






Se describen el primer y último estadios larvales y la pupa de Enagria ovata (Boheman, 1854). Al comparar el primer y último estadios de E. ovata con los de Asteriza flavicornis (Olivier, 1790), Cistudinella obducta (Boheman, 1854) y Physonota alutacea Boheman, 1854, se encontraron diferencias morfológicas en el número de tubérculos laterales (escolos), de artejos en el palpo labial y dientes en la mandíbula. La pupa es distinta a la de A. flavicornis, por la ausencia de 2 proyecciones en el pronoto. La actividad de E. ovata inicia a finales de mayo y termina a principios de noviembre. Las hembras ovipositan entre 25 y 35 huevos, las larvas eclosionan en un periodo de 9-10 días y pasan por 6 estadios larvales en un periodo de 22 días. Las larvas y los adultos fueron observados alimentándose de Montanoa tomentosa Cervant (Asteraceae).
Here we describe the first and the last immature stages and the pupa of Enagria ovata (Boheman, 1854). After a comparison between the first and last stages of E. ovata with those of Asteriza flavicornis (Olivier, 1790), Cistudinella obducta (Boheman, 1854) and Physonota alutacea Boheman, 1854, belonging to the Ischyrosonychini, we found differences in the number of scoli and teeth in the mandible. The pupa is different from that of A. flavicornis because of the absence of 2 processes in the anterior border of pronotum. Enagria ovata is present in the field from end of May to early November. Females laid 25-35 eggs, larvae hatch after 9-10 days and have 6 stages in a period of 22 days. Larvae and adults were observed on Montanoa tomentosa Cervant (Asteraceae).
La mayoría de los estudios morfológicos se enfocan en el estado adulto. Esta tendencia frecuentemente se debe a la concepción de que los adultos son la forma más completa y los estados juveniles son elementos en preparación hacia un estado adulto como resultado final (Minelli, 2003). Por ejemplo, la mayoría de las descripciones de los insectos se concentran principalmente en características de adultos y raramente incluyen características de los estados inmaduros. Sin embargo, los estados juveniles presentan características exclusivas (Costa, 2006; Meier y Shimin, 2009) y a pesar de que existen estados juveniles con importancia económica su estudio es poco atendido. Aunque se han realizado estudios en donde la inclusión de caracteres de estados juveniles ha aportado información filogenética (Beutel y Leschen, 2005; Marvaldi y Morrone, 2000; Pietrykowska-Trudruj, Stainec, Wojas y Solodovnikov, 2014), el tener el mismo estadio de todas las especies involucradas en un análisis es uno los retos principales para poder plantear hipótesis de homología con estados inmaduros (Meier y Shimin, 2009), y dado que existe falta de evidencia para poder plantear hipótesis de homología en la mayoría de análisis filogenéticos, es necesario incrementar las descripciones de los estados inmaduros. En Coleoptera se ha corroborado que el primer y último estadio, así como la pupa son las fases que aportan más caracteres informativos (Chiao y McHugh, 2000; Schultz y Meier, 1995).
Las larvas de los escarabajos se encuentran en una gran cantidad de ambientes y se pueden reconocer diferentes tipos larvales como los campodeiformes, eruciformes, escarabeiformes, ápodas o vermiformes, elateriformes y onisciformes (Costa e Ide, 2006). Dentro de Chrysomelidae varían de campodeiforme a eruciforme. Algunas especies cuentan con mecanismos químicos o físicos como medio de defensa contra sus depredadores, como espinas o escudos abdominales (Müller y Hilker, 2003). Cassidinae es una de las subfamilias que tiene características morfológicas distintivas en las larvas. Presentan tubérculos laterales (escolos) a lo largo del tórax y segmentos abdominales, y urogonfos. Como medio de defensa cubren sus urogonfos ya sea con sus exuvias o con excremento (Chaboo, 2007; Świętojańska, 2009). A pesar de las características propias de los casidinos son pocas las especies que cuentan con descripciones de sus larvas. Se ha observado que ciertos caracteres derivados de estas descripciones son útiles para análisis filogenéticos (Świętojańska, 2009).
