




Cosmos es uno de los géneros monofiléticos más diversos de la tribu Coreopsideae (Asteraceae). El grupo incluye 35 especies y 4 taxa infraespecíficos restringidos al continente americano. El objetivo del presente trabajo fue evaluar la distribución geográfica y la riqueza de Cosmos a través de un sistema de información geográfica (SIG). Se estimó el área de distribución geográfica de los taxa con más de un registro. También se evaluó la riqueza de las especies de Cosmos por región política, provincia geológica y una cuadrícula de 80×80km. Por último, se valoraron los patrones de distribución geográfica por hábito de crecimiento. Se encontraron 3 zonas de alta riqueza, todas en México, en los estados de Jalisco, Durango y México. Las hierbas anuales, las hierbas perennes y los sufrútices mostraron patrones de distribución geográfica diferente. Las hierbas anuales tuvieron la mayor área de distribución. En contraste, las hierbas perennes presentaron mayor cantidad de especies pero una distribución restringida. El Eje Neovolcánico ha sido el escenario de la diversificación de Cosmos y es la poliploidización el principal mecanismo de especiación.
Cosmos is a monophyletic genus and one of the most diverse groups within the tribe Coreopsideae (Asteraceae). It includes 35 species and 4 infraspecific taxa found in the Americas. The aim of this study was to evaluate the geographical distribution and richness of Cosmos using a geographical information system (GIS). We estimated the geographic range of taxa with more than one record. The species richness was analyzed by political region, geological province, biogeographical province and an 80×80km cell grid. Finally, we assessed the geographical distribution patterns of Cosmos according to growth habit. We found 3 areas of greatest richness, all in México, in the states of Jalisco, Durango and Mexico. Annual herbs, perennials herbs and subshrub had different patterns of geographical distribution. The annual herbs were the most widespread. In contrast, the perennial herbs were more diverse but exhibited a restricted distribution. The Trans-Mexican Volcanic Belt has been the scenario of Cosmos diversification and poliploidy seems to have been the primary mechanism of speciation.
La familia Asteraceae es el grupo de angiospermas más diverso en México y cerca del 63% de sus especies son endémicas (Turner y Nesom, 1998; Villaseñor, 1993). Un ejemplo al caso es la tribu Coreopsideae (Less.) Turner et Powell, con más del 50% de los géneros en México, esto es, 13 de los 24 géneros (Panero y Funk, 2002; Crawford et al. 2009). Además, 194 de las 590 especies se encuentran en México. A su vez, un buen ejemplo de la diversidad y endemismo de Coreopsideae en este país es el género Cosmos Cav., uno de los grupos monofiléticos más diversos de la tribu (Crawford et al., 2009). Las cabezuelas en Coreopsideae tienen un involucro con 2 series de filarias diferenciadas por color, consistencia y forma, canales resiníferos conspicuos en las filarias internas y la presencia de pigmentos flavonoides (Crawford y Stuessy, 1981; Funk et al., 2009; Robinson, 1981; Ryding y Bremer, 1992). Cosmos se distingue de otros géneros de Coreopsideae por presentar los filamentos de los estambres pilosos (Sherff, 1932, 1955; McVaugh, 1984; Funk et al., 2009).
Cosmos está formado por 35 especies y 4 variedades. Se distribuye desde el suroeste de Estados Unidos de América hasta el norte de Argentina, pero en México crecen 33 especies y las 4 variedades. El género se divide en 3 secciones (Sherff, 1932): 1) Cosmos sección Cosmos Sherff; 2) Cosmos sección Mesinenia Sherff y 3) Cosmos sección Discopoda (DC.) Sherff. Cosmos sección Cosmos agrupa 5 especies anuales con cabezuelas de 8 flores liguladas de color blanco, violado o anaranjado y aquenios pubescentes de rostro largo (Figs. 1 A-F). En contraste, Cosmos sección Mesinenia incluye 8 especies sufruticosas que producen cabezuelas con 5–8 flores liguladas de color blanco, violado o amarillo y aquenios pubescentes o glabros de rostro corto (Figs. 1 G-K). Por último, Cosmos sección Discopoda está representada por 22 especies de hábitos herbáceos, perennes, con raíces tuberosas, cabezuelas con 8–10 flores liguladas, blancas, violáceas o púrpuras y aquenios pubescentes o glabros de rostro exiguo (Figs. 1 L-Z). Todas las especies perennes tienen rizomas, pero el tipo de raíz que produce el rizoma es diferente: en Cosmos sección Mesinenia, son axonomorfas, mientras que en Cosmos sección Discopoda son tuberosas.
, Cosmos bipinnatus; (B), Cosmos caudatus; (C), Cosmos pacificus; (D), Cosmos parviflorus en vista frontal; (E), Cosmos parviflorus en vista posterior; (F), Cosmos sulphureus. Capítulos de sufrútices: (G), Cosmos carvifolius; (H), Cosmos crithmifolius; (I), Cosmos intercedens; (J), Cosmos landii var. landii; (K), Cosmos ochroleucoflorus. Capítulos de hierbas perennes: (L), Cosmos diversifolius var. diversifolius; (M), Cosmos jaliscensis; (N), Cosmos juxtlahuacensis; (O), Cosmos mcvaughii; (P), Cosmos modestus; (Q), Cosmos montanus; (R), Cosmos nitidus; (S), detalle de las flores del disco de Cosmos nitidus; (T), Cosmos purpureus; (U), detalle de las flores del disco de Cosmos purpureus; (V), Cosmos scabiosoides; (W), detalle de las flores del disco de Cosmos scabiosoides; (X), Cosmos schaffneri; (Y), Cosmos sessilis; (Z), Cosmos stellatus. Fotografías de G. Vargas-Amado (A-I, K-X, Z), A. Rodríguez (J) y A. Castro-Castro (Y).")
Capítulos de las hierbas anuales: (A), Cosmos bipinnatus; (B), Cosmos caudatus; (C), Cosmos pacificus; (D), Cosmos parviflorus en vista frontal; (E), Cosmos parviflorus en vista posterior; (F), Cosmos sulphureus. Capítulos de sufrútices: (G), Cosmos carvifolius; (H), Cosmos crithmifolius; (I), Cosmos intercedens; (J), Cosmos landii var. landii; (K), Cosmos ochroleucoflorus. Capítulos de hierbas perennes: (L), Cosmos diversifolius var. diversifolius; (M), Cosmos jaliscensis; (N), Cosmos juxtlahuacensis; (O), Cosmos mcvaughii; (P), Cosmos modestus; (Q), Cosmos montanus; (R), Cosmos nitidus; (S), detalle de las flores del disco de Cosmos nitidus; (T), Cosmos purpureus; (U), detalle de las flores del disco de Cosmos purpureus; (V), Cosmos scabiosoides; (W), detalle de las flores del disco de Cosmos scabiosoides; (X), Cosmos schaffneri; (Y), Cosmos sessilis; (Z), Cosmos stellatus. Fotografías de G. Vargas-Amado (A-I, K-X, Z), A. Rodríguez (J) y A. Castro-Castro (Y).
Villaseñor (1991) analizó la riqueza de especies endémicas de la tribu Heliantheae en México, incluyendo a Coreopsideae, considerada como subtribu. Además de cuantificar la riqueza por estados; este autor (1991) también localizó centros de concentración de endemismos en los estados de Baja California Sur, Chiapas, Coahuila y Jalisco. Esta última entidad acumuló la mayor cantidad de especies restringidas y la zona montañosa occidental que incluye las sierras de Cuale, Mascota y Cacoma, fue el área de mayor concentración de endemismos. En el análisis de Villaseñor (1991) no se empleó un SIG y en vista de la abundancia de datos, se analizaron 4 estados, que sólo representan el 20% de la superficie continental de México. No obstante, en las últimas 2 décadas la obtención rápida de resultados fundamentados en bases de datos grandes ha sido complementada con la introducción de herramientas computacionales y el empleo de los sistemas de información geográfica (SIG). Con la información obtenida de las colecciones biológicas y analizada con estas nuevas herramientas, ha sido posible valorar la diversidad biológica de un territorio de forma más rigurosa, fácil, rápida y relativamente económica (Paterson et al., 2000).
Los SIG se utilizan con varios propósitos; por ejemplo, permiten la evaluación de la riqueza y estiman las áreas de distribución geográfica de las especies (Hijmans y Spooner, 2001; Parthasarathy et al., 2006; Scheldeman et al., 2007). También son útiles para analizar las características morfológicas en un contexto geográfico. Abraham et al. (2010) analizaron la variación de las características morfológicas cuantitativas de la vaina en Vigna mungo (L.) Hepper, una especie de importancia alimenticia en India. Sunil et al. (2009) analizaron la distribución de las características morfológicas de Jatropha curcas L., una especie naturalizada en India y de importancia económica. Los mapas basados en cuadrículas de Abraham et al. (2010) y Sunil et al. (2009) mostraron los sitios geográficos que presentan la mayor riqueza de plantas con las características deseables.
El entendimiento de la distribución de las especies también es necesario para su conservación (Scheldeman et al., 2007). Por ejemplo, Cosmos jaliscensis Sherff, C. juxtlahuacensis Panero et Villaseñor, C. longipetiolatus Melchert, C. mattfeldii Sherff, C. nelsonii B. L. Rob. et Fernald y C. sherffii Melchert tienen una distribución geográfica restringida (McVaugh, 1984; Melchert, 1968; Panero y Villaseñor, 1996; Sherff, 1964). De forma similar, a C. diversifolius Otto ex Knowles et Westc. var. pumilus Sherff, C. herzogii Sherff, C. langlassei Sherff, C. linearifolius (Sch. Bip.) Hemsl. var. magnifolius Sherff, C. linearifolius (Sch. Bip.) Hemsl. var. maysilessi Sherff y C. nitidus Paray se les conoce sólo de la localidad tipo (McVaugh, 1984; Melchert, 1968). Sin embargo, ninguna de estas especies está incluida en la lista de especies en riesgo de la norma oficial mexicana Nom-059-Semarnat-2010. Tampoco están incluidas en la lista roja de las especies amenazadas de la Unión Internacional para la Conservación de la Naturaleza (IUCN, 2010). Por este motivo, el presente trabajo es una contribución al estudio de la distribución geográfica de las especies de Cosmos, como un punto de partida para proponer su estatus de amenaza y posteriormente sugerir estrategias para su conservación.
El objetivo del presente trabajo es evaluar la distribución geográfica y la riqueza de las especies de Cosmos mediante los datos de las colecciones biológicas y el empleo de los SIG. También se pretende encontrar los patrones de distribución geográfica de Cosmos relacionados con su hábito de crecimiento. La relevancia de los análisis de riqueza radica en que son un punto de partida para la identificación de regiones de endemismo y son útiles para determinar regiones prioritarias para la conservación. La importancia de la metodología biogeográfica radica en intentar conservar el mayor número de especies en el menor número de áreas (Rebelo y Siegfried, 1992; Willis et al.,1996;Hijmans et al., 2004). Al identificar los patrones de riqueza y endemismo de Cosmos, posteriormente se podrán valorar las áreas que han sido designadas como prioritarias para la conservación de la biodiversidad (Villaseñor, 2004).
