





Las selvas secas son ecosistemas que mantienen una gran diversidad de especies; sin embargo, son los hábitats mayormente amenazados. El objetivo de este estudio fue evaluar la diversidad de reptiles en 2 microcuencas con selva seca de la cuenca hidrológica Grijalva: Copainalá y Bombaná. Se muestrearon 19 sitios: 9 para Copainalá y 10 para Bombaná; en cada sitio se establecieron 2 transectos para el muestreo de reptiles y se utilizó la técnica de registros por encuentro visual. Se registró un total de 36 especies de reptiles, de las cuales en Copainalá fueron 28 y en Bombaná 23 especies, alcanzando el 80% de la riqueza esperada. La diversidad de especies y el patrón de abundancia fue similar en ambas microcuencas, entre las especies más abundantes se encuentran: Basiliscus vittatus, Sceloporus variabilis, Norops unilobatus y Sphaerodactylus glaucus. La complementariedad de especies entre ambas zonas fue del 58%. Las especies de reptiles utilizaron, principalmente 3 tipos de microhábitat terrestre: hojarasca, sobre roca y suelo. Se registraron 11 especies que se encuentran en algún estatus de conservación. Se sugiere que la región puede funcionar como un corredor biológico de especies de reptiles entre las áreas naturales protegidas adyacentes.
The dry forest is an ecosystem with a relatively high species diversity. It is also one of the most threatened habitats. The objective of this study was to evaluate the reptile diversity in 2 microbasins of dry forest from the Grijalva hydrological basin: Copainalá and Bombaná. We sampled 19 sites, 9 for Copainalá and 10 for Bombaná. At each site we laid out 2 transects along which we registered species of reptiles. In total, we recorded 36 reptile species: 28 in Copainalá and 23 in Bombaná, representing approximately 80% of the expected richness in the area. The species diversity and abundance patterns were similar in both microbasins. Among the more abundant species there were: Basiliscus vittatus, Sceloporus variabilis, Norops unilobatus and Sphaerodactylus glaucus. The complementarity of species between both zones was 58%. The most common microhabitat utilized by reptiles in these areas was terrestrial, including: leaf-litter, on rocks and on the ground. Among the species we registered, 11 present some conservation status. Our results also suggest that this region could function as a biological conservation corridor for reptiles between other near protected natural areas.
Las selvas secas se caracterizan por presentar una larga temporada de sequía, por perder entre 50 y 100% del follaje en época de estiaje y, generalmente se desarrollan por debajo de 1 200m snm (Bezaury, 2010). Están ampliamente distribuidas en México, en la vertiente del Pacífico desde Sinaloa hasta Chiapas, entrando al centro del país a través de la Cuenca del Balsas (Trejo, 2010), lo que les permite albergar cerca de una tercera parte de especies de vertebrados endémicas de México (Ceballos y García, 1995; Ceballos et al., 2010). No obstante, son los ecosistemas neotropicales altamente amenazados (Janzen, 1988).
En los últimos años, la cobertura original de las selvas secas se ha reducido considerablemente, al grado de mantener intacto sólo el 30% de su cobertura nacional (Trejo y Dirzo, 2000). Entre los factores que lo amenazan se encuentra el avance de la frontera agrícola y pecuaria, en especial el sobrepastoreo de ganado vacuno y de cabras (Trejo y Dirzo, 2002). Aunado a esto, existen pocas áreas naturales protegidas que conserven extensiones de selva seca que contengan gran riqueza de especies y endemismos (García, 2006; Ceballos et al., 2010). El WWF (Fondo mundial para la vida silvestre) identificó 7 ecorregiones de selva seca con gran riqueza, endemismo y diversidad y entre éstas se encuentra la depresión central de Chiapas. Dicha identificación busca promover el conocimiento de cada ecorregión y, con ello generar propuestas para su conservación (Ceballos et al., 2010).
La depresión central de Chiapas se caracteriza por tener una cobertura forestal principalmente de selva seca, no obstante, es también una zona altamente deforestada, manteniendo el 8.6% de cobertura original y sólo el 2% se encuentra en buen estado de conservación (García, 2006). Entre las causas por la cuales esta zona ha sido severamente afectada, se encuentra el cambio de uso de suelo, de selva seca a cultivos y/o pastizales (Trejo, 2010). A pesar de la alteración que presenta la zona, se han mostrado altos valores de diversidad herpetofaunística (García et al., 2007; García, 2010), registrándose aproximadamente 85 especies de anfibios y reptiles (García, 2006; García et al., 2007) lo que representa el 26% de la herpetofauna del estado. En Chiapas se han registrado 325 especies de herpetofauna, de las cuales 101 son anfibios y 223 son reptiles (Reynoso et al., 2011).
Además de la gran diversidad biológica que presenta el estado de Chiapas, contiene una amplia red hidrológica, entre las que se encuentra la cuenca del río Grijalva. Dentro de ésta se encuentra la subcuenca hidrológica río Grijalva -Tuxtla Gutiérrez (CNA, 1998), situada en los límites de la depresión central de Chiapas y las montañas del norte. El río Grijalva es el principal afluente que cruza esta región donde desembocan pequeños ríos tributarios. El objetivo del presente trabajo fue evaluar la diversidad de reptiles en 2 microcuencas con selva seca, en las áreas cercanas al cauce del río Grijalva y afluentes que desembocan en él.