En el presente trabajo describimos el primer y último estadios larvales de Enagria ovata (Boheman, 1854), así como también se registran datos de su biología. Además, se realizó una comparación con estados inmaduros de otras especies de la misma tribu descritas por Świętojańska y Windsor (2008), en las cuales se encontraron diferencias que podrían considerarse como caracteres diagnósticos supraespecíficos.
Materiales y métodosPara las observaciones en campo sobre el ciclo de vida de E. ovata, se visitó el cerro Chimalhuche (19°23′45″N, 98°59′18″O) localizado en el municipio de Chimalhuacán, Estado de México. Las visitas se realizaron durante los meses de mayo a diciembre del 2013. Enagria ovata fue encontrada sobre su planta huésped Montanoa tomentosa Cervant (Asteraceae). Para realizar las observaciones de su ciclo biológico se colectaron 20 ejemplares adultos. Las parejas fueron colocadas en recipientes plásticos con capacidad de 500ml. En su interior se colocó follaje fresco de M. tomentosa.
La determinación taxonómica de los adultos a nivel de género se realizó a partir de literatura especializada (Borowiec y Świętojańska, 2016; Sekerka, 2016). La determinación a nivel específica se llevó a cabo mediante la comparación con material de referencia de la Colección Nacional de Insectos del Instituto de Biología, UNAM, previamente identificada y corroborada con la descripción original. La determinación taxonómica de la planta huésped fue realizada de acuerdo con McVaugh (1984). Para la descripción del primer y último estadios, y la pupa, los ejemplares se sacrificaron en alcohol al 70%. Los tegumentos que impedían la observación de las larvas fueron retirados colocándolos en KOH al 10% en baño María durante 5min; después se retiraron y se enjuagaron con agua corriente. Posteriormente se montaron en laminillas con Euparal MTNG Medium (eucaliptol y resina de aldehído). La terminología se emplea de acuerdo a Borowiec y Świętojańska (2004), Costa e Ide (2006) y Świętojańska y Windsor (2008). Se realizaron comparaciones con estados inmaduros