Materiales y métodosBase de datos. Se construyó una base de datos a partir de la información disponible en las etiquetas de los ejemplares de herbario. Los datos se agruparon en 4 categorías: 1) datos taxonómicos, incluidos familia, género, epíteto específico, autor, variedad y autor de la variedad; 2) datos geográficos como país, estado, municipio, localidad, altitud, latitud y longitud; 3) datos ecológicos tales como hábitat, observaciones y fecha de colecta y 4) datos curatoriales como nombre y número del colector, nombre del determinador y nombre de la colección.
Se revisaron ejemplares de 12 colecciones botánicas: herbarios de los Centros Interdisciplinarios de Investigación para el Desarrollo Integral Regional del Instituto Politécnico Nacional, en Durango (CIIDIR) y en Michoacán (CIMI), Herbario de la Escuela Nacional de Ciencias Biológicas del Instituto Politécnico Nacional (ENCB), Herbario de Universidad Autónoma del Estado de Morelos (HUMO), Herbario Luz María Villarreal de Puga del Instituto de Botánica de la Universidad de Guadalajara (IBUG), Herbario del Instituto de Ecología, A.C., Centro Regional del Bajío (IEB), Herbario del Instituto Nacional de Estadística y Geografía (INEGI), Herbario Nacional del Instituto de Biología de la Universidad Nacional Autónoma de México (MEXU), Herbario J. Rzedowski de la Universidad Autónoma de Querétaro (QMEX), Herbario del Instituto de Ecología, A.C., Xalapa (XAL), Herbario de la Universidad Veracruzana (XALU) y Herbario del Centro Universitario de la Costa Sur de la Universidad de Guadalajara (ZEA).
Se consultaron bases de datos electrónicas para obtener los registros de Cosmos de Estados Unidos de América, Centroamérica y Sudamérica. Una base de datos importante es Tropicos, del Jardín Botánico de Missouri (Tropicos. org, 2011). La base de datos de la flora de Texas (Flora of Texas Consortium, 2007) contiene los registros del Herbario Lundell de la Universidad de Texas (LL-TEX). De forma similar, mediante la base de datos de Southwest Environmental Information Network (SEINet, 2010) se buscaron registros de Cosmos en las siguientes colecciones: University of Arizona Herbarium (ARIZ), University of California, Riverside Plant Herbarium (UCR), Arizona State University Vascular Plant Herbarium (ASU), New York Botanical Garden (NY), Intermountain Herbarium, Utah State University (UTC), Deaver Herbarium, Northern
Arizona University (ASC), Desert Botanical Garden Herbarium Collection (DES), Herbario de la Universidad de Sonora (USON), Navajo Nation Herbarium (NAVA), Cochise County Herbarium (COCHISE), TEUI Herbarium, US Forest Service Southwestern Region (TES), Colorado State University Herbarium (CS), University of New Mexico Herbarium (UNM), NMSU Center for Natural History Collections Herbarium (NMC), NMSU Center for Natural History Collections Range Science (NMCR), ENMU Natural History Collection Herbarium (ENMU) y Gila Center for Natural History Herbarium (SNM). Cuando se detectaron registros con identidad taxonómica dudosa, se solicitaron los ejemplares digitalizados a los encargados de los herbarios correspondientes.
Para utilizar la información, la base de datos fue convertida a un formato compatible con un SIG (Hijmans et al., 1999) mediante el programa de cómputo ArcView 3.3 (ESRI, 1992–2002). Los registros sin coordenadas fueron georreferenciados. Para obtener las coordenadas de los registros mexicanos, se empleó la cartografía digital de la Comisión nacional para el conocimiento y uso de la biodiversidad (Conabio) y la Comisión nacional forestal (Conafor). Se utilizaron los mapas de las localidades, los municipios y la hidrografía de México (Maderey y Torres-Ruata, 1990). También se emplearon los mapas de curvas de nivel (Conabio, 1998) y la división política estatal de México (Conabio, 2005). De forma análoga, para obtener las coordenadas geográficas fuera del territorio mexicano, se empleó la información de la Base de Datos Global de las Áreas Administrativas (GADM, 2011) e imágenes de satélite disponibles en la red.
La base de datos fue depurada con el fin de eliminar registros repetidos y disminuir errores de georreferenciación (Hijmans et al., 1999). Con ArcView 3.3 se verificó la proyección de los puntos mediante su superposición con los mapas de límites administrativos en 2 niveles: 1) los países del continente americano y 2) la división administrativa de cada país. Cuando se encontraron discrepancias, se revisó la descripción de la localidad y se asignaron nuevas coordenadas cuando así se requirió. Los registros con localidades ambiguas o repetidas fueron eliminados. Finalmente, para facilitar los cálculos de las áreas, toda la cartografía empleada y el mapa de las localidades de Cosmos fueron transformados a una proyección Azimutal Equivalente de Lambert usando como meridiano central −102° y el ecuador como latitud de referencia. Esta proyección mantiene la exactitud en las medidas de las áreas y muestra las direcciones correctas desde el punto central del mapa (Snyder, 1987).
Área de distribución. La distribución geográfica de las especies de Cosmos se estimó con el uso del programa de cómputo DIVA-GIS 4 (Hijmans et al., 2004). Se ejecutaron los siguientes cálculos para cada taxón: 1) la máxima distancia observada entre 2 registros, ya que su valor representa la mayor distancia observada entre todos los posibles pares y permite conocer la extensión del área geográfica; 2) la extensión de la presencia (EOO), que es el área contenida dentro del polígono de menor superficie que contenga todos los sitios que ocupa un taxón (IUCN Standards and Petitions Subcommittee, 2010); esta medida se calculó con el polígono convexo mínimo, eliminando las áreas del polígono fuera de la parte continental; 3) el área de ocupación (AOO), que es el área actualmente ocupada por un taxón que se encuentra dentro de la extensión de la presencia y representa el hábitat adecuado para la especie, se calculó empleando una cuadrícula. El tamaño de celda fue de 2×2km, que es la escala apropiada para permitir que un taxón pueda ser categorizado como “en peligro crítico de extinción” según la UICN (IUCN Standards and Petitions Subcommittee, 2010). Según Willis et al. (2003), AOO es la medida más adecuada para evaluar la información de los ejemplares de herbario. Además, con el fin de identificar tendencias en el rango de distribución geográfica respecto a su hábito de crecimiento, se comparó la EOO y el AOO de las hierbas anuales, las hierbas perennes y los sufrútices. Cosmos herzogii y C. langlassei no fueron incluidas en el análisis comparativo del área de distribución geográfica por hábito de crecimiento porque no fueron colectadas y la información sobre su hábito de crecimiento es confusa. Los valores de EOO y AOO se analizaron con base en los rangos geográficos propuestos por la IUCN (IUCN Standards and Petitions Subcommittee, 2010) para las diferentes categorías de riesgo.
Riqueza de especies. Se cuantificó la cantidad de especies de Cosmos por: 1) países del continente Americano, 2) división administrativa por país (GADM, 2011), 3) provincias geológicas propuestas por el U.S. Geological Survey (USGS) (Osmonson et al., 2000) y 4) cuadrícula. El tamaño de celda se eligió de acuerdo con el criterio de Willis et al. (2003), con la modificación de Suárez-Mota y Villaseñor (2011). Con base en esta metodología, se promedió la máxima distancia de cada taxón y el valor se dividió entre 10. El cociente fue usado para definir la longitud de la celda.
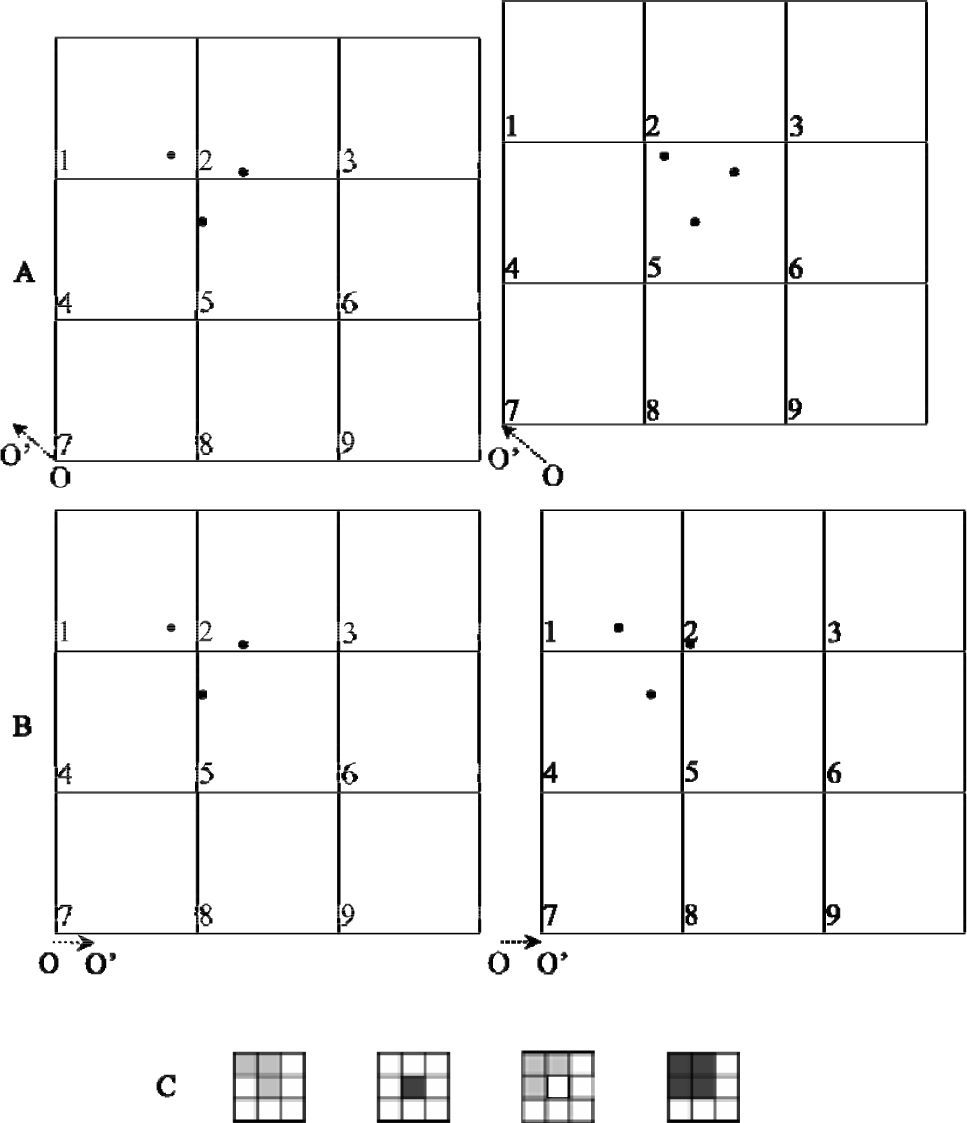
El origen de la cuadrícula es arbitrario e influencia los resultados. Por esta razón, se asignó un registro a más de una celda, en aquellos casos en que el registro se localizó adyacente a otra celda (Fig. 2). Los registros fueron asignados a las celdas utilizando una zona circular (vecindad circular) con un radio igual al tamaño de la celda. El tamaño de celda fue de 80×80km, es decir, un área de 6 400km2 y el área circular analizada para la asignación de taxa a cada celda fue de aproximadamente 20 106km2. La consecuencia del empleo de este método es la sobreestimación del área en la que una especie puede ocurrir. Sin embargo, el empleo de vecindad circular muestra los patrones constantes de riqueza de especies, sin importar el origen de la cuadrícula (Fig. 2).