Materiales y métodosZona de estudio. La zona que abarca el estudio está ubicada en la cuenca hidrológica río Grijalva. Se delimitaron 2 microcuencas (Fig. 1), entendidas como unidades geográficas caracterizadas por la hidrología, variaciones en la pendiente del terreno, temperatura y precipitación. Para la delimitación y caracterización de las microcuencas se utilizó un modelo digital de elevación generado con un sistema de información geográfica ArcMap/ArcInfo ver 9.2. Se utilizaron coberturas de hidrología superficial, cuerpos de agua y embalses, poblados a escala 1:50000 con coberturas de edafología y vegetación y uso del suelo del 2003 a una escala de 1 250000 (Inegi, 2000). Con base en este estudio de paisaje, se definieron 2 microcuencas: Copainalá con 59 subunidades de paisaje y Bombaná con 71 subunidades, con variaciones en topografía, hidrología y geología. Para más detalles revisar Comisión Federal de Electricidad y Universidad de Ciencias y Artes de Chiapas (2011). En estas 2 microcuencas se ubicaron 19 sitios de muestreo, de los cuales 9 sitios se establecieron en Copainalá y 10 en Bombaná. Estos sitios de muestreo se seleccionaron de manera que abarcaran la mayor diversidad de subunidades de paisaje encontradas en cada microcuenca y siempre a lo largo del río Grijalva y sus afluentes. La microcuenca Copainalá se caracteriza por tener una cobertura forestal de selva seca de 7 445 ha (selva baja caducifolia y subcaducifolia con vegetación secundaria arbustiva), una precipitación pluvial de 1 553mm anuales en el 2009, 20.49° y 34.51° de temperatura mínima y máxima promedio anual, respectivamente (CNA, 2009), pendiente del terreno desde 10 - 45 grados. La microcuenca Bombaná tiene una extensión de 6 657 ha de selva seca (selva baja caducifolia y subcaducifolia con vegetación secundaria), precipitación pluvial de 981.7mm en el 2009, 20.5° y 31.89° de temperatura mínima y máxima promedio anual, respectivamente (CNA, 2009), pendiente del terreno desde áreas de 1 hasta 45 grados. Trabajo de campo. El muestreo se realizó durante el periodo de abril—diciembre del 2009. Con una visita mensual para cada microcuenca, un mes se visitaba Copainalá y el siguiente Bombaná. En cada sitio de muestreo se establecieron 2 transectos con una extensión de 1 000m de largo y 10m de ancho. La duración del muestreo fue de un día para cada sitio, con un esfuerzo de muestreo de 4 personas, 2 personas por cada transecto. En la microcuenca Copainalá, de los 9 sitios de muestreo, 5 fueron visitados 4 veces y los restantes, 6 veces, mientras que en Bombaná todos los 10 sitios fueron visitados 4 veces. La diferencia entre las visitas a los sitios fue para tratar de realizar un esfuerzo de muestreo similar entre las 2 microcuencas. La búsqueda de los organismos se realizó con un tiempo fijo de 8 horas diarias para cada transecto, divididas de la siguiente manera: 3 horas en la mañana, 3 en la tarde y 2 en la noche, utilizando la técnica de registro por encuentros visuales (Heyer et al., 1994). El esfuerzo de muestreo fue de 8hrs/hombre x día, a excepción de 4 sitios muestreados en Copainalá, donde fue de 5hrs/hombre x día, siendo un acumulado de 44 días para Copainalá y 40 días para Bombaná. En estos transectos se realizaron caminatas buscando en los lugares potenciales donde se podrían encontrar estos organismos, tales como: en hojarasca, bajo rocas, bajo troncos y dentro de ellos, sobre ramas de árboles y entre ellas, registrando las especies de reptiles observadas y capturadas (Bruce, 1986). Posteriormente se hizo un análisis del uso de microhábitat. Para la captura de lagartijas se usaron las manos y cañas de pescar, para las serpientes se empleó un gancho herpetológico (Gaviño et al., 1977; Fitzgerald, 2012). Los organismos se capturaron y se identificaron a nivel de especie, después se liberaron en el lugar donde fueron capturados. Cada registro de captura y/o colecta se georreferenció mediante un GPS. Cuando no fue posible la identificación de los organismos en el campo éstos fueron colectados (permiso: SGPA/DGVS08440/09). De las especies listadas en la norma oficial mexicana Nom-059-Semarnat-2010, sólo se tomaron fotografías. Para el sacrificio de los organismos se siguieron las técnicas recomendadas por Pisani-Villa (1974) y la fijación de los organismos fue con formol al 10%. La preservación de los ejemplares fue en alcohol al 70% y éstos se depositaron en la Colección Herpetológica de la Universidad de Ciencias y Artes de Chiapas.

Análisis de los datos. Se calculó la riqueza y diversidad de especies, la primera definida como el número de especies dentro de cada microcuenca y la segunda considerada como el número de especies y su abundancia, empleando el Índice de Shannon-Wiener (H'). Este índice se utilizó debido a que es sensible a las especies raras por considerar la abundancia proporcional de individuos de cada especie (Magurran, 2004). Posteriormente se aplicó la prueba de “t” modificada por Hutcheson para apreciar si existían diferencias significativas entre los índices de diversidad de las microcuencas.