de otras especies de la misma tribu descritas en Świętojańska y Windsor (2008). Las observaciones y mediciones de los ejemplares revisados (=n) se hicieron utilizando un microscopio estereoscopio Carl Zeiss Discovery V8®. Las fotografías fueron tomadas con un estereoscópico ZEISS® AxioZoom V 16 y las micrografías con el microscopio electrónico de barrido Hitachi® S-2460N (15KV). Los ejemplares y laminillas utilizados para este estudio fueron depositados en la Colección Nacional de Insectos.
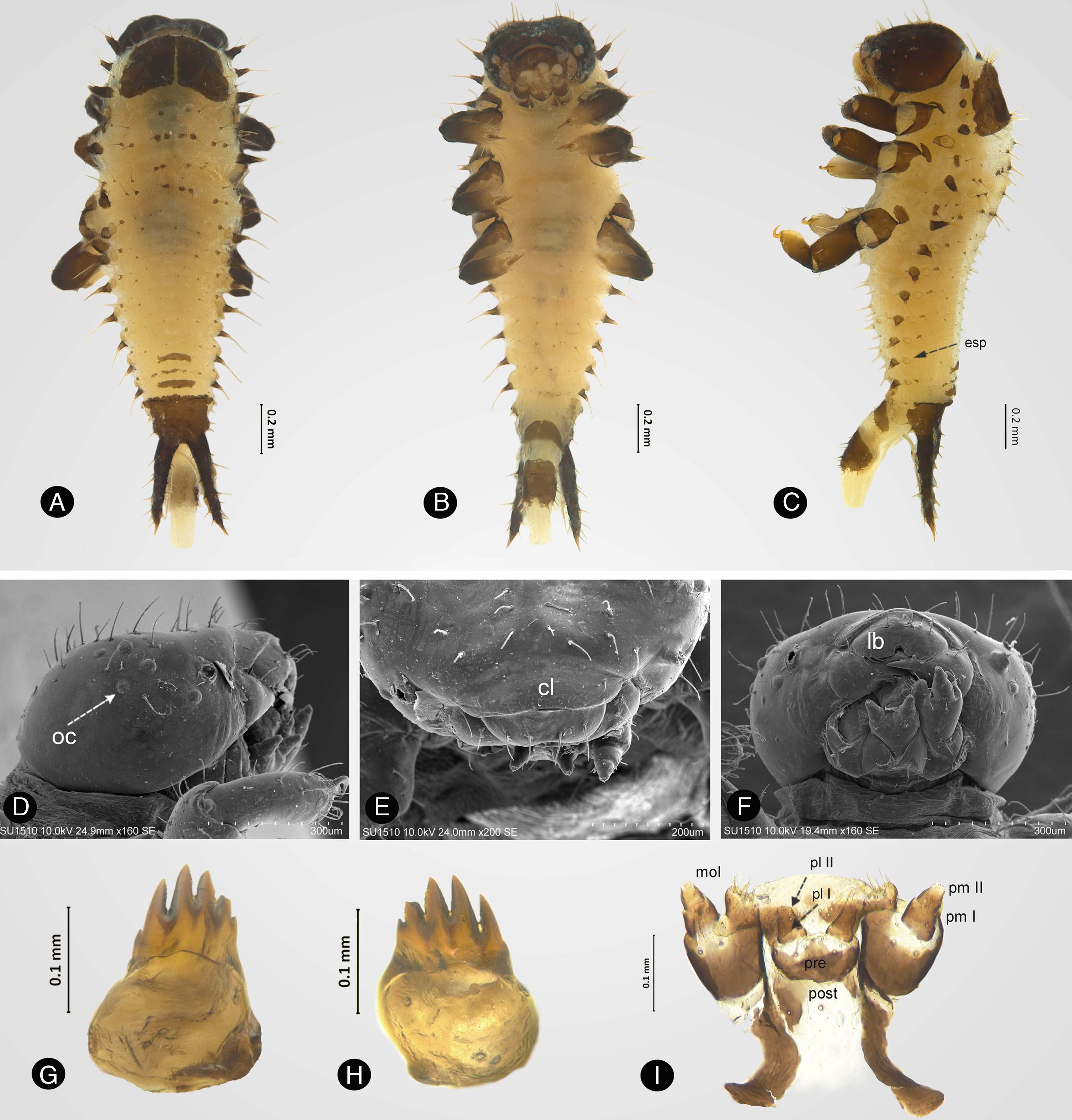
DescripciónPrimer estadio larvalMedidas (n=5; [mm]). Longitud total (desde la parte distal de la cabeza hasta el ápice de los urogonfos): 2.62, 2.3, 3.39, 2.30, 2.17. Ancho del mesonoto (parte media): 0.8, 0.64, 0.86, 0.38, 0.70. Longitud desde el borde anterior del pronoto a la base de los urogonfos: 1.76, 1.47, 2.24, 1.2, 1.44.
Cuerpo. Cilíndrico, más angosto posteriormente y más ancho en el mesonoto. De color amarillo en la parte dorsal, cubierta por sedas cortas esparcidas; pronoto con una mota negra cubriéndolo casi completamente (fig. 1A). Parte ventral completamente amarilla, cubierta por sedas esparcidas (fig. 1B). Presenta 9 pares de espiráculos, distintivamente elevados (fig. 1C). Un espiráculo en el tórax (visible solo en vista ventral) y 8 en el abdomen. Posee 12 pares de escolos laterales visibles en vista dorsal y un par de urogonfos. Escolos pardos, cónicos, sin ramificaciones laterales, cubiertos con pequeñas sedas. Cada escolo presenta una seda erecta en el ápice.
 Vista dorsal, B) vista ventral, C) vista lateral, D) ocelos (oc), E) clípeo (cl), F) labro (lb), G) mandíbula izquierda, H) mandíbula derecha, I) maxila y labio. mol: mola; pl: palpo labial; pm: palpo maxilar; post: postmentum; pre: prementum.")
Primer estadio larval de Enagria ovata. A) Vista dorsal, B) vista ventral, C) vista lateral, D) ocelos (oc), E) clípeo (cl), F) labro (lb), G) mandíbula izquierda, H) mandíbula derecha, I) maxila y labio. mol: mola; pl: palpo labial; pm: palpo maxilar; post: postmentum; pre: prementum.
