ryan et al., 1996). Estos últimos autores actualizaron ese catálogo y añadieron la información publicada durante los siguientes 40 años, lo que resultó en más de 1 300 nombres específicos válidos. Subsecuentemente se reportaron 500 especies (Ahti, 2000; Ryan et al., 2000; Herrera-Campos et al., 2004a, b, 2005) para un total aproximado de 1 800 especies de líquenes en el país.
Considerando que las investigaciones liquenológicas en México siguen siendo escasas y que hay pocos grupos de líquenes monografiados, es probable que el número de especies conocidas subestime por mucho el número real. Por ejemplo, Lücking et al. (2009) calculan que nada más en la parte tropical del país hay 3 600 especies. Esto significa la mitad de las especies esperadas para todo el Neotrópico (7 200 especies), el mismo número de especies de líquenes que Colombia, y ligeramente debajo de las de Perú (3 700), Bolivia y Venezuela tienen 4 200 y Brasil 4 900 sólo en sus partes tropicales. Sin embargo, si se toma en cuenta que México se caracteriza por una alta variedad de tipos de vegetación y mezclas de climas tropicales y templados, se puede conservadoramente suponer la existencia de alrededor de 5 000 especies liquénicas, una cifra por debajo únicamente de Brasil, entre los países neotropicales. Si la estimación de 5 000 especies de líquenes en México es una buena aproximación, su flora liquénica podría representar entre el 18 y el 37% respecto al mundo y 70% en comparación con el Neotrópico.
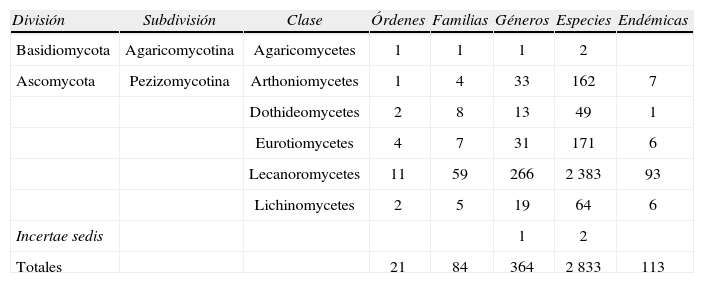
ResultadosA continuación se resumen los hallazgos de este estudio sobre la riqueza de especies conocida hasta ahora de los líquenes mexicanos. Esta recopilación arrojó un total de 2 833 taxa: 2 722 especies y 111 categorías infraespecíficas (18 subespecies, 68 variedades y 25 formas). Estas corresponden taxonómicamente a 2 divisiones, 2 subdivisiones, 6 clases, 21 órdenes, 84 familias y 364 géneros. (Cuadro 1, con un género incertae sedis en la categoría de subdivisión y otro en la de clase).
Resumen taxonómico de los líquenes mexicanos
| División | Subdivisión | Clase | Órdenes | Familias | Géneros | Especies | Endémicas |
| Basidiomycota | Agaricomycotina | Agaricomycetes | 1 | 1 | 1 | 2 | |
| Ascomycota | Pezizomycotina | Arthoniomycetes | 1 | 4 | 33 | 162 | 7 |
| Dothideomycetes | 2 | 8 | 13 | 49 | 1 | ||
| Eurotiomycetes | 4 | 7 | 31 | 171 | 6 | ||
| Lecanoromycetes | 11 | 59 | 266 | 2 383 | 93 | ||
| Lichinomycetes | 2 | 5 | 19 | 64 | 6 | ||
| Incertae sedis | 1 | 2 | |||||
| Totales | 21 | 84 | 364 | 2 833 | 113 |
La gran mayoría de las especies de líquenes conocidas para México son ascomicetos (2 829 vs. 2 basidiomicetos), lo cual es consistente con la partición mundial de los líquenes a nivel de división. Asimismo, la clase más diversa en especies y otras categorías supraespecíficas es la de los Lecanoromycetes. Esta clase agrupa alrededor del 96% de los hongos liquenizados y es la más diversa en el reino Fungi (Miądlikowska et al., 2006). Se caracteriza por el desarrollo ascohimenial de los apotecios y de los peritecios; la presencia de ascas amiloides o hemialoides, funcionalmente unitunicadas con distintas estructuras apicales y por la producción de una gran variedad de metabolitos secundarios (Lumbsch et al., 2004; Miądlikowska et al., 2006; Schmitt, 2011).