, cuando el origen de la cuadrícula está en O, las celdas 1, 2 y 5 contienen una especie cada una. Si el origen de la celda se mueve a la posición O")
El punto de origen de la cuadrícula puede modificar los resultados. (A), cuando el origen de la cuadrícula está en O, las celdas 1, 2 y 5 contienen una especie cada una. Si el origen de la celda se mueve a la posición O', ahora sólo la celda 5 contiene la diversidad estudiada. (B), si el origen de la cuadrícula se desplaza de O a O', las celdas 1, 2 y 4 son las que contienen la diversidad, con una especie cada una. (C), diferentes patrones de riqueza por cuadrícula. Las 3 primeras cuadrículas se hicieron sin el método de la vecindad circular y muestran patrones de riqueza desiguales debido a que el punto de origen es diferente. Al aplicar el método de vecindad circular, el punto de origen de la cuadrícula no influye en el patrón de riqueza, como se muestra en la última cuadrícula. Los tonos de sombreado son proporcionales a la cantidad de especies en las celdas.
Los resultados de la cuadrícula fueron sintetizados en un cuadro y comparados por provincia geológica. Este análisis permite conocer cuántas celdas existen por provincia y en cuántas se registra la presencia de especies para generar el promedio de especies por celda. De esta manera, se puede calcular el porcentaje de la provincia geológica ocupada por celdas con uno o más taxa y estimar el porcentaje de taxa en la celda con mayor riqueza.
Por último, se cuantificó la riqueza de taxa por latitud, longitud y altitud. Para el cálculo del número de taxa por latitud y longitud, se crearon 2 nuevas cuadrículas en el programa de cómputo DIVA-GIS 4. En la primera, se ajustó la resolución a una columna para crear filas de 80km correspondientes a la latitud. De forma análoga, en la segunda se generó una fila, para obtener columnas de 80km de longitud. Finalmente, se graficó la cantidad de taxa por paralelo (latitud) y meridiano (longitud). Para calcular la cantidad de especies por altitud, los registros se agruparon arbitrariamente en clases de 250m y se graficó la cantidad de taxones presentes en cada clase (Hijmans y Spooner, 2001). Los registros sin altitud, se completaron con los datos obtenidos de cuadrículas de elevación a una resolución de 30 segundos extraídos de la base de datos del Consorcio para la Información Espacial (CGIAR-CSI, 2011).
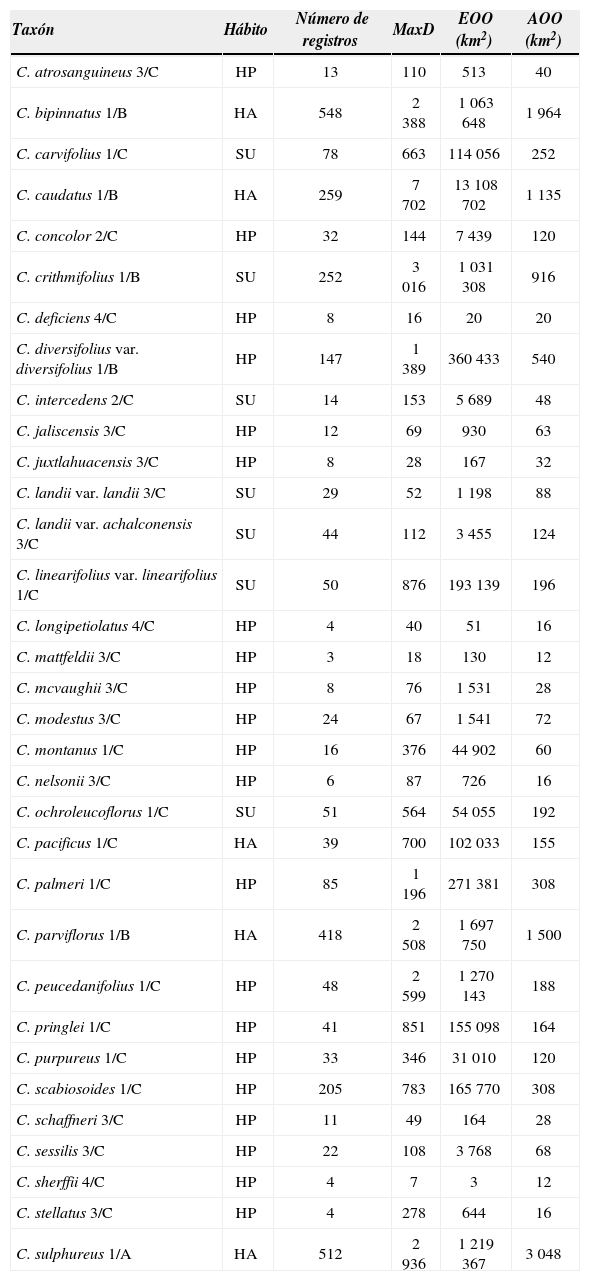
ResultadosBase de datos. Se obtuvieron 3 416 registros de 35 especies y 4 variedades. Se eliminaron 382 registros que contenían localidades repetidas para la misma especie o porque las localidades fueron ambiguas o dudosas. De esta manera, el número de registros analizados fue de 3 034. Así mismo, el número de registros por especie varió de uno en 6 especies hasta 548 en Cosmos sulphureus Cav. (Cuadro 1).
Resumen de las áreas de distribución geográfica de las especies de Cosmos con más de 1 registro. El número y letra antes y después de la diagonal sintetiza los valores de EOO y AOO, respectivamente: (1) mayor de 20 000km2, (2) menor de 20 000km2, (3) menor de 5 000km2, (4) menor de 100km2; (A) mayor de 2 000km2, (B) menor de 2 000km2; (C) menor de 500km2, (D) menor de 10km2. Abreviaturas: AOO (el área de ocupación del taxón), EOO (área estimada de la presencia de un taxón), MaxD (máxima distancia entre los 2 registros más distantes), HP (hierba perenne), HA (hierba anual), SU (sufrútice)
| Taxón | Hábito | Número de registros | MaxD | EOO (km2) | AOO (km2) |
|---|---|---|---|---|---|
| C. atrosanguineus 3/C | HP | 13 | 110 | 513 | 40 |
| C. bipinnatus 1/B | HA | 548 | 2 388 | 1 063 648 | 1 964 |
| C. carvifolius 1/C | SU | 78 | 663 | 114 056 | 252 |
| C. caudatus 1/B | HA | 259 | 7 702 | 13 108 702 | 1 135 |
| C. concolor 2/C | HP | 32 | 144 | 7 439 | 120 |
| C. crithmifolius 1/B | SU | 252 | 3 016 | 1 031 308 | 916 |
| C. deficiens 4/C | HP | 8 | 16 | 20 | 20 |
| C. diversifolius var. diversifolius 1/B | HP | 147 | 1 389 | 360 433 | 540 |
| C. intercedens 2/C | SU | 14 | 153 | 5 689 | 48 |
| C. jaliscensis 3/C | HP | 12 | 69 | 930 | 63 |
| C. juxtlahuacensis 3/C | HP | 8 | 28 | 167 | 32 |
| C. landii var. landii 3/C | SU | 29 | 52 | 1 198 | 88 |
| C. landii var. achalconensis 3/C | SU | 44 | 112 | 3 455 | 124 |
| C. linearifolius var. linearifolius 1/C | SU | 50 | 876 | 193 139 | 196 |
| C. longipetiolatus 4/C | HP | 4 | 40 | 51 | 16 |
| C. mattfeldii 3/C | HP | 3 | 18 | 130 | 12 |
| C. mcvaughii 3/C | HP | 8 | 76 | 1 531 | 28 |
| C. modestus 3/C | HP | 24 | 67 | 1 541 | 72 |
| C. montanus 1/C | HP | 16 | 376 | 44 902 | 60 |
| C. nelsonii 3/C | HP | 6 | 87 | 726 | 16 |
| C. ochroleucoflorus 1/C | SU | 51 | 564 | 54 055 | 192 |
| C. pacificus 1/C | HA | 39 | 700 | 102 033 | 155 |
| C. palmeri 1/C | HP | 85 | 1 196 | 271 381 | 308 |
| C. parviflorus 1/B | HA | 418 | 2 508 | 1 697 750 | 1 500 |
| C. peucedanifolius 1/C | HP | 48 | 2 599 | 1 270 143 | 188 |
| C. pringlei 1/C | HP | 41 | 851 | 155 098 | 164 |
| C. purpureus 1/C | HP | 33 | 346 | 31 010 | 120 |
| C. scabiosoides 1/C | HP | 205 | 783 | 165 770 | 308 |
| C. schaffneri 3/C | HP | 11 | 49 | 164 | 28 |
| C. sessilis 3/C | HP | 22 | 108 | 3 768 | 68 |
| C. sherffii 4/C | HP | 4 | 7 | 3 | 12 |
| C. stellatus 3/C | HP | 4 | 278 | 644 | 16 |
| C. sulphureus 1/A | HA | 512 | 2 936 | 1 219 367 | 3 048 |
Área de distribución. Cosmos tiene sus límites naturales de distribución en América. Las especies de este género crecen desde los 38.1° N, en el centro del estado de Colorado, Estados Unidos de América, hasta los 26.7° S, en la provincia de Tucumán, en el norte de Argentina por la vertiente del Pacífico y los 25.5° S, en el estado de Paraná, en el sur de Brasil, por la vertiente del Atlántico. Su distribución inicia en el centro de la cordillera de Rocky Mountains, Estados Unidos de América, cruza México a través de las sierras Madre Occidental, Oriental y del Sur y finaliza en el sur de los Andes. Cosmos parviflorus (Jacq.) Pers. presenta la distribución más boreal, mientras que C. caudatus Kunth, C. herzogii y C. peucedanifolius Wedd. son encontradas en las regiones más australes. Por su parte, las especies con preferencias tropicales como C. caudatus, C. pacificus Melchert y C. sulphureus se encuentran además en las islas del Caribe y planicies costeras de México, Centroamérica y Sudamérica.