Cabeza. Marrón, elipsoidal, con 6 ocelos (fig. 1D), cubierta por sedas esparcidas; antena con 2 segmentos. Clípeo glabro, más ancho que largo (fig. 1E). Labro más ancho que largo, emarginado, 8 sedas gruesas en los costados del margen anterior y 4 sedas en la superficie dorsal (fig. 1F). Mandíbulas pardas, fuertemente esclerosadas, con 4 dientes cónicos y un apéndice (fig. 1G, H). Palpo maxilar y labial conados (fig. 1I). Estipe con una seda larga y 2 cortas; mola con ápice truncado y 8 sedas con ápice agudo; palpo maxilar con 2 artejos, primero con una seda en cada costado, último con un grupo de sensilas en el ápice; palpo labial con 2 artejos, último con una agrupación de sensilas en el ápice; prementum con 2 sedas cortas dorsalmente y 2 en los costados; posmentum con 2 sedas largas y 4 cortas.
Pronoto. Presenta 2 motas pardas, superficie cubierta por sedas erectas y dispersas. Presenta 3 pares de escolos laterales, el segundo un poco más pequeño que el primero y que el tercero. Mesotórax blanco-cremoso, cubierto por sedas de diferente tamaño, con un par de escolos laterales.
Abdomen. Cubierto por sedas, algunas acomodadas en líneas horizontales. Segmentos abdominales i-vii con un par de escolos laterales; segmento viii sin escolos laterales. Urogonfos esclerosados, doblados hacia la parte dorsal; cubierto por pequeñas sedas erectas; ápice sin sedas. Patas marrón, cubiertas de sedas; parte posterior amarilla con mayor densidad de sedas; uña esclerosada, curva y aguda.
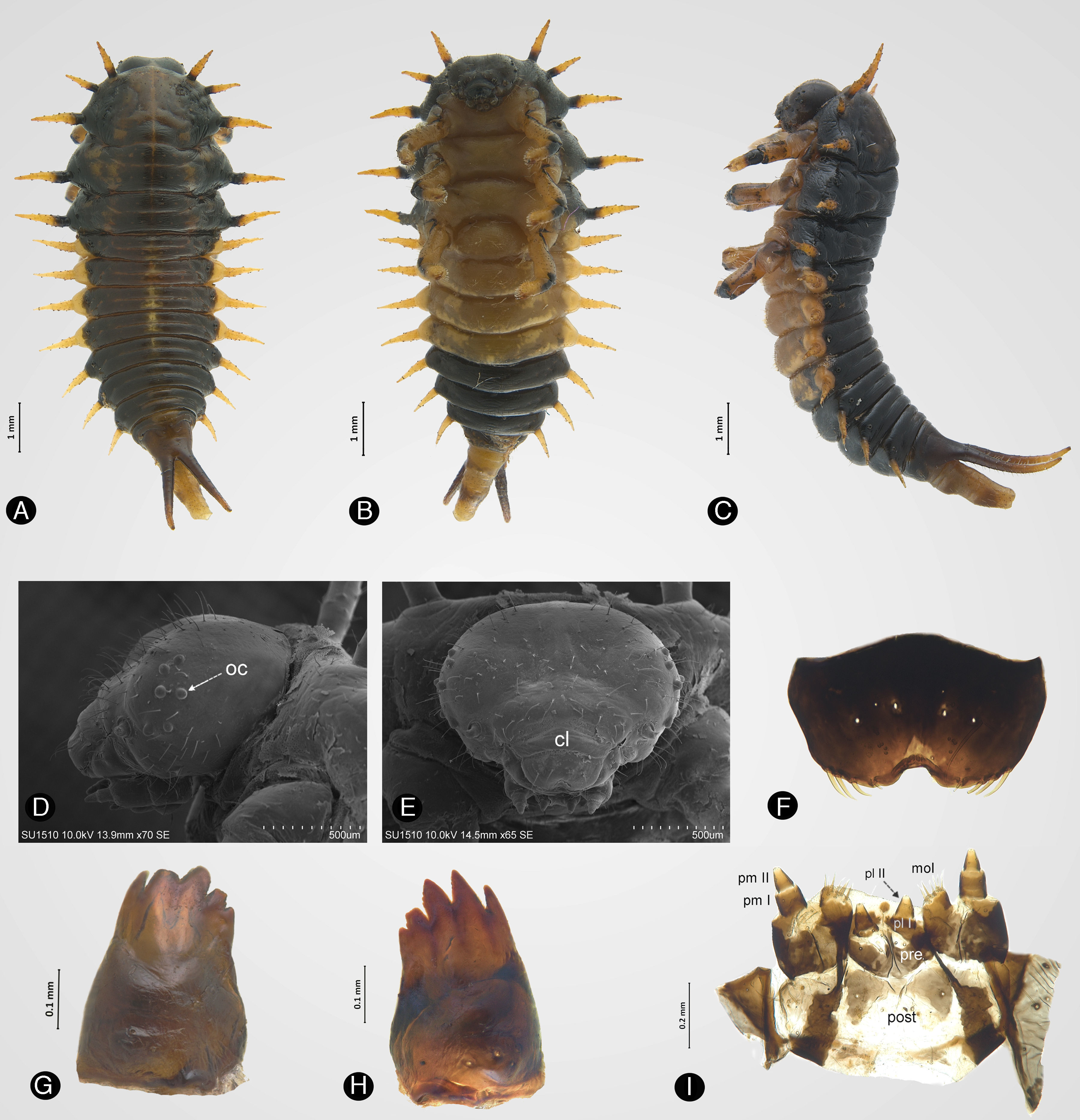
Último estadio larvalMedidas (n=5; [mm]). Longitud total (desde la parte distal de la cabeza hasta el ápice de los urogonfos): 10.9, 10, 10, 10.45, 10.18. Ancho del mesonoto (parte media): 3.27, 3.36, 3.45, 3.45, 3.45. Longitud desde el borde anterior del pronoto a la base de los urogonfos: 6.81, 6.36, 6.36, 6.63, 6.36.