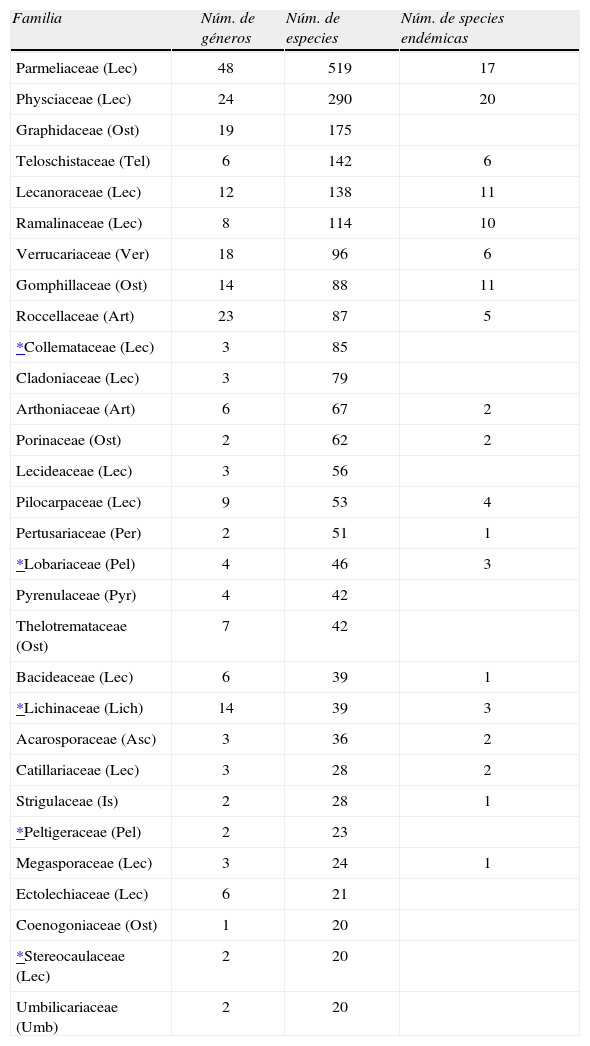
Del total de especies documentadas, 113, casi 4%, son endémicas para el país, las cuales incluyen, además de las 108 consideradas en el Cuadro 2, 5 especies endémicas más de las familias Gloeoheppiaceae (2), Haematommataceae (1), Peltulaceae (1) y Psoraceae (1).
Riqueza de géneros, especies y endemismos de las familias más diversas de líquenes mexicanos. Entre paréntesis, las iniciales de los órdenes a los que pertenecen
| Familia | Núm. de géneros | Núm. de especies | Núm. de species endémicas |
| Parmeliaceae (Lec) | 48 | 519 | 17 |
| Physciaceae (Lec) | 24 | 290 | 20 |
| Graphidaceae (Ost) | 19 | 175 | |
| Teloschistaceae (Tel) | 6 | 142 | 6 |
| Lecanoraceae (Lec) | 12 | 138 | 11 |
| Ramalinaceae (Lec) | 8 | 114 | 10 |
| Verrucariaceae (Ver) | 18 | 96 | 6 |
| Gomphillaceae (Ost) | 14 | 88 | 11 |
| Roccellaceae (Art) | 23 | 87 | 5 |
| *Collemataceae (Lec) | 3 | 85 | |
| Cladoniaceae (Lec) | 3 | 79 | |
| Arthoniaceae (Art) | 6 | 67 | 2 |
| Porinaceae (Ost) | 2 | 62 | 2 |
| Lecideaceae (Lec) | 3 | 56 | |
| Pilocarpaceae (Lec) | 9 | 53 | 4 |
| Pertusariaceae (Per) | 2 | 51 | 1 |
| *Lobariaceae (Pel) | 4 | 46 | 3 |
| Pyrenulaceae (Pyr) | 4 | 42 | |
| Thelotremataceae (Ost) | 7 | 42 | |
| Bacideaceae (Lec) | 6 | 39 | 1 |
| *Lichinaceae (Lich) | 14 | 39 | 3 |
| Acarosporaceae (Asc) | 3 | 36 | 2 |
| Catillariaceae (Lec) | 3 | 28 | 2 |