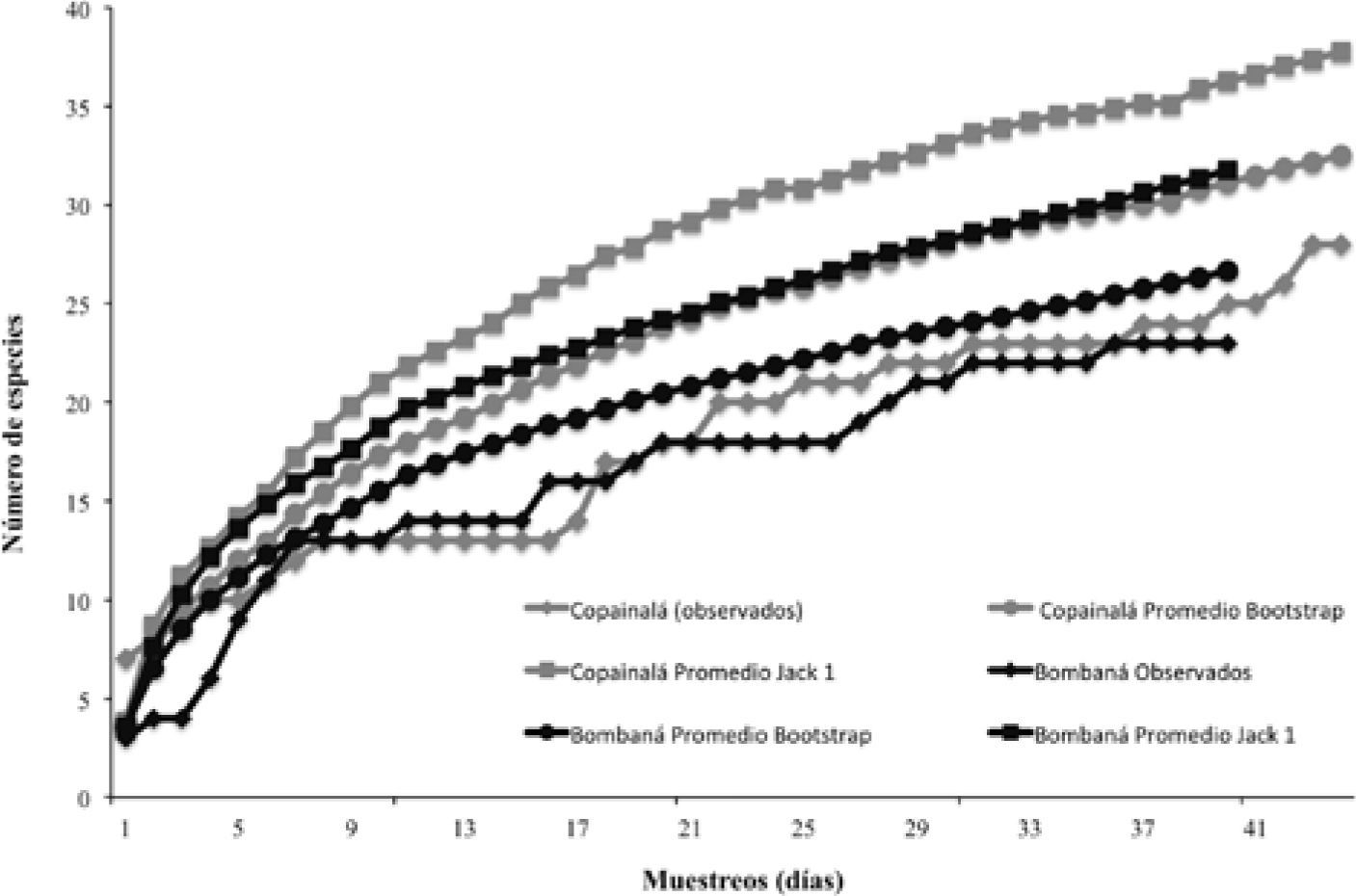
Para evaluar la eficiencia del muestreo y estimar la riqueza de especies esperada en cada microcuenca se utilizaron curvas de acumulación de especies (Soberón y Llorente, 1993; Colwell y Coddington, 1995; Moreno y Halffter, 1999). Estas curvas se generaron para cada microcuenca, utilizando 2 estimadores no paramétricos Bootstrap y Jacknife de primer orden, usando el programa estadístico EstimateS 8.2 (Colwell, 2005). El primer estimador se ha recomendado porque es poco sensible a la presencia de especies raras (Urbina-Cardona y Reynoso, 2005) y por lo tanto, puede ser usado en ensambles que presentan un considerable número de estas especies. El segundo estimador, porque constituye un buen estadístico, dado que sus valores medios tienen una distribución normal (Magurran, 2004), además de que ha sido empleado en algunos estudios de herpetofauna (Carvajal-Cogollo y Urbina-Cardona, 2008; Fernández-Badillo y Goyenechea-Mayer, 2010) y, por lo tanto, los resultados pueden ser comparables. Adicionalmente, el método de Jacknife tiende a dar estimaciones de mayor valor que Boostrap, y consecuentemente podría estar cubriendo un mayor rango de las especies esperadas. Una vez que se obtuvieron los valores esperados de riqueza de cada estimador, se calculó la eficiencia del muestreo por microcuenca expresada en porcentaje, del total de la riqueza registrada respecto a la predicha por los estimadores.