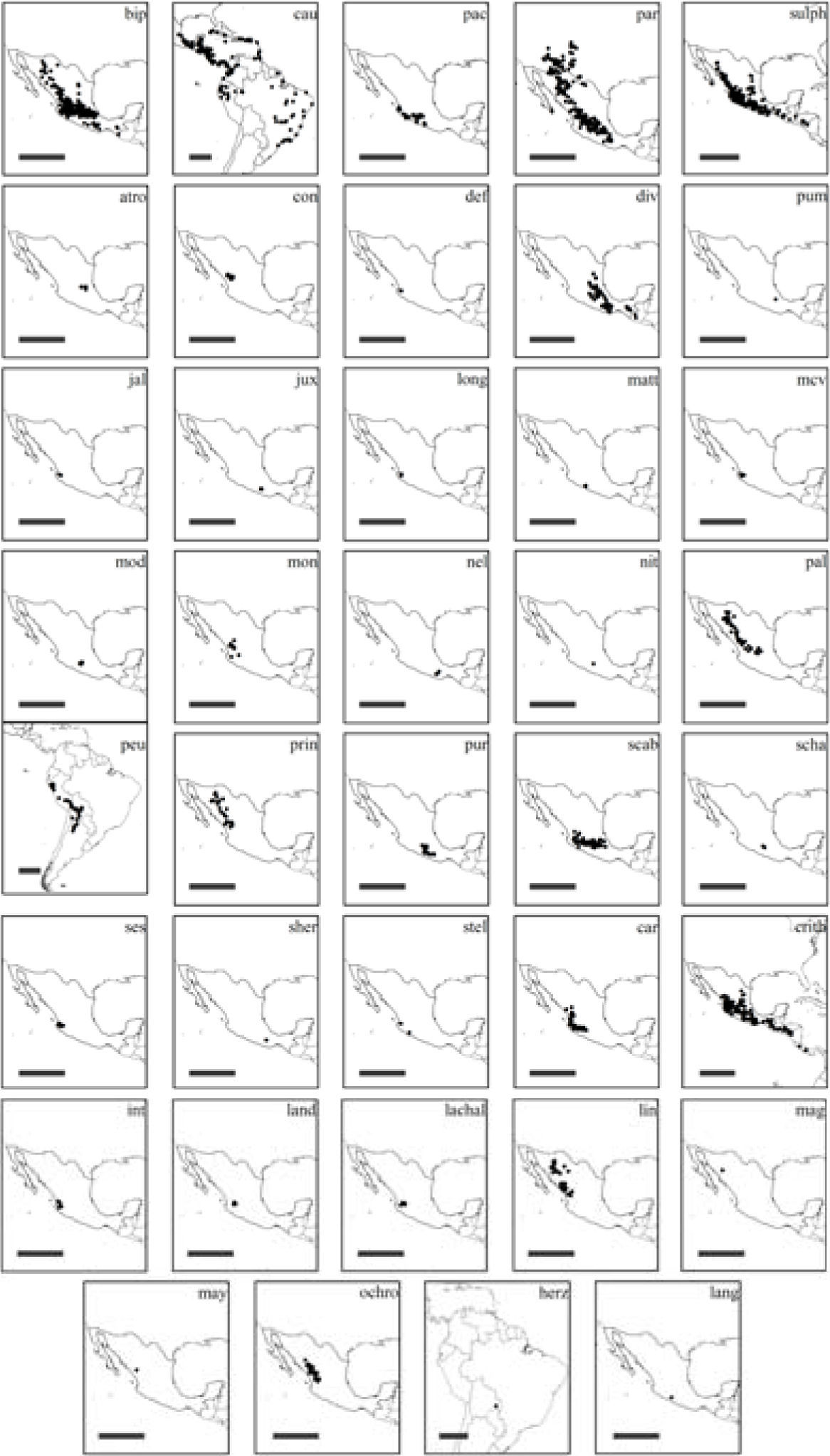
Cosmos bipinnatus Cav., C. caudatus, C. crithmifolius Kunth, C. parviflorus, C. peucedanifolius y C. sulphureus, tuvieron la mayor área de distribución y la máxima distancia entre 2 puntos (Fig. 3, Cuadro 1). Cosmos caudatus tuvo la mayor distancia con un valor de 7 702km. El caso contrario se observó en C. deficiens (Sherff) Melchert, C. juxtlahuacensis, C. longipetiolatus, C. mattfeldii, C. schaffneri Sherff y C. sherffii, con menos de 50km de distancia entre los puntos más distantes.

Distribución geográfica de los taxa de Cosmos. La barra de escala equivale a 1 000km. Se presentan los nombres de los taxa conforme a las siguientes claves: hierbas anuales: bip= Cosmos bipinnatus, cau= Cosmos caudatus, pac= Cosmos pacificus, par= Cosmos parviflorus, sulph= Cosmos sulphureus. Hierbas perennes: atro= Cosmos atrosanguineus, con= Cosmos concolor, def= Cosmos deficiens, div= Cosmos diversifolius var. diversifolius, pum= Cosmos diversifolius var. pumilus, jal= Cosmos jaliscensis, jux= Cosmos juxtlahuacensis, long= Cosmos longipetiolatus, matt= Cosmos mattfeldii, mcv= Cosmos mcvaughii, mod= Cosmos modestus, mon= Cosmos montanus, nel= Cosmos nelsonii, nit= Cosmos nitidus, pal= Cosmos palmeri, peu= Cosmos peucedanifolius, prin= Cosmos pringlei, pur= Cosmos purpureus, scab= Cosmos scabiosoides, scha= Cosmos schaffneri, ses= Cosmos sessilis, sher= Cosmos sherffii, stel= Cosmos stellatus. Sufrútices: car= Cosmos carvifolius, crith= Cosmos crithmifolius, int= Cosmos intercedens, land= Cosmos landii var. landii, lachal= Cosmos landii var. achalconensis, lin= Cosmos linearifolius var. linearifolius, mag= Cosmos linearifolius var. magnifolius, may= Cosmos linearifolius var. maysilessi, ochro= Cosmos ochroleucoflorus. Hábito de crecimiento no determinado: herz= Cosmos herzogii, lang= Cosmos langlassei.
Los valores de EOO y AOO mostraron que Cosmos agrupa especies con una gran amplitud geográfica (Fig. 3, Cuadro 1). Cosmos sherffii es la especie de menor EOO, con 3km2 y C. caudatus es la de mayor extensión, con 13 108 702km2. Son 16 taxa los que tienen un valor de EOO mayor a 20 000km2: Cosmos bipinnatus, C. carvifolius Benth., C. caudatus, C. crithmifolius, C. diversifolius Otto ex Knowles et Westc. var. diversifolius, C. linearifolius (Sch. Bip.) Hemsl. var. linearifolius, C. montanus Sherff, C. ochroleucoflorus Melchert, C. pacificus, C. palmeri B. L. Rob., C. parviflorus, C. peucedanifolius, C. pringlei B. L. Rob. et Fernald, C. purpureus (DC.) Benth. et Hook. f. ex Hemsl., C. scabiosoides Kunth y C. sulphureus. Los taxa con EOO menores a 20 000km2 y mayores a 5 000km2 son: C. concolor Sherff y C. intercedens Sherff; mientras 12 taxa, C. atrosanguineus (Ortgies) Hemsl., C. jaliscensis, C. juxtlahuacensis, C. landii Sherff var. landii, C. landii Sherff var. achalconensis Melchert, C. mattfeldii, C. mcvaughii Sherff, C. modestus Sherff, C nelsonii, C. schaffneri, C. sessilis Sherff y C. stellatus Sherff, tienen intervalos geográficos entre 100 y 5 000km2. Finalmente, los 9 taxa que no sobrepasan los 100km2 en su EOO (Cuadro 1) son: Cosmos deficiens, C. diversifolius var. pumilus, C. herzogii, C. langlassei Sherff, C. linearifolius var. magnifolius, C. linearifolius var. maysilessi, C. longipetiolatus, C. nitidus y C. sherffii.
En Cosmos, los valores de AOO variaron de 12km2 en C. mattfeldii y C. sherffii a 3 048km2 en C. sulphureus. De hecho, este es el único taxón con más de 2 000km2. Los taxones que no superan este límite, pero tienen áreas de más de 500km2 son 5, C. bipinnatus, C. caudatus, C. crithmifolius, C. diversifolius var. diversifolius y C. parviflorus. La mayoría de los taxa analizados tienen AOO entre los 10 y 500km2. Por último, con menos de 10km2 se encontró a C. diversifolius var. pumilus, C. herzogii, C. langlassei, C. linearifolius var. magnifolius, C. linearifolius var. maysilessi y C. nitidus.
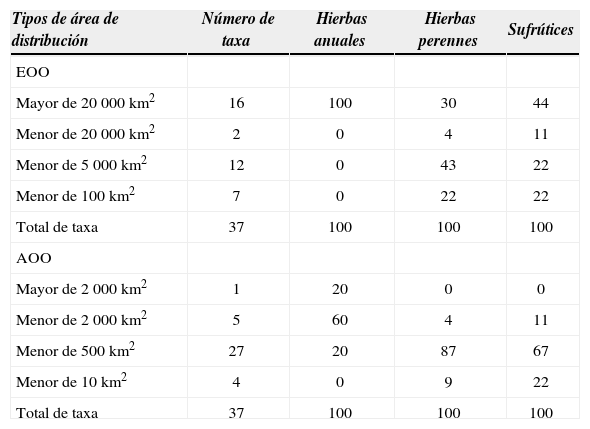
En general, los valores de EOO y AOO revelaron la existencia de 3 patrones en las áreas de distribución, que corresponden con los hábitos de crecimiento de Cosmos (Fig. 3). Las hierbas anuales y la mayoría de los sufrútices (44%) tuvieron una EOO de más de 20 000km2 (Cuadro 2). La mayoría de las hierbas perennes (43%) tuvieron áreas de distribución entre 100 y 5 000km2 y casi la mitad de ellas fueron endémicas del occidente de México. Las hierbas perennes que tienen rangos geográficos mayores a 20 000km2 (30%) fueron: Cosmos diversifolius var. diversifolius, C. montanus, C. palmeri, C. peucedanifolius, C. pringlei, C. purpureus y C. scabiosoides y únicamente C. concolor (4%) presentó valores entre los 5 000 y 20 000km2 y otros 5 taxa (22%) tuvieron una distribución muy restringida, es decir, menor a 100km2 (Cuadros 1 y 2).Cosmos intercedens Sherff fue la única especie sufruticosa (11%) con un rango geográfico entre los 5 000 y 20 000km2 y en el resto se observaron áreas menores a 5 000km2.
Porcentaje de las formas de crecimiento que tienen un valor de EOO (área estimada de presencia de un taxón) y AOO (el área de ocupación del taxón) por categorías de área que se utilizan para evaluar la ubicación de un taxón en alguna categoría de riesgo según la IUCN
| Tipos de área de distribución | Número de taxa | Hierbas anuales | Hierbas perennes | Sufrútices |
|---|---|---|---|---|
| EOO | ||||
| Mayor de 20 000km2 | 16 | 100 | 30 | 44 |
| Menor de 20 000km2 | 2 | 0 | 4 | 11 |
| Menor de 5 000km2 | 12 | 0 | 43 | 22 |
| Menor de 100km2 | 7 | 0 | 22 | 22 |
| Total de taxa | 37 | 100 | 100 | 100 |
| AOO | ||||
| Mayor de 2 000km2 | 1 | 20 | 0 | 0 |
| Menor de 2 000km2 | 5 | 60 | 4 | 11 |
| Menor de 500km2 | 27 | 20 | 87 | 67 |
| Menor de 10km2 | 4 | 0 | 9 | 22 |
| Total de taxa | 37 | 100 | 100 | 100 |
Los patrones de AOO son similares a los mostrados por EOO (Cuadro 2). De acuerdo con este valor, la mayoría de las especies anuales (60%) tienen áreas muy amplias, e incluso C. sulphureus rebasa los 2 000km2. Cosmospacificus, la hierba anual con menor área de distribución, tiene un área geográfica entre los 10 y 500km2. En este ámbito de variación se encuentran la mayoría de las hierbas perennes (87%) y los sufrútices (67%). Cosmos diversifolius var. diversifolius, una hierba perenne y C. crithmifolius, una planta sufruticosa, tienen AOO tan amplias como las de las hierbas anuales. Al igual que su EOO, C. intercedens fue el único sufrútice (11%) con superficies de ocupación entre los 500 y 2 000km2. Los sufrútices C. linearifolius var. magnifolius y C. linearifolius var. maysilessi y las hierbas perennes C. diversifolius var. pumilus y C. nitidus, restringidos a su localidad tipo, no se extienden a más de 10km2 (Cuadro 2).