| Strigulaceae (Is) | 2 | 28 | 1 |
| *Peltigeraceae (Pel) | 2 | 23 | |
| Megasporaceae (Lec) | 3 | 24 | 1 |
| Ectolechiaceae (Lec) | 6 | 21 | |
| Coenogoniaceae (Ost) | 1 | 20 | |
| *Stereocaulaceae (Lec) | 2 | 20 | |
| Umbilicariaceae (Umb) | 2 | 20 |
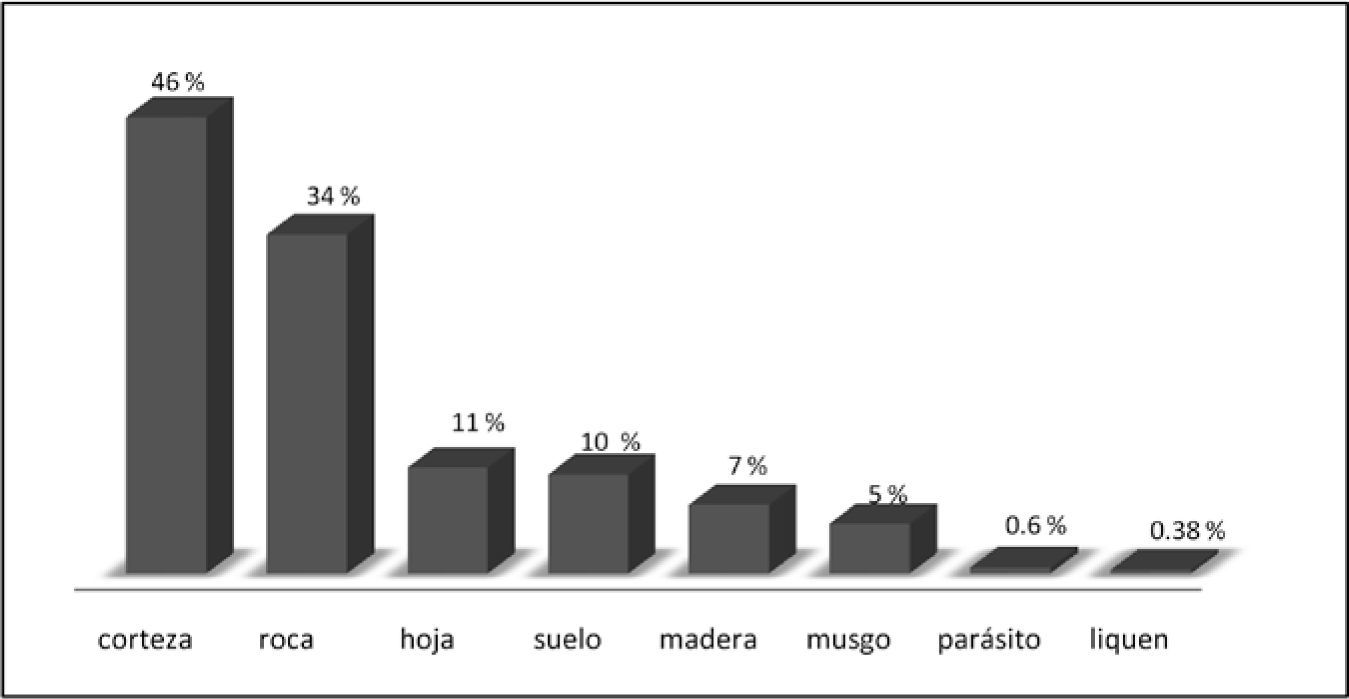
La mayoría de las especies conocidas (2 554, 90%) son clorolíquenes y el 10% restante (279) son cianolíquenes. La forma de crecimiento mejor representada en el país (1 716 especies, 60% del total) es la de los líquenes costrosos. Le siguen las formas foliosas (805, 28%) y las fruticosas con 228 especies (8%). Las formas compuestas están representadas por 101 especies. Algunas especies pueden presentar formas de crecimiento intermedias entre las anteriores. El tipo de sustrato con más especies de líquenes es la corteza vegetal, seguido por rocas, hojas y suelo. El 12% de las especies crece en madera o sobre musgo (sin ser foliícolas). Se tienen registros muy escasos en el país de especies de líquenes parásitos o liquenícolas (Fig. 3).

El orden más grande y diverso es Lecanorales con 1 573 especies, que representan 55.5% del total registrado hasta ahora. Le siguen en diversidad de especies Ostropales (14%), Arthoniales (5.7%), Teloschistales (5.1%), Verrucariales (3.3%), Peltigerales (2.9%), Pertusariales (2.8%), Pyrenulales (2.4%), Lichinales (2.1%), Acarosporales (1.2%) y Umbilicariales (0.7%).