Para los patrones de abundancia, se elaboraron curvas de rango-abundancia (Magurran, 2004) tanto para Copainalá y Bombaná, utilizando la abundancia relativa entendida como el número de individuos registrados por microcuenca. El grado de complementariedad entre las 2 microcuencas fue evaluado mediante un coeficiente de similitud: el índice cuantitativo de Sorensen, el cual asume que todas las especies tienen un peso igual en la ecuación independientemente si son raras o abundantes (Magurran, 2004), es decir, toma en cuenta la abundancia de cada especie en los diferentes tipos de hábitat. Los valores van de cero si las microcuencas son completamente diferentes y no comparten especies, a 1 si hay una completa similitud.
Se describió el uso de microhábitat de las especies de reptiles (Duellman, 1987; Gray-Lovich y Lovich, 2012), considerado como el número de individuos por cada especie encontrados en el sustrato donde fue observado por primera vez, el cual es expresado en porcentaje con respecto al total de microhábitats identificados. Se identificaron 3 categorías de microhábitats con respecto a los hábitos terrestres, arborícolas y acuáticos y, posteriormente, se subdividieron de acuerdo con la gama de sustratos donde se identificaron (Gray-Lovich y Lovich, 2012).
Finalmente, se identificó qué especies se encuentran en alguna categoría de riesgo, de acuerdo con la Nom-059-Semarnat-2010, así como en la lista roja de la Unión Internacional para la Conservación de la Naturaleza (IUCN, 2010).
ResultadosRiqueza, diversidad y composición taxonómica. Se registró un total de 903 reptiles en ambas microcuencas que corresponden a 36 especies (Cuadro 1). Con 2 240 horas de esfuerzo de muestreo acumulado para la microcuenca Copainalá, se registró una mayor riqueza de especies (28), siendo el grupo de lagartijas el que en mayor medida contribuyó a esta riqueza (Cuadro 1), mientras que en Bombaná se aplicó un esfuerzo de muestreo acumulado de 2 560 horas y se registraron 23 especies.
Especies de reptiles registradas en su microhábitat y en ambas microcuencas: Copainalá y Bombaná. El código es una letra o número asignado a cada especie para la figura 3
| Copainalá | Bombaná | Microhábitat | Código | |
|---|---|---|---|---|
| Sauria (lagartijas) | ||||
| Corytophanidae | ||||
| Basiliscus vittatus | X | X | 1, 2, 3, 4, 6, 12, 13, 14, 15 | A |
| Eublepharidae | ||||
| Coleonyx elegans | X | 3 | B | |
| Gekkonidae | ||||
| Hemidactylus frenatus | X | X | 1, 3, 4, 8, 9, 12, 13, 14 | C |
| Phyllodactylus tuberculosus | X | X | 1, 2, 4, 7, 11, 12 | D |
| Sphaerodactylus glaucus | X | X | 1, 2, 3, 4, 5, 6, 7, 8, 12, 13, 14, 15 | E |
| Iguanidae | ||||
| Ctenosauria acanthura | X | X | 2, 3, 13 | F |
| Iguana iguana | X | 3 | G | |
| Phrynosomatidae | ||||
| Sceloporus carinosus | X | X | 1, 3, 5 | H |
| Sceloporus melanorhinus | X | 4 | I | |
| Sceloporus varibilis | X | X | 1, 2, 3, 4, 6, 7, 12, 13, 14 | J |
| Dactyloidae | ||||
| Norops laeviventris | X | 2, 13 | K | |
| Norops unilobatus | X | X | 1, 2, 3, 4, 5, 6, 7, 8, 9 12, 13 | L |
| Norops tropidonotus | X | 2, 3 | M | |
| Mabuyidae | ||||
| Marisora | ||||
| brachypoda | X | X | 1, 2, 3, 4, 5 | N |
| Scincidae | ||||
| Scincella assatus | X | X | 1, 2 | O |
| Teiidae | ||||
| Holcosus undulatus | X | X | 3 | P |
| Aspidocelis depii | X | X | 1, 2, 3 | Q |
| Xantusiidae | ||||
| Lepidophyma chicoasensis | X | 2 | R | |
| Serpentes | ||||
| Boidae | ||||
| Boa constrictor | X | 2, 12 | S | |
| Colubridae | ||||
| Coniophanes piceivittis | X | X | 2, 3, 6 | T |
| Drymobius margaritiferus | X | X | 2, 3, 15 | U |
| Drymarchon melanurum | X | 3, 15 | V | |
| Leptodeira annulata | X | 1, 3, 15 | W | |
| Imantodes cenchoa | X | 2, 6, 13 | X | |
| Masticophis mentovarius | X | 3 | Y | |
| Mastigodryas melanolomus | X | 2 | Z | |
| Salvadora lemniscata | X | X | 1, 2, 3, 15 | 1 |
| Spilotes pullatus | X | 10 | ||
| Trimorphodon biscutatus | X | 13 | 3 | |
| Tropidodipsas fasciatus | X | 3 | 4 | |
| Elapidae | ||||
| Micrurus browni | X | 3 | 5 | |
| Leptotyphlopidae | ||||
| Epictia goudotii | X | 6 | 6 | |
| Loxocemidae | ||||
| Loxocemus bicolor | X | 2 | 7 | |
| Crotalidae | ||||
| Crotalus simus | X | 1, 3 | 8 | |
| Viperidae | ||||
| Atropoides mexicanus | X | 2 | 9 | |
| Testudines | ||||
| Kinosternidae | ||||
| Kinosternon scorpioides | X | 15 | 10 |
Microhábitat: terrestre: 1, sobre roca; 2, hojarasca; 3, sobre suelo; 4, sobre tronco caído; 5, bajo corteza; 6, bajo roca; 7, bajo tronco caído; 8, bromelia; 9, agave; 10, tranca; 11, túnel. Arborícola: 12, en fuste de árbol; 13, en ramas de arbusto; 14, pared. Acuático: 15, orilla de río.