Riqueza de especies. El análisis de riqueza confirmó que México es el país que tiene la mayor riqueza de Cosmos. En su territorio crecen 37 taxa, incluyendo 33 especies y 4 variedades. Sólo Cosmos peucedanifolius y C. herzogii no se encuentran en el país. Guatemala fue el segundo lugar con 4 taxones (Cuadro 3): C. caudatus, C. crithmifolius, C. diversifolius var. diversifolius y C. sulphureus.
Países con sus respectivos departamentos, estados o provincias de América que acumularon más de 2 taxa. Los nombres de los taxa corresponden a las claves de la Fig. 3. Las localidades en cada país se enlistan con base en el número de taxones
| Localidad (taxa) | Abreviatura del taxón |
|---|---|
| BOLIVIA (2) | |
| Santa Cruz (2) | herz, peu |
| COSTA RICA (2) | |
| San José (2) | cau, crith |
| GUATEMALA (4) | |
| Chimaltenango (2) | crith, div |
| Chiquimula (2) | cau, crith |
| Huehuetenango (2) | crith, sulph |
| Izabal (2) | cau, sulph |
| HONDURAS (3) | |
| Francisco Morazán (3) | cau, crith, sulph |
| Comayagua (2) | cau, crith |
| Cortés (2) | cau, crith |
| MÉXICO (37) | |
| Jalisco (19) | bip, car, crith, def, int, jal, land, lachal, long, mcv, mon, ochro, pac, pal, par, scab, ses, stel, sulph |
| Durango (12) | bip, car, con, crith, lin, may, mon, ochro, pal, par, prin, sulph |
| Estado de México (11) | bip, crith, div, mod, nit, pac, par, pur, scab, scha, sulph |
| Oaxaca (10) | bip, cau, crith, div, jux, nel, par, pur, sher, sulph |
| Michoacán (9) | bip, car, crith, div, pac, par, scab, stel, sulph |
| Nayarit (9) | bip, car, crith, int, lin, mon, ochro, par, sulph |
| Sinaloa (8) | bip, car, crith, lin, ochro, par, prin, sulph |
| Veracruz (8) | bip, cau, crith, div, pum, par, scab, sulph |
| Zacatecas (8) | bip, crith, mon, ochro, pal, par, scab, sulph |
| Chihuahua (7) | bip, lin, mag, ochro, pal, par, prin |
| Guanajuato (7) | atro, bip, crith, div, pal, par, sulph |
| Morelos (7) | bip, crith, div, par, pur, scab, sulph |
| San Luis Potosí | (7) bip, cau, crith, div, pal, par, sulph |
| Sonora (6) | bip, lin, pal, par, prin, sulph |
| Hidalgo (6) | atro, bip, crith, div, par, scab |
| Puebla (6) | bip, crith, div, par, scab, sulph |
| Querétaro (6) | atro, bip, crith, div, par, sulph |
| Aguascalientes (5) | bip, crith, pal, par, sulph |
| Chiapas (5) | bip, cau, crith, div, sulph |
| Distrito Federal (5) | bip, crith, div, par, scab |
| Nuevo León (4) | bip, crith, div, sulph |
| Tlaxcala (4) | bip, div, scab, par |
| Baja California Sur (2) | par, sulph |
| Coahuila (2) | bip, par |
| Colima (2) | pac, sulph |
| Tamaulipas (2) | crith, div |
| NICARAGUA (3) | |
| Estelí (3) | cau, crith, sulph |
| Nueva Segovia (2) | cau, crith |
| PERÚ (2) | |
| Amazonas (2) | cau, peu |
Cosmos tiene una amplia distribución en México. Se encuentra en todos los estados, excepto Baja California y Yucatán. El estado con la mayor riqueza es Jalisco, con 19 taxa, seguido por Durango (12), el Estado de México y Guerrero, cada uno con 11 (Cuadro 3). El resto de los estados tienen menos de 11 taxa (Cuadro 3) y en Campeche, Quintana Roo y Tabasco se registró una sola especie.
En los Estados Unidos de Norteamérica se registró solamente a C. parviflorus y C. caudatus es el único representante del grupo en los países de la cuenca del Mar Caribe. Las especies de Cosmos prosperan en todos los países de América Central. En Costa Rica, Guatemala, Honduras y Nicaragua se registran más de una especie (Cuadro 3). En la provincia de San José, Costa Rica, crecen C. caudatus y C. crithmifolius. En Guatemala prosperan C. caudatus, C. crithmifolius, C. diversifolius var. diversifolius y C. sulphureus. Estos taxa están repartidos en los departamentos de Chimaltenango, Chiquimula, Huehuetenango e Izabal (Cuadro 3). En Honduras, en los departamentos de Francisco Morazán, Comayagua y Cortés, están registrados C. caudatus, C. crithmifolius y C. sulphureus, siendo Francisco Morazán el único con las 3 especies. En Nicaragua habitan C. caudatus, C. crithmifolius y C. sulphureus en los departamentos de Estelí y Nueva Segovia (Cuadro 3).
En Sudamérica, la distribución de Cosmos es casi general. Cosmos caudatus se registró de Brasil, Colombia, Ecuador, Guyana, Surinam y Venezuela. Cosmos herzogii y C. peucedanifolius crecen en el departamento de Santa Cruz, Bolivia y C. caudatus y C. peucedanifolius habitan en el departamento de Amazonas, Perú. En Argentina sólo se ha documentado la presencia de C. peucedanifolius. En Chile, Paraguay, Guyana Francesa y Uruguay no se encontraron registros para este género.
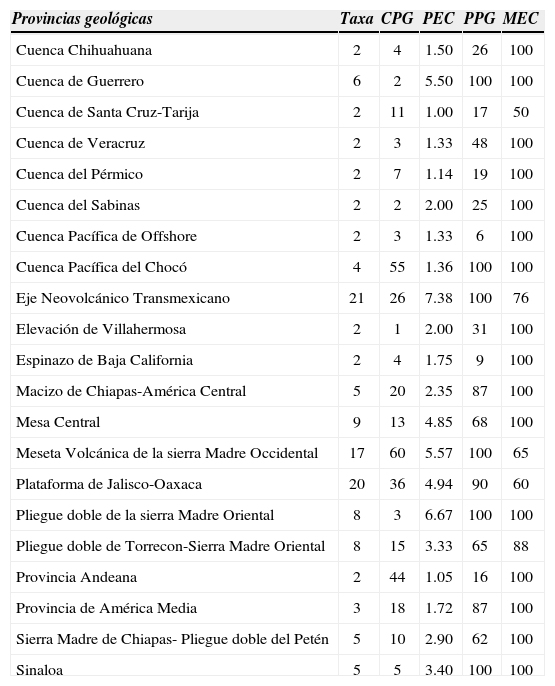
Cosmos está representado en 66 de las 257 provincias geológicas de América continental e insular, lo que representa el 25.7% de ellas. Cuarenta y cinco provincias están ocupadas por una sola especie y en 21 hay entre 2 y 21 taxa (Cuadro 4). Las provincias más ricas en especies son el Eje Neovolcánico Transmexicano (21), seguida por la Plataforma de Jalisco-Oaxaca y la Meseta Volcánica de la Sierra Madre Occidental, con 20 y 17 taxa, respectivamente (Cuadro 4). Estas provincias geológicas se encuentran en el territorio mexicano y corresponden a las provincias fisiográficas Eje Neovolcánico, Sierra Madre del Sur y Sierra Madre Occidental, según Inegi (2001).