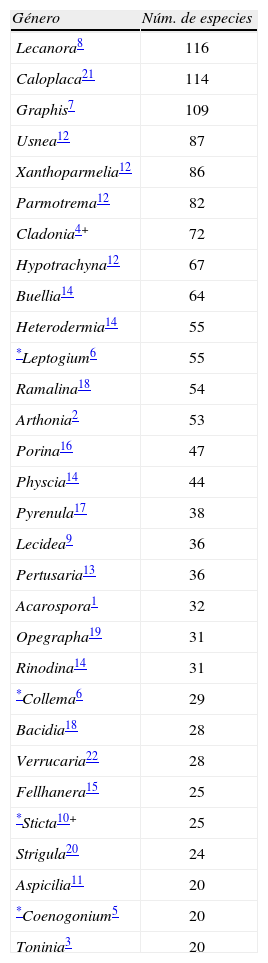
Las familias con más de 20 especies concentran el 60% de las especies totales (Cuadro 2). Las familias más ricas de los clorolíquenes son: Parmeliaceae (18.3%), Physciaceae (10.2%), Graphidaceae (6%), Teloschistaceae (5%) Lecanoraceae (4.8%), Ramalinaceae (4.0%) y con 3% Verrucariaceae, Gomphilaceae y Roccellaceae. Entre los cianolíquenes: Collemataceae (3%), Lobariaceae (1.6%) y Lichinaceae (1. 3 %). En el Cuadro 3 se muestran los géneros con más de 20 especies en México y sus familias. Lecanora, Caloplaca y Graphis rebasan las 100 especies. Les siguen con más de 80 especies las Parmeliaceae Usnea, Xanthoparmelia y Parmotrema. Entre las Cladoniaceae, el género mejor representado es Cladonia. Dentro de la familia Physciaceae son Buellia, Physcia y Rinodina. Los cianolíquenes con más especies son Collema y Sticta, Collemataceaea y Lobariaceae, respectivamente.
Géneros más diversos de líquenes mexicanos
| Género | Núm. de especies |
| Lecanora8 | 116 |
| Caloplaca21 | 114 |
| Graphis7 | 109 |
| Usnea12 | 87 |
| Xanthoparmelia12 | 86 |
| Parmotrema12 | 82 |
| Cladonia4+ | 72 |
| Hypotrachyna12 | 67 |
| Buellia14 | 64 |
| Heterodermia14 | 55 |
| *Leptogium6 | 55 |
| Ramalina18 | 54 |
| Arthonia2 | 53 |
| Porina16 | 47 |
| Physcia14 | 44 |
| Pyrenula17 | 38 |
| Lecidea9 | 36 |
| Pertusaria13 | 36 |
| Acarospora1 | 32 |
| Opegrapha19 | 31 |
| Rinodina14 | 31 |
| *Collema6 | 29 |
| Bacidia18 | 28 |
| Verrucaria22 | 28 |
| Fellhanera15 | 25 |
| *Sticta10+ | 25 |
| Strigula20 | 24 |
| Aspicilia11 | 20 |
| *Coenogonium5 | 20 |
| Toninia3 | 20 |
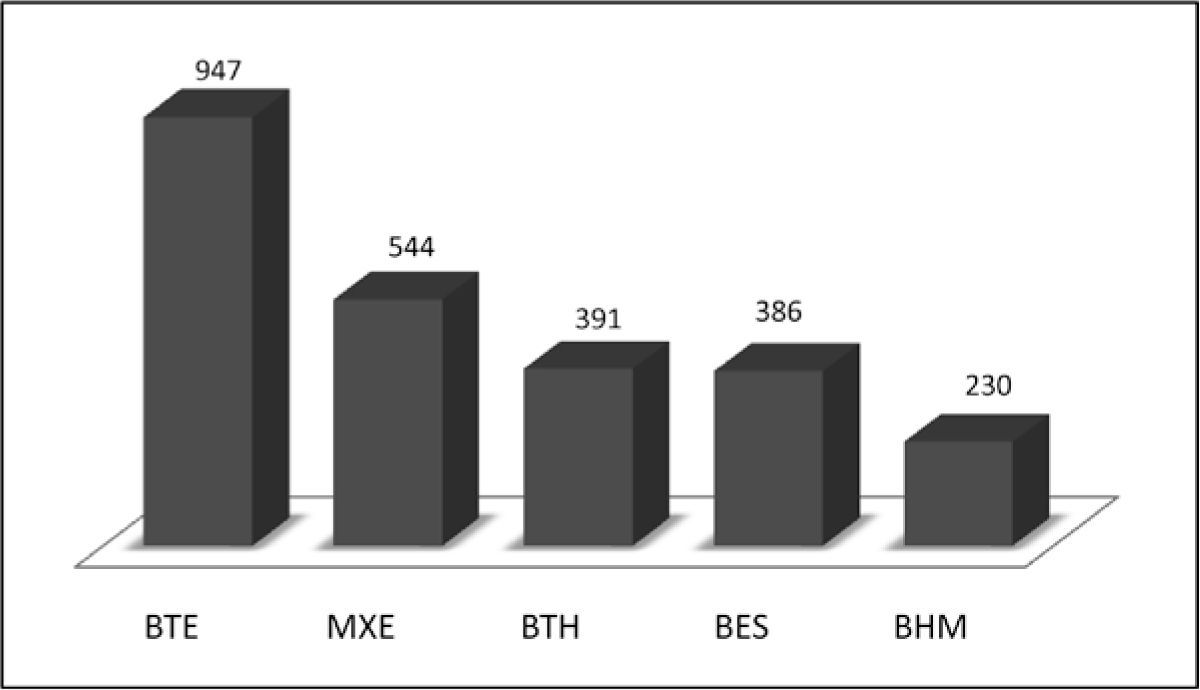
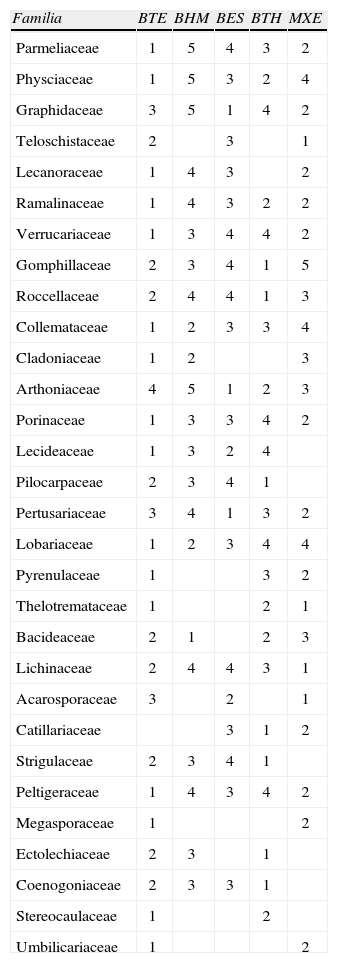