Conjuntando los datos de ambas microcuencas, los estimadores Boostrap y Jacknife 1, predicen entre 41 y 48 especies, respectivamente; es decir, que se registró entre un 75 y un 88% de la riqueza esperada. En la microcuenca Copainalá, éstos mismos estimadores indicaron que se alcanzó entre el 74 y el 86% de las especies esperadas, respectivamente. El primer estimador predice alrededor de 32 especies de reptiles, mientras que el segundo, predice alrededor de 38 potencialmente presentes (Fig. 2). En el caso de Bombaná, los algoritmos Bootstrap y Jacknife 1, predicen 27 y 32 especies de reptiles, respectivamente (Fig. 2), es decir, que se alcanzó con el muestreo realizado, entre 72 y 86% de la riqueza esperada.

Respecto a la diversidad de especies de reptiles encontradas, Copainalá presentó un valor de diversidad de H'= 2.24, mientras que Bombaná tuvo un valor de H'= 2.16. La diferencia entre estos valores no fue significativa (t0.05(2),900= 1.96 3; p>0.05).
Abundancia relativa. El patrón de abundancia de las especies de reptiles fue relativamente similar entre las 2 microcuencas, siendo Basiliscus vittatus, Sceloporus variabilis, Norops unilobatus y Sphaerodactylus glaucus las especies dominantes. Sin embargo, en Copainalá, Aspidocelis deppi fue la especie que presentó la mayor dominancia, mientras que en Bombaná, esta especie tuvo escasos registros (Fig. 3).
 y Bombaná (círculos cerrados). Las letras y números son los códigos asignados a las especies que están en el cuadro 1.")
Curvas de rango-abundancia de las especies de reptiles para las 2 microcuencas: Copainalá (rombos negros) y Bombaná (círculos cerrados). Las letras y números son los códigos asignados a las especies que están en el cuadro 1.
La complementariedad de especies entre las microcuencas estudiadas fue relativamente alta, con un recambio de especies de 58%. En Copainalá se registraron 13 especies que no se encontraron en Bombaná, mientras que en Bombaná se presentaron 8 especies que no estuvieron registradas en Copainalá (Cuadro 1).
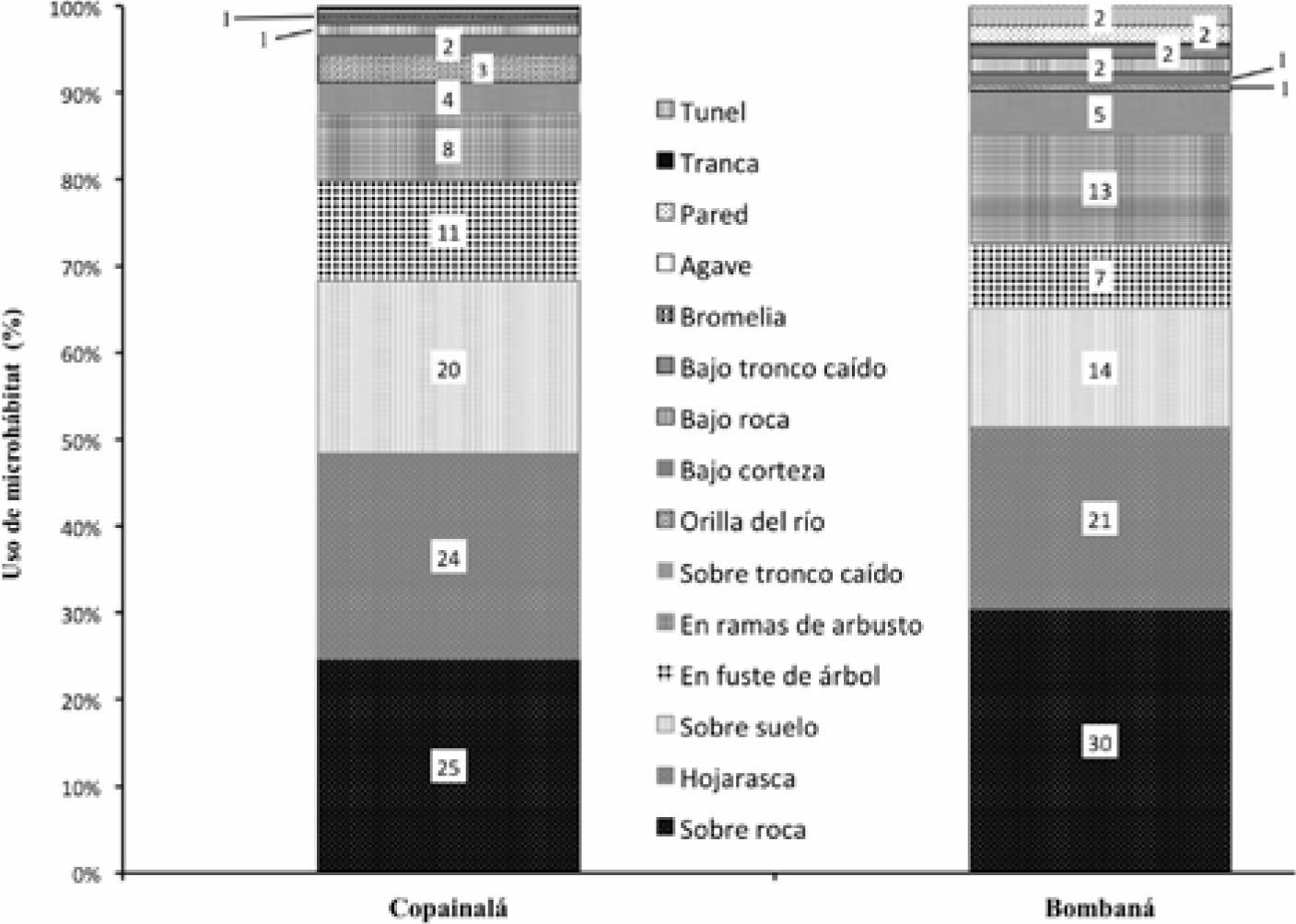
Uso de microhábitat. Se identificaron 3 diferentes tipos de microhábitats en ambas microcuencas: terrestre, arborícola y rípario; los cuales a su vez se subdividen en 15 distintos tipos (Cuadro 1, Fig. 4). El microhábitat más utilizado fue el terrestre, destacando por un uso más frecuente las categorías de “sobre roca”, “hojarasca” y “sobre suelo” (Fig. 4).