Cuerpo. Ovalado, meso- y metatórax más amplios que el resto del cuerpo. Segmentos abdominales delgados. Dorso negro, parte del tegumento de coloración parda; se distingue una línea longitudinal en la parte media que llega poco después de la mitad del abdomen (fig. 2A). Parte ventral del tórax amarilla; segmentos abdominales i-iv amarillos, segmentos v-viii negros (fig. 2B). Con 12 pares de escolos laterales y un par de urogonfos amarillos. Un espiráculo en el tórax (visible solo en vista ventral) y 8 en el abdomen (fig. 1C).
 Vista dorsal, B) vista ventral, C) vista lateral, D) ocelos (oc), E) clípeo (cl), F) labro, G) mandíbula izquierda, H) mandíbula derecha, I) maxila y labio. mol: mola; pl: palpo labial; pm: palpo maxilar; post: postmentum; pre: prementum.")
Último estadio larval de Enagria ovata. A) Vista dorsal, B) vista ventral, C) vista lateral, D) ocelos (oc), E) clípeo (cl), F) labro, G) mandíbula izquierda, H) mandíbula derecha, I) maxila y labio. mol: mola; pl: palpo labial; pm: palpo maxilar; post: postmentum; pre: prementum.
Cabeza. Esclerosada, hipognata, retraída hacia el pronoto, cubierta de sedas distribuidas en toda la cabeza; sutura media completa conectada con la sutura del clípeo. Seis ocelos presentes a cada lado (fig. 2D). Antenas con 2 segmentos, el primero más ancho que el segundo. Clípeo glabro, más ancho que largo (fig. 2E). Labro liso, emarginado, más ancho que largo, con 8 sedas gruesas en los costados del margen anterior y 4 sedas en la superficie dorsal (fig. 2F). Mandíbula fuertemente esclerosada, con 6 dientes cónicos (fig. 2G, H). Palpo maxilar y labial conados (fig. 2I). Estipe con una seda larga y 4 cortas; mola con ápice truncado y 9 sedas con ápice agudo; palpo maxilar con 2 artejos, el primero con una seda en cada costado, el último con una agrupación de sensilas en el ápice; palpo labial con 2 artejos, el último con una agrupación de sensilas en el ápice; prementum con 4 sedas cortas dorsalmente; posmentum con 2 sedas largas y 4 cortas.