En este trabajo se tomó como base la clasificación y agrupación de los tipos de vegetación de México que hace Villaseñor (2013), la cual sintetiza la de Rzedowski (1978), a saber: 1) bosque estacionalmente seco (BES); 2) bosque húmedo de montaña (BHM); 3) bosque templado (BTE); 4) bosque tropical húmedo (BTH) y 5) matorral xerófilo (MXE). En la figura 4 se comparan los números de especies registradas por tipo de vegetación. La mayoría de especies (947) se ha recolectado en los bosques templados de coníferas y de encinos (BTE). Las familias más frecuentes son Parmeliaceae, Physciaceae, Lecanoraceae, Ramalinaceae, Colletamataceae, Cladoniaceae, Lobariaceae y Stereocaulaceae y microlíquenes de Verrucariaceae. En el matorral xerófilo (MXE) que incluye al chaparral, matorral desértico y zonas alpinas, se han reportado 544 especies, principalmente de las familias Teloschistaceae, Lichinaceae y Acarosporaceae.

La vegetación con más recolectas del bosque tropical húmedo (BTH) es el tropical perennifolio. De ahí se reportan 391especies de microlíquenes, principalmente foliícolas de las familias Pilocarpaceae, Gomphillaceae, Roccellaceae, Strigulaceae, Ectolechiaceae, Coenogoniaceae y Catillariaceae. En el bosque estacionalmente seco (BES) se han recolectado 386 especies sobre todo del bosque tropical caducifolio. Sobresalen las familias Graphidaceae, Arthoniaceae y Pertusariaceae. Finalmente, del bosque húmedo de montaña (BHM), que incluye a los bosques mesófilo de montaña, de neblina y Liquidambar, se han reportado 230 especies de líquenes, con algunas localidades próximas a bosque de coníferas o bosque tropical húmedo, por lo que las especies pueden extenderse a éstos. Las familias de macrolíquenes mejor representadas en el BHM son las Parmeliaceae, Physciaceae, Collemataceae, Cladoniaceae, Lobariaceae Peltigeraceae, y entre los microlíquenes, Bacideaceae.