Estadísticas de riqueza de especies por provincia geológica basadas en la cuadrícula de 80×80km. Sólo se tomaron en cuenta las celdas con más de un registro. Abreviaturas: CPG (número de celdas por provincia geológica), PEC (promedio de especies por celda), PPG (porcentaje de la provincia geológica ocupada por celdas con al menos un registro) y MEC (porcentaje del mayor número de especies en una celda)
| Provincias geológicas | Taxa | CPG | PEC | PPG | MEC |
|---|---|---|---|---|---|
| Cuenca Chihuahuana | 2 | 4 | 1.50 | 26 | 100 |
| Cuenca de Guerrero | 6 | 2 | 5.50 | 100 | 100 |
| Cuenca de Santa Cruz-Tarija | 2 | 11 | 1.00 | 17 | 50 |
| Cuenca de Veracruz | 2 | 3 | 1.33 | 48 | 100 |
| Cuenca del Pérmico | 2 | 7 | 1.14 | 19 | 100 |
| Cuenca del Sabinas | 2 | 2 | 2.00 | 25 | 100 |
| Cuenca Pacífica de Offshore | 2 | 3 | 1.33 | 6 | 100 |
| Cuenca Pacífica del Chocó | 4 | 55 | 1.36 | 100 | 100 |
| Eje Neovolcánico Transmexicano | 21 | 26 | 7.38 | 100 | 76 |
| Elevación de Villahermosa | 2 | 1 | 2.00 | 31 | 100 |
| Espinazo de Baja California | 2 | 4 | 1.75 | 9 | 100 |
| Macizo de Chiapas-América Central | 5 | 20 | 2.35 | 87 | 100 |
| Mesa Central | 9 | 13 | 4.85 | 68 | 100 |
| Meseta Volcánica de la sierra Madre Occidental | 17 | 60 | 5.57 | 100 | 65 |
| Plataforma de Jalisco-Oaxaca | 20 | 36 | 4.94 | 90 | 60 |
| Pliegue doble de la sierra Madre Oriental | 8 | 3 | 6.67 | 100 | 100 |
| Pliegue doble de Torrecon-Sierra Madre Oriental | 8 | 15 | 3.33 | 65 | 88 |
| Provincia Andeana | 2 | 44 | 1.05 | 16 | 100 |
| Provincia de América Media | 3 | 18 | 1.72 | 87 | 100 |
| Sierra Madre de Chiapas- Pliegue doble del Petén | 5 | 10 | 2.90 | 62 | 100 |
| Sinaloa | 5 | 5 | 3.40 | 100 | 100 |
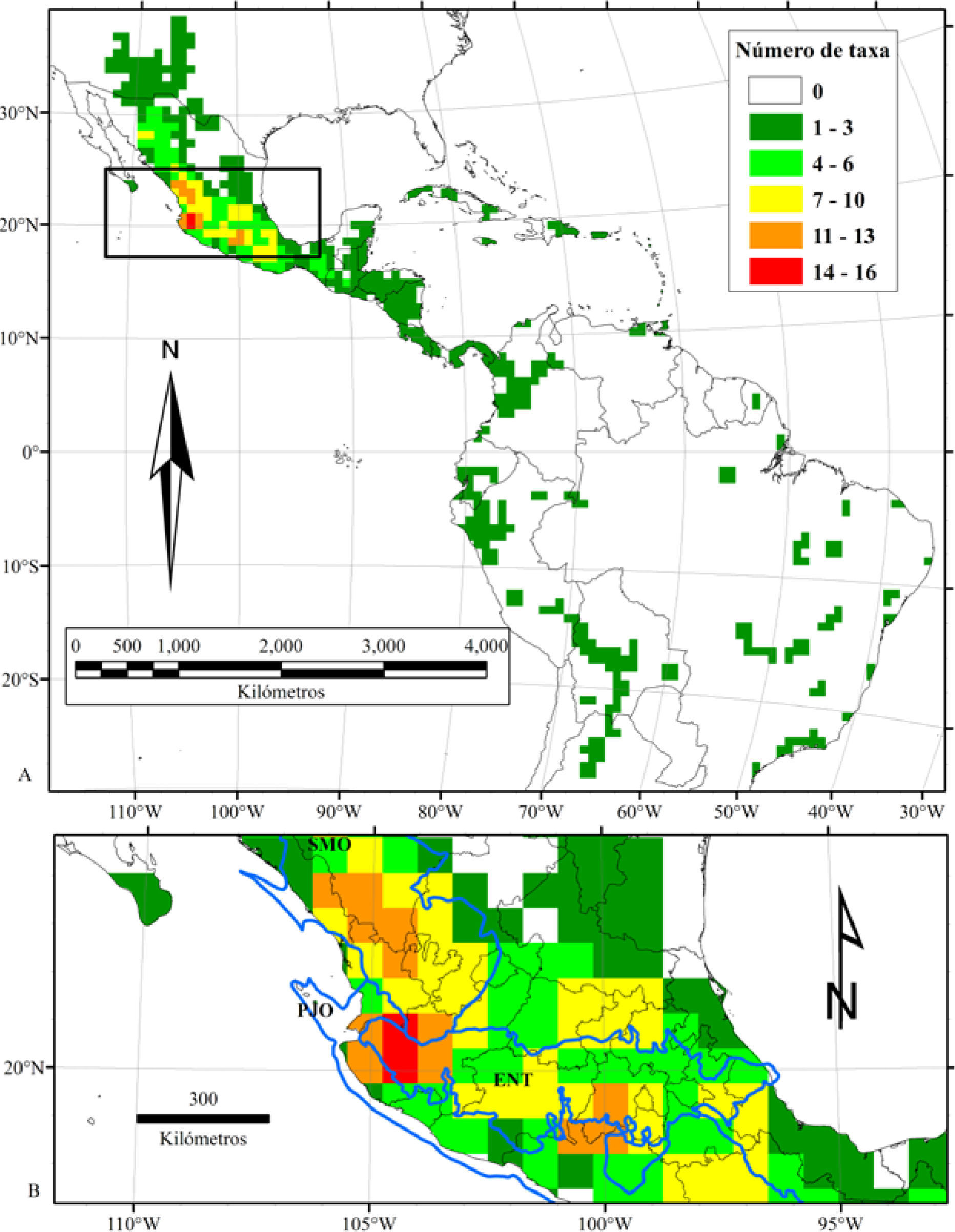
El análisis de la riqueza de Cosmos, basado en la cuadrícula de 80×80km, permitió ubicar con más detalle las regiones más ricas. De esta manera, fue posible identificar 14 celdas agrupadas en 3 áreas aisladas como sitios de alta riqueza (11 o más taxa), todas localizadas en México. La primera se encuentra en el occidente de Jalisco, la segunda en el Estado de México y la tercera en el sur de Durango (Fig. 4). El occidente de Jalisco está integrado por 6 celdas que contienen a 17 especies, es decir, 45.9% de la diversidad de Cosmos. El área se localiza en la confluencia de las 3 provincias geológicas más diversas, el Eje Volcánico Transmexicano, la Plataforma de Jalisco-Oaxaca y la Meseta Volcánica de la Sierra Madre Occidental (Fig. 4).
, riqueza de Cosmos por cuadrícula de 80×80km. (B), detalle los 3 sitios de mayor riqueza del género y las provincias geológicas más diversas en México. (SMO), Meseta Volcánica de la Sierra Madre Occidental; (ENT), Eje Neovolcánico Transmexicano; (PJO), Plataforma de Jalisco-Oaxaca.")
(A), riqueza de Cosmos por cuadrícula de 80×80km. (B), detalle los 3 sitios de mayor riqueza del género y las provincias geológicas más diversas en México. (SMO), Meseta Volcánica de la Sierra Madre Occidental; (ENT), Eje Neovolcánico Transmexicano; (PJO), Plataforma de Jalisco-Oaxaca.
A la mitad del Eje Volcánico Transmexicano y en sus límites con la Plataforma de Jalisco-Oaxaca y la Cuenca de Guerrero, se encontró la segunda área de alta riqueza. Se compone de 3 celdas que incluyen el Nevado de Toluca y se extiende hacia el sur hasta la Plataforma de Jalisco- Oaxaca. La zona está localizada entre el Estado de México, Guerrero y Michoacán, donde coinciden 12 especies, que representan 32.4% de la diversidad de Cosmos (Fig. 4). Por último, en una zona que agrupa 5 celdas en el sur de Durango, donde colindan los estados de Sinaloa, Nayarit, Jalisco y Zacatecas, es posible encontrar a 11 taxa, es decir, 29.7% de la diversidad del género. Esta área se localiza en el extremo sur de la Meseta Volcánica de la Sierra Madre Occidental (Fig. 4).
Cada una de las 3 áreas está bien definida y contiene tanto especies endémicas como compartidas. En el occidente de Jalisco se localizan 17 taxa, los cuales son C. bipinnatus, C. carvifolius, C. crithmifolius, C. deficiens, C. intercedens, C. jaliscensis, C. landii var. landii, C. landii var. achalconensis, C. longipetiolatus, C. mcvaughii, C. montanus, C. pacificus, C. parviflorus, C. scabiosoides, C. sessilis, C. stellatus y C. sulphureus. A sus vez, en la región del sur de Durango se localizaron 11 taxa, que son C. bipinnatus, C. carvifolius, C. concolor, C. crithmifolius, C. linearifolius var. linearifolius, C. montanus, C. ochroleucoflorus, C. palmeri, C. parviflorus, C. pringlei y C. sulphureus. Entre ambas, existe una distancia de 50km y comparten 6 especies. En el área del Estado de México ocurren 12 taxa, C. bipinnatus, C. crithmifolius, C. diversifolius var. diversifolius, C. langlassei, C. modestus, C. nitidus, C. pacificus, C. parviflorus, C. purpureus, C. scabiosoides, C. schaffneri y C. sulphureus. Esta área comparte con el occidente de Jalisco 6 especies y están separadas por aproximadamente 250km. Entre el sur de Durango y el Estado de México existen 470km de distancia y comparten a C. bipinnatus, C. crithmifolius, C. parviflorus y C. sulphureus.
El análisis de riqueza por cuadrícula y las estadísticas por provincia geológica basadas en la cuadrícula de 80×80km, revelaron que la diversidad en Cosmos está distribuida de manera heterogénea (Fig. 4, Cuadro 4). Las estadísticas indican que el número de celdas con 1 o más registros varió de 1 a 60 celdas por provincia. La Cuenca de Pernambuco, la Cuenca de Santos, la Cuenca del Chicontepec, la Cuenca del Espíritu Santo, la Elevación de Villahermosa y la Sierra Nevada de Santa Marta tuvieron una celda con al menos un registro. En el otro extremo, la Meseta Volcánica de la Sierra Madre Occidental presentó 60 celdas con registros (Cuadro 4). De las provincias que tuvieron sólo una celda con una o más observaciones, esa única celda abarcó desde el 2 hasta el 64% de su territorio y las provincias que fueron cubiertas en un 100% por celdas que tuvieron al menos un registro, tuvieron de 2 a 60 celdas (Cuadro 4). La Meseta Volcánica de la Sierra Madre Occidental fue la provincia con mayor número de celdas con registros, seguido de la Cuenca Pacífica de Chocó y la Andeana (Cuadro 4). La Meseta Volcánica de la Sierra Madre Occidental, la Plataforma de Jalisco-Oaxaca y el Eje Volcánico Transmexicano tuvieron el mayor número de especies en una sola celda. En estas últimas 3 provincias es evidente la repartición desigual de la riqueza, pues en ninguna de las celdas de mayor riqueza se encontró el 100% de las especies de la provincia (Cuadro 4, Fig. 4). En cambio, en la mayoría de las provincias, la totalidad de su diversidad específica se incluyó en una sola celda.