Pronoto. Presenta 3 pares de escolos laterales; los primeros 2 se encuentran muy cerca uno del otro, más cortos que los 2 pares siguientes, simples, junto con el resto presentan una forma cónica, cubiertos de finas sedas erectas, ápice con una seda erecta. Meso- y metatórax con 2 pares de escolos laterales.
Abdomen. Segmentos i-vii con un par de escolos laterales, los 3 últimos segmentos con escolos laterales un poco más cortos que los anteriores; el segmento viii no presenta escolos laterales. Urogonfos esclerosados, cubiertos con pequeñas sedas erectas, no más largo que el abdomen; ápice recubierto por heces fecales. Patas amarillas con una mota negra en su parte dorsal entre el fémur y la tibia, cubiertas de sedas cortas y erectas; en la parte posterior se encuentra una uña esclerosada, curva y aguda.
PupaMedidas (n=4; [mm]). Longitud total: 9.09, 8.18, 8.63, 8.63. Ancho (segundo segmento abdominal): 3.36, 3, 3.36, 3.18. Largo del pronoto: 1.8, 1.6, 1.8, 1.8. Ancho del pronoto: 4.27, 3.90, 4.27, 4.
Cuerpo. Ovalado, amarillo (fig. 3A, B). Pro-, meso- y metanoto sin escolos laterales. Superficie del pronoto cubierto por tubérculos pequeños con una seda erecta en el ápice. Segmentos abdominales con escasos tubérculos esparcidos; segmentos i-v con escolos laterales distintivos, cada escolo cubierto con tubérculos con una seda en el ápice; segmentos vi-vii con escolos laterales más pequeños y delgados que los anteriores, dirigidos hacia la parte ventral, sin tubérculos; segmento viii sin escolos. Urogonfos cubiertos por sedas erectas esparcidas.
Notas biológicas Pupa en vista dorsal de Enagria ovata, B) pupa en vista ventral, C) adulto recién emergido.")
Los adultos de E. ovata (figs. 3C y 4) inician su actividad a fínales del mes de mayo junto con las primeras lluvias de la temporada, concluyendo a principios de noviembre. La puesta de huevos sobre la planta huésped M. tomentosa corresponde al intervalo entre los primeros días de junio y principios de octubre. En cautiverio, después de cópulas sucesivas se contabilizaron hasta 3 puestas por hembra en periodos de entre 3 y 6 días. Para la puesta la hembra selecciona una hoja vigorosa. Se posiciona en dirección contraria al ápice de la hoja sobre la nervadura principal y coloca los huevos en el envés de la hoja. Del ovipositor se secreta una sustancia viscosa en forma de hilo. La hembra selecciona 2 puntos en la superficie de la hoja y comienza a construir un tipo de red. Mediante movimientos laterales con el ovipositor coloca dicha sustancia sobre la superficie del envés de la hoja. Posteriormente deposita de 2-4 huevos los cuales son cubiertos por la sustancia secretada, y repite el proceso hasta colocar de 25-35 huevos (fig. 5). Al entrar en contacto con el aire la sustancia secretada se contrae y se torna fibrosa, lo que ocasiona que la hoja tome forma de cono. Los huevos quedan en hilera dentro del cono. El proceso de oviposición tarda aproximadamente 60min y el proceso de incubación tiene una duración de 9-10 días. Después de la eclosión E. ovata pasa por 6 estadios larvales. El ciclo completo tiene una duración promedio de 22 días.