Catorce familias son comunes a todos los tipos de vegetación mencionados, aunque tienden a ser más diversas en 1 o 2 de ellos. Por ejemplo, algunas bien representandas en BTE: Parmeliaceae, Physciaceae, Verrucariaceae, Porinaceae y Peltigeraceae, son también muy frecuentes en el MXE; Collemataceae y Lobariaceae, ambas de cianolíquenes, son comunes además en BHM, y Ramalinaceae ha sido recolectada también por igual en BTH y MXE. Por su parte, la familia Lichinaceae (cianolíquenes) que es muy común en MXE, también ha sido ampliamente recolectada en BES. En este último tipo de vegetación, las familias más comunes son Graphidaceae, Pertusariaceae y Arthoniaceae. Las 2 primeras también son abundantes en MXE, mientras que Arthoniaceae tiene una representación importante además en BTH. Dos familias dominantes en este bosque, Gomphillaceae y Roccellaceae, también están bien representadas en bosque templado. No se ha determinado hasta qué punto la riqueza de especies conocidas por tipo de vegetación refleja las diferencias reales en diversidad de líquenes, el esfuerzo de recolecta o ambas cosas. Por ejemplo, si se suman las especies de los 2 tipos de vegetación más estudiados en tiempos recientes, BES (386 especies) y en MXE (544 especies), tenemos que las zonas áridas del país tienen un número de especies registradas de 930, que es muy cercano a las 947 especies reportadas para el bosque templado hasta ahora.
En resumen, en el Cuadro 4 se comparan los tipos de vegetación en donde se reportan las familias con más de 20 especies. Se utilizó una escala de 1 a 5, donde el valor 1 indica la representación más alta y el 5 la más baja. Por ejemplo, Graphidaceae es más común en BES (1) y en MXE (2), que en BTE (3) y aparece menos en BTH (4) y BHM (5). Mientras que Cladoniaceae es más frecuente en BTE (1) y BHM (2) y menos en MXE (3). Los espacios en blanco se deben a que las especies de dicha familia no se desarrollan en esos sitios o se desconoce su presencia porque los colectores no incluyeron información relativa a la vegetación (alrededor del 30% de las especies recopiladas).
Presencia de las familias con más de 20 especies por tipo de vegetación
| Familia | BTE | BHM | BES | BTH | MXE |
| Parmeliaceae | 1 | 5 | 4 | 3 | 2 |
| Physciaceae | 1 | 5 | 3 | 2 | 4 |
| Graphidaceae | 3 | 5 | 1 | 4 | 2 |
| Teloschistaceae | 2 | 3 | 1 | ||
| Lecanoraceae | 1 | 4 | 3 | 2 | |
| Ramalinaceae | 1 | 4 | 3 | 2 | 2 |
| Verrucariaceae | 1 | 3 | 4 | 4 | 2 |
| Gomphillaceae | 2 | 3 | 4 | 1 | 5 |
| Roccellaceae | 2 | 4 | 4 | 1 | 3 |
| Collemataceae | 1 | 2 | 3 | 3 | 4 |