Especies en categoría de riesgo. De las 36 especies de reptiles encontradas en ambas microcuencas, 11 se encuentran en alguna categoría de riesgo según la Nom-059-Semarnat-2010. De estas especies, Coleonyx elegans, Lepidophyma chicoasensis y Boa constrictor se encuentran en la categoría de amenazadas, mientras que 8 de las especies registradas están consideradas en la categoría de protección especial (Sphaerodactylus glaucus, Ctenosaura acanthura, Iguana iguana, Imantodes cechoa, Salvadora lemniscata, Loxocemus bicolor, Crotalus simus y Kinosternon scorpioides). De acuerdo con la Unión Internacional para la Conservación de la Naturaleza (UICN), 5 especies están en la categoría de preocupación menor, estas son: Sceloporus carinatus, Coniophanes piceivittis, Mastigodryas melanolomus, Salvadora lemniscata, Micrurus browni y sólo Lepidophyma chicoasensis, especie endémica del estado de Chiapas está en la categoría de datos insuficientes (IUCN, 2012). Finalmente B. constrictor, I. iguana y Loxocemus bicolor están enlistadas en el apéndice II; especies que no necesariamente son amenazadas de peligro de extinción, pero que podrían llegar a serlo si no hay un control de su comercio; de la Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres.
DiscusiónRiqueza de especies. La región de la depresión central de Chiapas abarca las selvas secas del centro del estado de Chiapas hasta la frontera con Guatemala (Pérez-Farrera y Espinoza, 2010), registrándose en la zona 86 especies de herpetofauna, de la cuales 65 pertenecen a reptiles, siendo una de las zonas de selva seca de México con mayor riqueza herpetofaunística (García et al., 2007; García, 2010). En nuestro estudio registramos 36 especies de reptiles en total para ambas microcuencas, cerca del 55% de la fauna de reptiles registradas. A pesar que la extensión de selva seca de las 2 microcuencas estudiadas es pequeña (14 002 ha, lo que corresponde a 4.8% de la selva seca que se encuentra en la ecorregión de la depresión central, cuya extensión es de 290 719 ha [Pérez-Farrera y Espinoza, 2010]) es posible encontrar cerca de la mitad de las especies de reptiles distribuidas en la selva seca de la depresión central. Hidalgo (2008) en su estudio sobre la diversidad de reptiles en el Parque Nacional Cañón del Sumidero (PNCS), registró 43 especies de reptiles, de las cuales 32 fueron encontradas exclusivamente en selva seca. El parque tiene una extensión un poco más grande que nuestra zona de estudio (21 789 ha), lo que nos indica que la riqueza de especies que se registró en nuestro trabajo no está alejada de la riqueza de especies de la zona. De acuerdo con los modelos de curvas de acumulación de especies, se estiman 42 especies de reptiles para ambas microcuencas, aproximadamente las que se han reportado para el Cañón del Sumidero, sitio que limita al sur con nuestros sitios de muestreo.