Las larvas al emerger se alejan del lugar de eclosión, permaneciendo en la misma planta huésped comienzan a alimentarse y agruparse. Presentan cicloalexia, fenómeno que consiste en la agrupación de las larvas de manera circular y el movimiento de los últimos segmentos abdominales recubiertos por excremento, el cual se distingue como una sustancia viscosa de color negro. La cicloalexia se presenta cuando las larvas se sienten atacadas. Este comportamiento se observa en los distintos estadios larvales (fig. 6). Los estadios i, ii y iii tienen una duración aproximada de 3 días cada uno. El estadio iv tiene una duración de 6 días, y el v estadio es el de mayor duración pues consta de 7 días; algunas larvas en este estadio son de coloración negra. El de menor duración es el vi, que cumple con el proceso en 2 días. La ecdisis se presenta en cada cambio de estadio y la cutícula desprendida es desechada en cada estadio larval.

La prepupa se observa después de que el estadio larval vi alcanza su máxima dimensión, tiene una duración aproximada de 12h. En este lapso dejan de alimentarse y se vuelven solitarias, buscando un lugar con poca incidencia de luz para fijarse. La pupa tiene una duración de 6-7 días (fig. 3A, B), la última muda es retenida en los últimos segmentos abdominales. El adulto recién emergido es de color amarillento con los élitros traslucidos y suaves (fig. 3C). El tiempo en que los adultos tardan para alcanzar la coloración verde metálica es de 10-15 días; sin embargo, pueden tardar hasta 40 y 70 días (Gutiérrez-Díaz, 1981).
En algunos ejemplares en estado de pupa, colectados en campo, se encontraron pupas de dípteros Tachinidae. De acuerdo con Cox (1994), los estados de larvas y pupas son más propensos a ser parasitados por Diptera e Hymenoptera.
DiscusiónLa falta de descripciones sobre la morfología de los estados inmaduros reduce el número de caracteres potencialmente útiles en los estudios sistemáticos. Por ejemplo, al compararse los primeros estadios de Cassida nobilis Linnaeus, 1758 y C. vittata Villers, 1789 se observó que cada especie poseía características propias en el primer estadio, las cuales permiten hacer una diferenciación interespecífica (Świętojańska, 2005). Al comparar las descripciones del primer estadio de E. ovata con los de Cistudinella obducta (Boheman, 1854), Physonota alutacea Boheman, 1854 y Asteriza flavicornis (Olivier, 1790) se encontraron diferencias en el número de escolos laterales y en el de artejos en el palpo labial. El primer y último estadios de E. ovata presentan 12 pares de escolos y 2 artejos labiales, P alutacea, A. flavicornis y C. obducta tienen solo un artejo en el palpo labial; P. alutacea tiene 14 pares de escolos, A. flavicornis 15 y C. obducta 16 pares. El número de dientes en la mandíbula del primer estadio también difiere, P. alutacea tiene 5 dientes triangulares y un apéndice. En los últimos estadios de E. ovata y A. flavicornis el número de dientes en las mandíbulas es distinto, E. ovata presentan 6 dientes y A. flavicornis 4 dientes distintivos y 5 más pequeños retráctiles. La pupa de E. ovata es distinta a la de A. flavicornis por no tener 2 proyecciones en el borde anterior del pronoto.
El estudio de los estados inmaduros aporta caracteres que pueden utilizarse para discriminar entre especies estrechamente relacionadas o diferenciar entre distintos géneros; incluso pueden reflejar relaciones de parentesco como los sugeridos por Świętojańska y Windsor (2008) para los géneros Physonota, Cistudinella y Asteriza. La consideración de caracteres de estados inmaduros en análisis filogenéticos podrá ayudar a tener una mejor resolución en la reconstrucción de relaciones de parentesco, ya que ofrecen información filogenética que se pierde en el estado adulto.
Además, el reconocimiento de los estados inmaduros de los insectos es de gran importancia en áreas de interés económico. En la mayoría de los casos las formas larvales presentan hábitos alimenticios totalmente distintos a los adultos, por lo que con el estudio integral de la morfología y biología de las diferentes etapas de desarrollo se podría reconocer a una especie a pesar de la ausencia del adulto.
A Santiago Zaragoza Caballero (IB, UNAM) por sus comentarios al manuscrito. A María Magdalena Ayala (Herbario-FEZA, UNAM) por la determinación taxonómica de Montanoa tomentosa. A M. Berenit Mendoza (IB, UNAM) por la toma de micrografías de microscopía electrónica. A 2 revisores anónimos por sus valiosos comentarios.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.