| Cladoniaceae | 1 | 2 | 3 | ||
| Arthoniaceae | 4 | 5 | 1 | 2 | 3 |
| Porinaceae | 1 | 3 | 3 | 4 | 2 |
| Lecideaceae | 1 | 3 | 2 | 4 | |
| Pilocarpaceae | 2 | 3 | 4 | 1 | |
| Pertusariaceae | 3 | 4 | 1 | 3 | 2 |
| Lobariaceae | 1 | 2 | 3 | 4 | 4 |
| Pyrenulaceae | 1 | 3 | 2 | ||
| Thelotremataceae | 1 | 2 | 1 | ||
| Bacideaceae | 2 | 1 | 2 | 3 | |
| Lichinaceae | 2 | 4 | 4 | 3 | 1 |
| Acarosporaceae | 3 | 2 | 1 | ||
| Catillariaceae | 3 | 1 | 2 | ||
| Strigulaceae | 2 | 3 | 4 | 1 | |
| Peltigeraceae | 1 | 4 | 3 | 4 | 2 |
| Megasporaceae | 1 | 2 | |||
| Ectolechiaceae | 2 | 3 | 1 | ||
| Coenogoniaceae | 2 | 3 | 3 | 1 | |
| Stereocaulaceae | 1 | 2 | |||
| Umbilicariaceae | 1 | 2 |
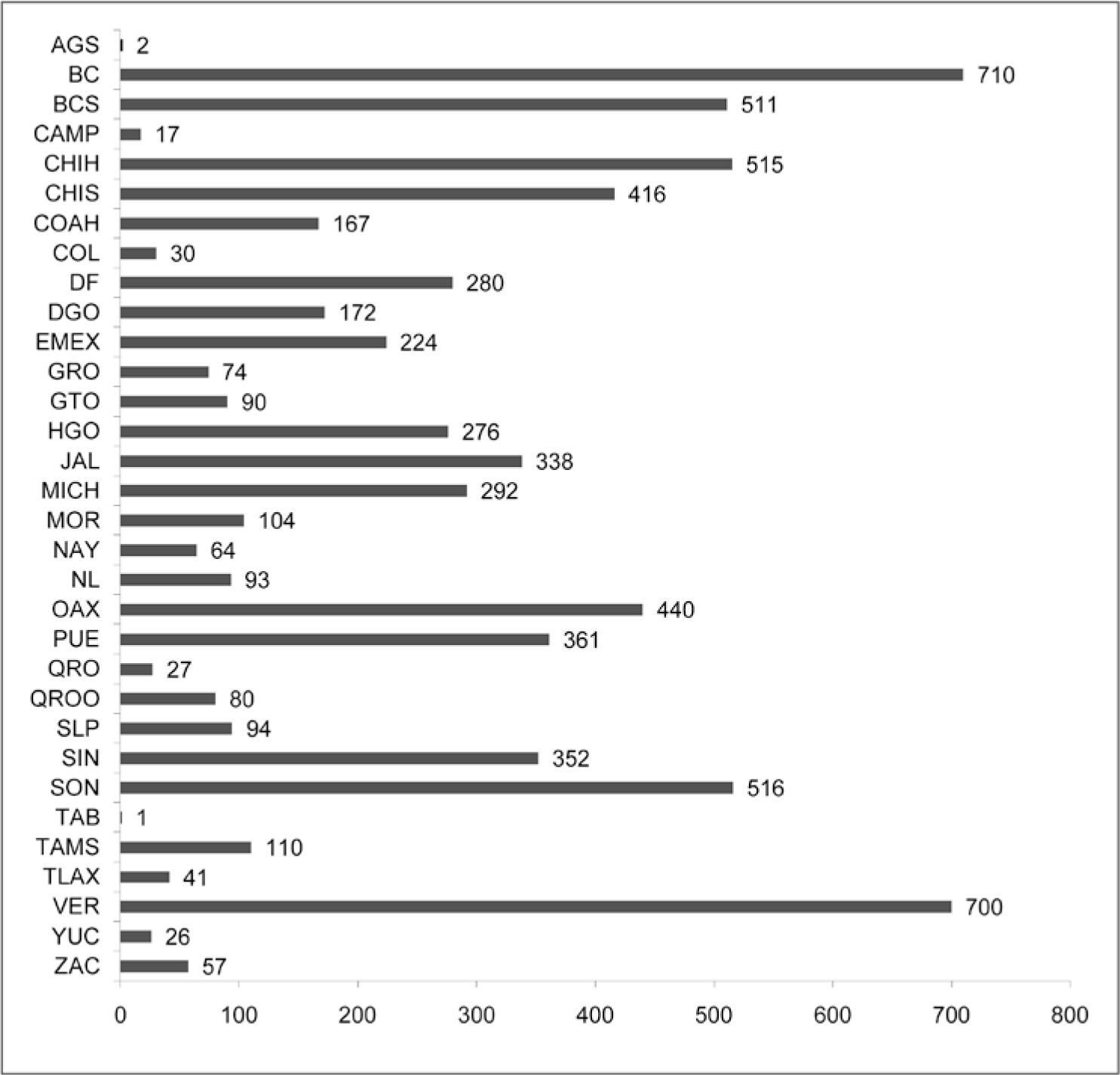
El conocimiento de la flora liquénica es muy desigual entre los estados y regiones de México (Fig. 5). En los estados del noroeste como: Baja California, Baja California Sur, Sonora, Sinaloa, Durango y Chihuahua, que abarcan el 38% de la superficie (Inegi, 2005), hay 1 342 especies, lo que representa aproximadamente el 47% del total para el país. En contraste, para la región noreste (Coahuila, Nuevo León y Tamaulipas) hay registradas sólo 295 especies. La región centro-norte (Aguascalientes, Guanajuato, Querétaro, San Luis Potosí y Zacatecas) tiene el menor número de especies reportadas (216), sobresaliendo la falta de registros en Aguascalientes, con sólo 2 especies. Estas 2 regiones del norte de México, que son predominantemente semiáridas, suman 511 especies, el 38% de las conocidas en el noroeste. Los estados occidentales (Nayarit, Jalisco, Colima y Michoacán) registran 575 especies, con mejor representación de Jalisco y Michoacán. En el oriente, en Puebla, Veracruz, Tlaxcala e Hidalgo, se detectaron 975 especies, por lo cual esta región es la segunda mejor conocida del país, siendo Veracruz el que presenta más especies (700). Las 3 entidades de la región centro-sur, Morelos, Estado de México y Distrito Federal concentran 446 especies, siendo el DF el que registra el mayor número (280); pero es necesario mencionar que se requiere una revisión más detallada de los reportes de especies realizados para la capital durante las primeras décadas del siglo XX. Para la región suroeste (Guerrero, Oaxaca y Chiapas) se conocen 732 especies, de las cuales 74 corresponden al estado de Guerrero, el cual es un estado muy poco explorado en materia de líquenes. Finalmente, la región sureste del país (Tabasco, Campeche, Quintana Roo y Yucatán) es la menos estudiada, con reportes únicamente de 112 especies.