Comparando la riqueza de especies de reptiles registradas para otras áreas de selva seca de México, encontramos que nuestra zona de estudio alberga una considerable riqueza de especies, dado que Martín-Regalado et al. (2011) en su estudio sobre la herpetofauna del cerro Guiengola, Oaxaca, registraron 31 especies de reptiles en selva seca (selva baja caducifolia, selva baja caducifolia espinosa). Mientras que Vite-Silva et al. (2010) en su estudio sobre la diversidad de herpetofauna en bosque tropical caducifolio (selva seca) de la Reserva de la Biosfera Barranca de Metztitlain, Hidalgo, registraron un total de 11 especies de reptiles. Reyna-Álvarez et al. (2010) en su estudio de la herpetofauna de Huetamo, Michoacán, donde el principal tipo de vegetación es selva seca registraron 39 especies de reptiles, realizando 2 ciclos anuales de muestreo. Mientras que Reyna et al. (2007) en el bosque tropical caducifolio de La Primavera, Jalisco reportan 34 especies de reptiles. Castro-Franco y Bustos (1994) en una revisión de la especies de lagartijas que habitan en selva baja caducifolia encontraron un total de 19 especies de lacertilios, riqueza muy parecida a la del presente estudio, donde registramos 18 especies de lacertilios. Con respecto a estudios realizados en selvas secas de América, Carvajal-Cogollo y Urbina-Cardona (2008) mencionan en su estudio de fragmentación del bosque seco tropical en Colombia, un total de 35 especies de reptiles, encontrando mayor riqueza de especies de reptiles en fragmentos más grandes con respecto a los pequeños.
Con base en los estudios citados anteriormente, nuestros resultados sugieren que ambas microcuencas presentan una riqueza de reptiles importante y esto puede estar relacionado a que el estudio fue llevado a cabo en áreas cercanas a ríos, donde se ha documentado que la vegetación riparia dentro de una matriz de selvas secas, juega un papel importante en el mantenimiento del ecosistema, aportando sitios de refugio para numerosas especies de fauna asociada (García, 2006; García y Cabrera, 2008; García, 2010; Suazo-Ortuño et al., 2011) y, por lo tanto, alberga considerable diversidad de especies de herpetofauna.
Composición de especies. Se ha mencionado que la depresión central de Chiapas alberga una alta diversidad de herpetofauna, registrándose un total de 65 especies de reptiles para esta zona (García, 2006; García et al., 2007). De éstas, en nuestro estudio se adicionan 2 registros de especies de serpientes como Spilotes pullatus y Atropoides mexicanus, que en el caso de ésta última especie, se encontró en los límites de selva seca y bosque de encino. El estudio de Hidalgo (2008) en el PNCS, el cual es parte de la ecorregión depresión central de Chiapas, agrega 5 registros adicionales a las especies que previamente García (2006) había registrado. Estas 5 especies: Corytophanes hernandezi, Scincella gemmingeri, Scincella incertum, Xenosaurus grandis y Pseustes poecilonotus, no fueron registradas en nuestro estudio.
Realizando un análisis por microcuenca, se observó que la diversidad de especies es muy similar entre ambas microcuencas; no obstante, en Copainalá se registró mayor riqueza de reptiles, 5 especies más que en Bombaná. Además, entre ambas microcuencas sólo se comparten 15 especies de reptiles, es decir, un 50%, y el resto de las especies que componen a cada microcuenca son distintas. En Copainalá se registraron 13 especies de reptiles diferentes a las encontradas en Bombaná, entre ellas están 5 especies de lagartijas: C. elegans, I. iguana, Sceloporus melanorhinus, Norops tropidonotus, L. chicoasensis. Estas especies están más asociadas a características que se presentan en la selva seca de estas zona, como son: áreas de acantilados y pendientes pronunciadas, hábitat característicos para estas lagartijas, lo cual ha sido observado también por Castro-Franco y Bustos (2003), a excepción de N. tropidonotus, la cual fue encontrada en los límites de selva seca y bosque de encino.
En el caso de las serpientes, 7 especies fueron sólo encontradas en Copainalá, entre las cuales están B. constrictor, Imantodes cenchoa, Masticophis mentovarius y S. pullatus, las cuales, además, no fueron encontradas en el trabajo de Hidalgo (2008) del PNCS. Los requerimientos de cada una de estas especies hace difícil el explicar la presencia de éstas en Copainalá y no en Bombaná. No obstante, estas especies son de hábitos arborícolas que necesitan la presencia de cobertura forestal y sitios no perturbados. La selva seca de Copainalá es relativamente menos alterada que la de Bombaná, lo cual probablemente pueda estar afectando en términos de recursos alimentarios, etc., exceptuando a M. mentovarius que está más asociada a lugares abiertos y de cultivos agrícolas. Las otras especies de serpientes encontradas sólo en Copainalá y que también fueron registradas por Hidalgo (2008) en el PNCS son: Epictia goudotii y L. bicolor, que son de hábitos fosoriales, lo que las hace más difíciles de observar y registrarlas, aunque no necesariamente son especies raras. En el caso de C. simus por lo observado en el estudio, prefiere sitios de selva seca relativamente no alterados, y en nuestro estudio no fue registrada en sitios cercanos de población antropogénica. Bombaná es una zona que tiene selva seca relativamente más fragmentada y con mayor sitios de población humana.
Con respecto a Bombaná, se encontró una sola especie de lagartija que no estuvo presente en Copainalá, Norops laevivientris, la cual se registró en los límites de selva seca con bosque de encino. En cuanto a serpientes, fueron 7 que se presentaron solamente en Bombaná, entre las cuales destacan. Leptodeira annulata, M.melanolomus y Trimorphodon biscutatus que son especies que se asocian a cuerpos de agua. En la microcuenca Bombaná los cuerpos de agua, donde se registraron estas especies, son ríos con un cauce menor al río Grijalva que cruza la microcuenca Copainalá, son más estrechos y prácticamente son arroyos que van a desembocar al río Grijalva y lo que se observó en el presente estudio, fue que estas especies están más asociadas a arroyos como los que se presentan en Bombaná. La especie de nauyaca A. mexicanus fue registrada en una zona límite de selva seca con bosque de encino y con una altitud de 1 027m. El resto de las especies de serpientes que se encontraron solamente en Bombaná fueron Drymarchon melanurum, Tropidodipsas fasciatus y M. browni, las cuales son altamente probable que puedan encontrarse en Copainalá, dado que son especies que tienen una distribución más amplia, Hidalgo (2008) también las registró en el PNCS.