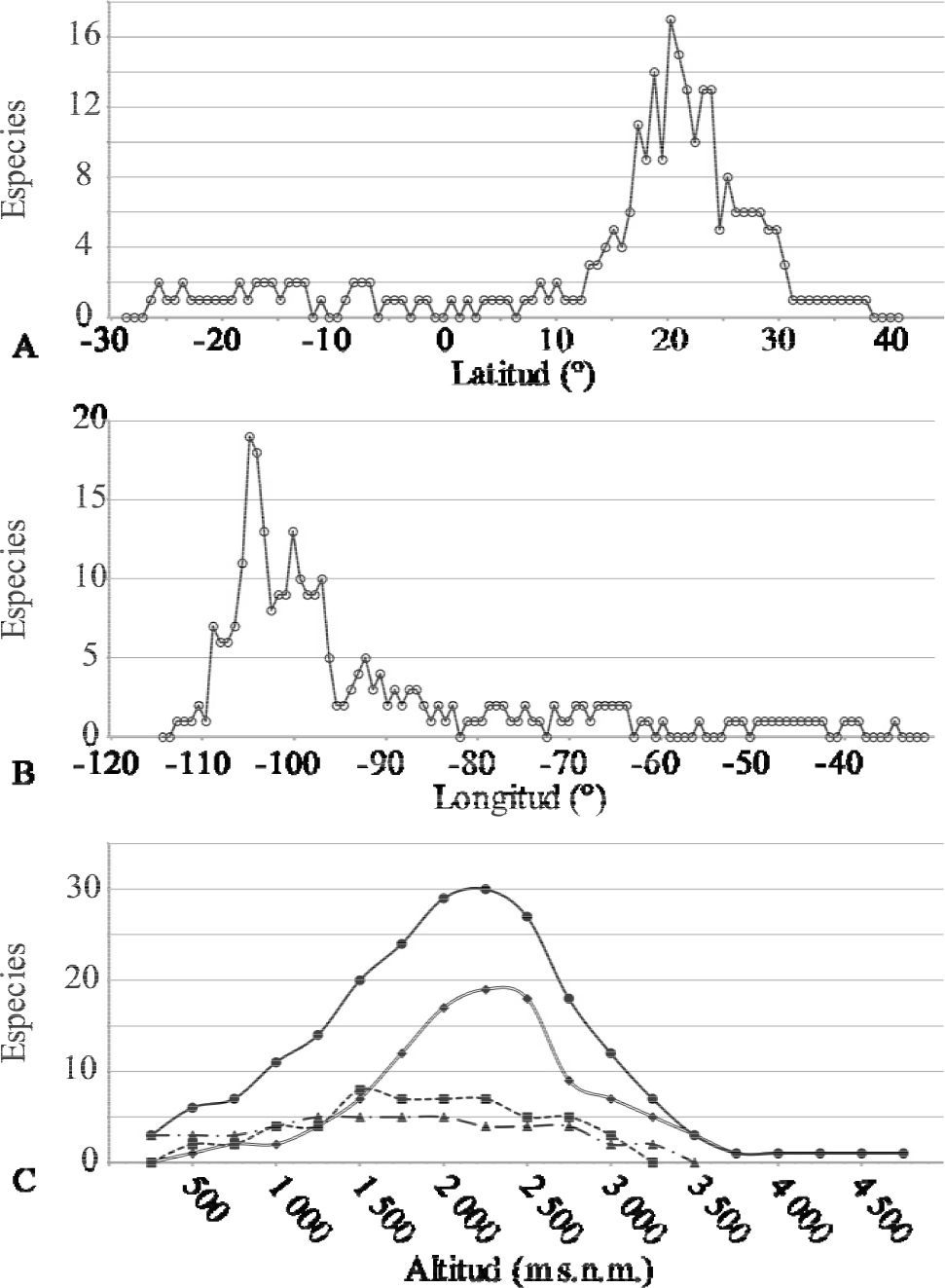
El mayor número de taxa se concentra entre los 16 y 23° N (Fig. 5A), con su máxima riqueza entre los 20 y 21.4° N. Para la longitud, se observa una riqueza máxima entre los 103.7 y 105.2° O y otro, menos diverso, entre los 99 y 101° O (Fig. 5B). En cuanto a la altitud, la mayor cantidad de taxa se localiza entre los 1 750 y 2 500m. Las especies anuales y los sufrútices se concentraron en altitudes entre los 500 y 2 500m, mientras que la mayoría de las hierbas perennes se localizaron en altitudes que superan los 2 000m (Fig. 5C).
, longitud (B) y altitud (C). La línea continua con círculos, representa a todos los taxa; la línea doble con rombos, las hierbas perennes; línea intermitente con cuadros, los sufrútices y la línea intermitente con triángulos, las hierbas anuales.")
Riqueza de especies del género Cosmos, con base en latitud (A), longitud (B) y altitud (C). La línea continua con círculos, representa a todos los taxa; la línea doble con rombos, las hierbas perennes; línea intermitente con cuadros, los sufrútices y la línea intermitente con triángulos, las hierbas anuales.
Base de datos. Los datos de los ejemplares de herbario y su análisis mediante un SIG permitieron la determinación de los patrones de distribución y la detección de áreas con mayor riqueza de Cosmos. Villaseñor (1991), al evaluar las especies endémicas de Heliantheae, analizó 4 de los 33 estados de México. En el presente trabajo fue posible estudiar al género en toda su área de distribución. Por otra parte, como ya había sido indicado por Paterson et al. (2000), el uso de la información resguardada en las colecciones biológicas representó una ventaja por la disponibilidad y rapidez en el manejo de gran cantidad de información a un costo razonable.
Área de distribución. El cálculo de EOO y AOO fue útil para determinar las especies con rangos geográficos amplios y restringidos. Aunque EOO y AOO se emplean para calcular las áreas de distribución, cada una contribuye con información diferente (Callmander et al., 2007) y analizadas en conjunto, resultan complementarias. EOO y AOO no siempre varían en la misma magnitud; en otras palabras, un taxón puede tener una EOO amplia o restringida en comparación a otras especies, pero no sería el caso para su AOO. Un ejemplo es lo que ocurre con C. caudatus y C. sulphureus. Ambas especies están ampliamente distribuidas, pero el valor de EOO de la primera tiene un polígono 10 veces mayor al de la segunda. En consecuencia, C. caudatus es la especie más ampliamente distribuida. Sin embargo, C. sulphureus supera casi 3 veces el AOO de C. caudatus, lo que la convierte en la especie con mayor área de distribución. El análisis de estas 2 fuentes de información indica que C. caudatus tiene la mayor distancia entre los 2 puntos más lejanos, pero C. sulphureus tiene más registros espacialmente distribuidos. Es decir, los registros de C. caudatus generan un polígono muy amplio, lo que tiene como consecuencia un valor mayor de EOO y la distribución de C. sulphureus genera muchas celdas de ocurrencia, lo que produce un valor mayor de AOO.
Con base en el valor de EOO, C. caudatus tiene la distribución geográfica más amplia. Woodson et al. (1975) especularon que esta especie es nativa de Centroamérica y Las Antillas y su presencia en Sudamérica es el resultado de introducciones recientes, por su valor ornamental. Posiblemente por esta razón, la especie ha sido colectada en sitios cercanos a la influencia humana y los sitios de colecta se encuentren alejados entre sí. Su distribución en Sudamérica es muy dispersa, lo que la coloca entre las especies de Cosmos de mayor distribución. Una explicación alternativa es la falta de registros entre localidades distantes como consecuencia de falta de colecta.
Otro caso semejante, pero con especies con una distribución restringida, lo presentan Cosmos deficiens y C. longipetiolatus. La primera de éstas tiene el doble de registros que la segunda y, sin embargo, el valor de su EOO es menos de la mitad que el mismo valor para C. longipetiolatus. Por el contrario, el valor de AOO es mayor en C. deficiens que en C. longipetiolatus. Esto se explica porque los registros de C. longipetiolatus están muy dispersos y producen una EOO muy amplia. Para el cálculo de EOO se ha recomendado la exclusión de disyunciones y discontinuidades dentro del rango del taxón cuando incluya áreas de establecimiento inadecuadas que son obvias (IUCN, 2001). Aplicando este criterio y sólo considerando la parte del polígono entre 1 900 y 2 000m snm (datos no mostrados), la EOO de C. longipetiolatus fue menor que la de C. deficiens. Este criterio fue empleado por Callmander et al. (2007) para eliminar del territorio de la EOO las áreas de mar y las zonas con altitudes menores a 800m, sitios improbables para encontrar especies de Pandanaceae. Su discusión al respecto fue que pueden existir sesgos importantes cuando se cuenta con pocos registros. Willis et al. (2003) rechazan este criterio de exclusión debido a la incertidumbre en la ausencia de la especie. Sin embargo, la exploración exhaustiva de las regiones adyacentes puede revelar la presencia o ausencia del taxón. En tanto no se explore el sitio, el tamaño del área de distribución más apropiado para las especies con grandes superficies sin registro es el AOO (Willis et al., 2003).
Los valores de EOO y AOO también son una pauta para la evaluación de la amenaza de las especies de Cosmos, la asignación de alguna categoría de riesgo y su posterior inclusión en la lista de especies amenazadas de la IUCN (IUCN Standards and Petitions Subcommittee, 2010). Según el criterio “tamaño de rango geográfico y fragmentación, declinación o fluctuaciones”, todas las especies anuales y la mayoría de los sufrútices estarían afuera de la lista roja de las especies amenazadas de la IUCN por tener una EOO de más de 20 000km2 (Cuadro 2). Sin embargo, utilizando AOO, con excepción de C. sulphureus, todos los Cosmos son propensos a estar en alguna categoría de riesgo de la IUCN (IUCN Standards and Petitions Subcommittee, 2010). Consideramos que, mientras no se retire de EOO los sitios donde es evidente que la especie no esté, el área que refleja con mayor realidad la presencia de la especie es AOO.
El análisis comparativo de las áreas de distribución por hábito de crecimiento, mostró diferentes patrones de distribución geográfica en las hierbas anuales, las hierbas perennes y los sufrútices (Fig. 3), ya que las primeras tienen mayores áreas de distribución geográfica. Por el contrario, las hierbas perennes tienen mayor diversidad, pero las áreas de distribución son, en general, pequeñas. Se trata de especies con distribución restringida, con excepción de Cosmos scabiosoides, C. diversifolius var. diversifolius y C. peucedanifolius. Los taxa herbáceos perennes están repartidos en poblaciones pequeñas (la mayoría de menos de 500km2) por todo el continente, desde el norte de Chihuahua, en México, hasta la provincia de Tucumán, en el norte de Argentina. Excepto por C. crithmifolius, los sufrútices crecen principalmente en el occidente de México. Están ubicados a lo largo de la sierra Madre Occidental, en el extremo occidental del Eje Neovolcánico y en el norte de la sierra Madre del Sur.
El análisis de los hábitos de crecimiento de Cosmos a través de una filogenia podría explicar la amplitud de la distribución en las hierbas anuales. El análisis filogenético de Muñiz (2008) basado en secuencias de los espacios internos transcritos del ADN ribosomal (ITS) reveló que el hábito de crecimiento anual es el carácter ancestral. Suponiendo que las hierbas anuales fueron las primeras en diversificarse, también se podría plantear la hipótesis de que éstas han tenido el tiempo suficiente para alcanzar una distribución geográfica más amplia que las hierbas perennes.
Las características de las semillas y los números cromosómicos podrían explicar los patrones de distribución geográfica observados. Rabinowitz (1978) encontró una correlación entre el peso de las semillas de diferentes especies de gramíneas perennes y su abundancia. Sherff (1932) diferenció las secciones de Cosmos por el hábito de crecimiento y la forma de los aquenios. Los aquenios en las secciones exhiben diferencias en la pubescencia y en la forma y longitud del rostro, así como también diferentes patrones de distribución. Un análisis detallado de la morfología de los aquenios y su relación con la distribución geográfica permitirían corroborar esta hipótesis.
Los niveles de ploidía podrían también explicar los patrones de distribución geográfica en las hierbas perennes y los sufrútices de Cosmos. De acuerdo con Melchert (1968, 2010), los sufrútices (Cosmos sección Mesinenia) son diploides o tetraploides (n=11, 22) y crecen en el occidente de México. La excepción es Cosmos crithmifolius, que es hexaploide (n=33), lo cual tiene implicaciones ecológicas y geográficas. Soltis et al. (2003) consideran que un nuevo poliploide puede coexistir con sus progenitores, pero tiene la capacidad de ocupar más sitios. Además, los híbridos suelen tener mayor aptitud en ambientes diferentes a los de sus especies parentales (Grant, 1989; Soltis et al., 2003). Prueba de ello, es que C. crithmifolius abarca los rangos altitudinales de todas las especies sufruticosas, cohabita con casi cualquiera de las especies de Cosmos sección Mesinenia y además se extiende más al sur de los límites geográficos de cualquier Cosmos con hábito sufruticoso. En Cosmos, más de la mitad de los taxa son hierbas perennes (Cosmos sección Discopoda) que habitan áreas de menos de 500km2. La perennidad de las hierbas mediante el desarrollo de rizomas y raíces tuberosas es una adaptación a la vida subterránea, en sitios muy fríos o muy secos, para defenderse de la marcada estacionalidad (Font Quer, 1953). La dominancia de las hierbas perennes de Cosmos entre los 1 750 y 2 500m snm apoya esta idea. Su diversidad morfológica y la gran variedad de hábitats que ocupan es un reflejo de su complejidad cromosómica, pues son las hierbas perennes las que exhiben la mayor diversidad cromosómica (n=12, 13, 24, 36). Incluso, C. diversifolius var. diversifolius y C. scabiosoides, taxa de amplia distribución, presentan en sitios específicos de su distribución geográfica, plantas con el número cromosómico duplicado (Melchert, 1968, 2010). Esta diversidad ecológica, aunada a su variedad cariológica, también se presenta en Chaetanthera Ruiz et Pav., una asterácea sudamericana (Baeza et al., 2009).