Los números de la figura 5 reflejan los resultados de algunos estudios liquenológicos en México, principalmente los del Desierto Sonorense y proyectos en curso sobre microlíquenes de bosques secos y húmedos, así como sobre la familia Parmeliaceae. Estos trabajos han aportado información que permite entrever la enorme riqueza de las especies de líquenes del país. Sin embargo, también ponen en evidencia que se requiere realizar estudios en: 1, regiones o estados donde o no se ha recolectado o las recolectas son muy escasas; son los casos de Tabasco, Campeche, Quintana Roo y Yucatán en el sureste, y Aguascalientes y Querétaro en la región centro norte; 2, regiones conocidas por su riqueza biológica y/o por la presencia de muchas especies endémicas, como el Eje Neovolcánico, la Sierra Madre del Sur y la península de Yucatán; y 3, tipos de vegetación con recolectas escasas de líquenes, como el bosque húmedo de montaña y otros bosques tropicales húmedos como selvas medianas y manglares en diferentes estados del país.
Este trabajo agrega 1 424 especies conocidas a las 1 408 del catálogo de Ryan et al. (1996) y 12 familias cuya existencia en México se desconocía en aquel entonces: Aspidotheliaceae, Brigantiaeaceae, Eremithallaceae, Loxosporaceae, Mycoblastaceae, Microtheliopsidaceae, Mycocaliciaceae, Naetrocymbaceae, Patellariaceae, Phyllobatheliaceae, Pyrenothrichaceae y Thrombiaceae.
ConclusiónCon base en los resultados de esta revisión de la literatura liquenológica y bases de datos especializadas, en México se conocen 2 833 taxa específicos e infraespecíficos de líquenes. Es claro que existen grandes diferencias en riqueza de especies de estos organismos por tipo de vegetación, región y estados, aunque no se puede determinar todavía en qué medida se deben a desigualdades reales en la diversidad de sus líquenes o la diferente intensidad de los estudios en ellos.
Los tipos de vegetación con más especies de líquenes reportadas son el BTE (947) y el MXE (544). Les sigue el BTH (391), el bosque tropical caducifolio del BES (391) y el BHM (230).
La región más estudiada liquenológicamente es el noroeste del país, que concentra casi la mitad (47%) de las especies registradas hasta ahora para México. Le sigue el oriente con 975 especies, el suroeste (732 especies), oeste (575) y el centro-sur (446), por lo que el conocimiento de la flora liquénica en estas regiones se puede considerar como en fase intermedia. Es más escaso para las regiones NE (295 especies), centro-norte (216) y, todavía menor para el SE, con apenas 112 especies reportadas. Los datos obtenidos muestran también grandes desigualdades entre estados. Los estados con más especies de líquenes conocidas son Baja California (710 especies) y Veracruz (700), seguidos por Chihuahua, Baja California Sur y Sonora con más de 500 especies cada uno. En contraste, las cifras para Campeche (17), Aguascalientes (2) y Tabasco (1), son un fuerte indicio de que estos estados han sido muy poco explorados en cuanto a los líquenes.
Agradecemos al Dr. Fernando Álvarez Noguera, editor responsable de la RMB, por la invitación para presentar esta contribución. A la M. en C. Ma. Antonieta Arizmendi, editora técnica de RMB, la asesoría y recomendaciones para mejorarla. A la Lic. Georgina Ortega Leite por su ayuda en la búsqueda y recuperación de material bibliográfico. Al D. G. Julio César Montero Rojas su asesoría en la elaboración de las figuras. Asimismo, expresamos nuestro agradecimiento al Instituto de Biología, UNAM, a Arizona State University y a The Field Museum por el apoyo logístico otorgado durante años a la investigación de los líquenes de México. Agradecemos el financiamiento recibido de los proyectos: BSR-920-111NSF, 35008-V CONACYT, IN025808 e IN210811-3 PAPIIT. Al Dr. Claudio Delgadillo por sus oportunos comentarios a lo largo del desarrollo de este trabajo. Por la revisión del manuscrito al Dr. Mario Sousa Peña y a 2 revisores anónimos por su aporte para mejorarlo. Esta contribución no habría sido posible sin el trabajo liquenológico realizado por el Dr. Bruce Ryan (ASU) en México, lo incluimos como un reconocimiento póstumo.