Abundancia relativa. El patrón de abundancia fue muy similar en las 2 microcuencas, lo cual se observa visualmente en las pendientes que presentan las gráficas (Fig. 3). Básicamente son las mismas especies dominantes en las 2 zonas, siendo todas especies de lagartijas (A. deppii, B. vittatus, S. glaucus, N. unilobatus y S. variabilis). No obstante, el orden de cuál fue la especie más abundante es diferente, en el caso de Copainalá A. deppii fue la especie más abundante, mientras que en Bombaná, esta especie es la quinta más abundante. En contraste, en Bombaná la especie más abundante fue N. unilobatus ocupando el cuarto lugar en Copainalá. Comparando las especies de reptiles abundantes encontradas en el PNCS, éstas son un tanto diferentes, dado que S. glaucus no fue una especie abundante en el parque, de hecho es de las menos abundantes (Hidalgo, 2008). Mientras que S. variabilis,A. deppii y B. vittatus sí están dentro de las 5 especies dominantes que menciona Hidalgo (2008) en su trabajo. En general, las especies abundantes registradas en nuestro estudio son abundantes en una gran diversidad de hábitats y consideradas como especies generalistas (Urbina-Cardona y Reynoso, 2005; Urbina-Cardona et al., 2006; Hidalgo, 2008; Martín-Regalado et al., 2011), a excepción de S. glaucus. Esta especie se encuentra sujeta a protección especial por la Nom-059-Semarnat-2010 y aparentemente es poco común; sin embargo, en nuestro trabajo se encontró dentro de las 5 especies abundantes. Además, fue encontrada en una gran variedad de microhábitats, que van desde los terrestres hasta los arborícolas, siendo mayormente abundante en los microhábitat terrestres como hojarasca y sobre roca.
En lo que se refiere a especies poco comunes, en ambas microcuencas se registraron entre 8 y 9 especies para Copainalá y Bombaná, respectivamente, la mayoría fueron especies de serpientes. Generalmente, las serpientes son más difíciles de observar y ubicar, debido a sus hábitos discretos y escurridizos. Sin embargo, las abundancias de las especies tiende a cambiar si se aplica mayor esfuerzo de muestreo o aplicando diferentes métodos de muestreo que complementen la búsqueda directa (Urbina-Cardona y Reynoso, 2005).
Uso de microhábitat. Con respecto al uso de microhábitat, ambas microcuencas presentan gran diversidad de ellos, siendo que la mayor parte de los reptiles que habitan en ambas zonas son de microhábitat terrestre, utilizando principalmente “sobre roca”, “hojarasca” y “sobre suelo”, además, cerca de un 20% de los reptiles también está usando microhábitat arborícolas, como son fuste de árboles y ramas de arbustos (Fig. 4). En el estudio de Martín-Regalado (2011) también se menciona que se encontró que la mayoría de las especies que registró en selva seca utilizan mayormente el microhábitat terrestre.
Implicaciones de conservación. Nuestro estudio es un alcance sobre el entendimiento de la diversidad de especies de reptiles en un paisaje con selva seca en el estado de Chiapas, con cierto grado de alteración (Inegi, 2000). La cantidad de especies de reptiles que alberga es considerable y, por lo tanto, sugiere que como hábitats alterados pueden mantener un importante número de especies de fauna. El 30% de las especies de reptiles registradas en nuestro estudio, se encuentran en alguna categoría de riesgo. Cabe señalar que la zona limita al sur con el PNCS, el cual también alberga gran cantidad de especies de reptiles, por lo que las microcuencas Copainalá y Bombaná podrían funcionar como un corredor de especies de reptiles entre toda la región prioritaria llamada La Chacona - Cañón del Sumidero (Arriaga et al., 2000) y el corredor biológico Laguna Bélgica - Sierra El Limón - Cañón del Sumidero (Arizmendi y Márquez, 2000). Estudios futuros deben poner atención en documentar la diversidad de reptiles en diferentes zonas de selva seca a lo largo del estado de Chiapas y evaluar el papel de estos sitios para el mantenimiento de esta diversidad, dado que gran parte de selva seca no está bien representada en áreas naturales protegidas.
De manera muy especial a las personas que facilitaron nuestro trabajo en campo y permiso para acceder a sus propiedades: don Heraclio Juárez Orantes y don Pablo González López. A Ever Vázquez Pérez y David Bonilla por su ayuda en el trabajo de campo, así como a Yasminda García del Valle. Los autores agradecen el apoyo financiero otorgado por la Universidad de Ciencias y Artes de Chiapas y la Comisión Federal de Electricidad, en especial a Lidia Vázquez Hernández y Horacio Rodríguez López por sus atenciones. Adrián Sarabia Rangel por la elaboración de la figura 1. A Juan Carlos Windfield y Aaron T. Dossey por las observaciones hechas al manuscrito. A los 2 revisores anónimos por las valiosas correcciones y comentarios que mejoraron sustancialmente el manuscrito.