Riqueza de especies. El registro fósil y los análisis filogenéticos sugieren que las Asteraceae se originaron en el sur de Sudamérica (Katinas et al., 2009; Tellería et al., 2010). Sin embargo, México es un importante centro de diversificación del grupo, y prueba de ello, es que las tribus Helenieae, Astereae y Heliantheae (escindida en varias tribus y la más diversa de Asteraceae), contienen la mayor cantidad de géneros y especies en México (Turner y Nesom, 1998; Villaseñor, 1991). Para Cosmos, México es el centro de diversificación (Karis y Ryding, 1994) y los resultados del análisis de distribución por latitud y longitud lo comprueban. Este mismo patrón de diversidad se observa en otros géneros como Pinus L. (Styles, 1998), Pedilanthus Neck. ex Poit. (Martínez et al., 2002), Quercus L. (Valencia, 2004) y Dahlia Cav. (Castro-Castro et al., 2012) y familias como Agavaceae (García-Mendoza, 2004), Cactaceae (Hernández et al., 2004) y Crassulaceae (Pérez-Cálix y Franco, 2004), los cuales tienen su mayor riqueza en este país.
La mayor diversidad de Cosmos en México respecto a Sudamérica puede ser la expresión del elemento histórico. La diversidad biológica de México es el resultado de diferentes factores (Toledo, 1994; Rzedowski, 1998), ya que su territorio es complejo desde el punto de vista geológico, fisiográfico y climatológico. En conjunto, forman el escenario ideal para moldear la diversidad biológica de cualquier grupo taxonómico. A nivel mundial, Kreft y Jetz (2007) encontraron que la complejidad topográfica es una causa de diversidad en plantas vasculares. Toledo (1994) mostró que existen variables del medio físico que pueden explicar la distribución de las especies. Si se toma en cuenta que el territorio mexicano posee diversas combinaciones de estas variables, entonces hay una plétora de condiciones disponibles para la diversificación, incluso en áreas geográficas pequeñas. Sin embargo, la cordillera de los Andes, otro sitio de gran complejidad topográfica, está poco representada por el género. Sólo C. herzogii y C. peucedanifolius se registraron en este territorio montañoso y además como especies endémicas. Sin embargo, otros grupos como Solanum L. sección Petota Dumort. y Peperomia subgénero Tildenia (Miq.) Dahlst., presentes en el Eje Neovolcánico, son más ricos en los Andes (Hijmans y Spooner, 2001; Symmank et al., 2011). Ambos géneros están ampliamente distribuidos y en el caso de la sección Petota, existen varios endemismos de distribución muy restringida (Hijmans y Spooner, 2001). En contraste, C. peucedanifolius está presente a lo largo de la cordillera y es muy variable morfológicamente a lo largo de su distribución geográfica y altitudinal (desde los 420m hasta los 4 500m.). Estas variaciones locales han motivado la descripción de nuevas especies y variedades que ahora son consideradas sinónimos de una especie polimórfica. (Hind, 2005). Es posible que las diferencias en el color de las lígulas, la etapa de floración y la amplitud altitudinal sean adaptaciones locales que han conducido a la diversificación de C. peucedanifolius en los Andes.
La diversidad de especies endémicas es otra consecuencia de la topografía accidentada y la compleja historia geológica de México (Rzedowski, 1991, 1998). Ramamoorthy y Elliot (2008) sugirieron que la flora de México es similar a la de una isla por el alto contenido de endemismos. Un área particularmente rica en endemismos es el occidente de Jalisco, donde se conjuntan las provincias fisiográficas Eje Neovolcánico, Sierra Madre del Sur y Sierra Madre Occidental, en un territorio de poco más de 25 000km2. Villaseñor et al. (1998) y Delgadillo et al. (2003) encontraron que el oeste de México es uno de los sitios más diversos en géneros nativos de Asteraceae. Además, Villaseñor (1991) ubicó la mayor cantidad de especies de Heliantheae endémicas en la porción más occidental del Eje Neovolcánico, en Jalisco. Los resultados del análisis de riqueza por cuadrícula y riqueza por latitud y longitud en Cosmos concuerdan con estas observaciones.
El endemismo del occidente de Jalisco no es exclusivo de Cosmos. También se han encontrado especies de pinos (Pérez de la Rosa, 1983, 2009), encinos (González-Villarreal, 2003), agaves (Cházaro-Basáñez et al., 2008), orquídeas (González y Hernández, 2010), salvias (González-Gallegos et al., 2012a, b) y otros géneros de hábitos arbóreos (Villaseñor e Ibarra, 1998). Otro hallazgo relevante en el occidente de Jalisco, es el nodo biogeográfico que localizaron Torres y Luna (2006), mediante un análisis panbiogeográfico con especies de aves, mamíferos, reptiles, helechos y encinos presentes en la Faja Volcánica Transmexicana. Basados en un índice de complementariedad, Torres y Luna (2006) sugieren este nodo como uno de los 4 prioritarios para la conservación. Aunque para el presente trabajo no se obtuvieron índices de complementariedad, es claro que el occidente de Jalisco debe ser considerado un sitio prioritario para la conservación. Cosmos sessilis, C. deficiens, C. jaliscensis, C. longipetiolatus y C. mcvaughii crecen sólo en el occidente de Jalisco y no están compartidas con otras áreas de alta riqueza.
En un contexto similar, varios grupos de celdas de alta diversidad fueron localizados en el sur de Durango (Fig. 4). También se encontraron varios taxa endémicos, como Cosmos concolor, C. linearifolius var. linearifolius, C. linearifolius var. magnifolius, C. linearifolius var. maysilessi, C. ochroleucoflorus y C. pringlei. Resultados similares fueron obtenidos por Villaseñor (1991). Este sitio está ubicado en el sur de la Sierra Madre Occidental, que es una cadena montañosa que funciona como corredor, barrera o refugio de especies endémicas. Por su orientación noroeste-sureste, Bye (1994) sugiere que esta sierra ha funcionado para la migración de las especies templadas hacia el sur y el desplazamiento de la flora tropical hacia el norte.
La zona de alta diversidad del Estado de México se localizó en el centro del Eje Neovolcánico, en el límite con la provincia fisiográfica de la Sierra Madre del Sur (provincias geológicas Plataforma de Jalisco-Oaxaca y Cuenca de Guerrero). Este sitio no fue detectado por Villaseñor (1991) debido a que esta entidad no es de las más ricas en especies endémicas, ya que estuvo representado sólo por las endémicas Cosmos purpureus y C. modestus y otras 6 especies de Heliantheae. Villaseñor y Ortiz (2009) explican que el endemismo en el Eje Neovolcánico es menor al de otras provincias por la migración de sus especies a través de esta cadena montañosa y hacia otras provincias. Por ejemplo, su aparición le permitió funcionar como un puente entre la Sierra Madre Occidental y la Sierra Madre Oriental. Su formación, por un lado dejó nichos disponibles que favorecieron el establecimiento de 2 especies endémicas, de la misma forma que ocurrió para los pinos (Contreras-Medina et al., 2007) y encinos (Valencia, 2009). Pero mientras el endemismo en el área es bajo, la diversidad de especies de Cosmos es alta debido a que su posición geográfica favorece la presencia de especies con diferentes preferencias geográficas, entre ellas C. bipinnatus, C. crithmifolius, C. parviflorus, C. sulphureus, de amplia distribución, C. scabiosoides, característica del Eje Neovolcánico, C. pacificus, endémica de la vertiente occidental y C. diversifolius var. diversifolius, característica de la vertiente oriental. Esta provincia fisiográfica es una región particularmente interesante por su gran diversidad de especies de diferentes linajes vegetales (Rzedowski, 1998; Styles, 1998; Villaseñor et al., 2006).
La presencia de los 3 sitios de alta riqueza en el centro y norte de México concuerda con los patrones encontrados por Rzedowski (1972) respecto a la familia Asteraceae. Además de la complejidad topográfica, México posee en esta zona las preferencias ecológicas de la familia, como son los climas secos y frescos. De esta forma, ésta es la parte de México que podría constituirse como el centro de diversificación del género, ya que abarca las principales zonas montañosas y los climas templados y secos. Además, Rzedowski (1972) encontró que los ecosistemas más ricos en Asteraceae son aquellos que se encuentran en un estado temprano de sucesión. Los suelos del Eje Neovolcánico se derivan principalmente de lavas volcánicas, superficies recién perturbadas, lo que convierte a esta región en el mejor candidato para la diversificación de los miembros de esta familia.
El presente trabajo representa un punto de partida para buscar, con otras herramientas, algunas respuestas para las hipótesis aquí planteadas. Así, con el fin de descifrar la dinámica que han seguido las poblaciones y especies para exhibir los patrones citológicos, ecológicos y geográficos actuales, es necesario recurrir a estudios sobre genética de poblaciones en plantas anuales y perennes, así como comparaciones entre plantas diploides y poliploides. Un análisis geográfico, ejecutado en un contexto filogenético, podría definir el área geográfica basal a partir de la cual Cosmos ocupó gran parte del continente americano y en qué sitios se originaron los diferentes grupos. Finalmente, la filogenia podría revelar si las hierbas perennes comparten un origen común o si son el resultado de adaptaciones independientes a los nuevos ambientes de las “islas” del Eje Neovolcánico. Con base en la filogenia, se podría descifrar la trayectoria evolutiva de los hábitos de crecimiento y números cromosómicos.
Los autores agradecen a Conacyt por el financiamiento de este trabajo, mediante el proyecto 80200 “Filogenia, evolución y potencial ornamental del género Cosmos (Asteraceae)” y las becas 228901 y 229039 de la primera y segundo autor, respectivamente. A los encargados de los herbarios visitados y a todo el personal que nos brindó su apoyo. Se reconoce la ayuda de Conabio, Conafor e Inegi en México y de USGS en Estados Unidos de América por poner a la disposición la cartografía digital. A las instituciones educativas nacionales y extranjeras se les agradece el acceso a las bases de datos electrónicas a través de su personal, principalmente a Tom Wendt y a Amber Schoneman del herbario de la Universidad de Texas. Se reconoce la asesoría de María Elena Vargas Amado en la elaboración de los mapas. Los autores desean expresar su agradecimiento al editor asociado Dr. Guillermo Ibarra Manríquez y a los 2 revisores anónimos por sus apreciables sugerencias al manuscrito.