



La información sobre la ecología espacial de la tortuga de bisagra es limitada, pese a ser una de las especies más importantes para desarrollar estrategias de conservación del valle de Cuatro Ciénegas. Durante este trabajo se monitorizaron los movimientos diarios de 7 hembras de tortuga de bisagra de agosto a octubre de 2014 y 12 hembras de abril a junio del 2015, en el área de Los Gatos, dentro del valle de Cuatro Ciénegas. Se estimó el ámbito hogareño mediante el polígono mínimo convexo y el método de Kernel fijo al 95%; las áreas núcleo de actividad fueron estimadas mediante el Kernel fijo al 50%. La direccionalidad fue estimada mediante la prueba de Rao. El ámbito hogareño varió entre ambos periodos mediante el polígono mínimo convexo, mientras que el método de Kernel fijo al 95% no presentó diferencias entre ambos periodos; lo mismo ocurrió para las áreas núcleo de actividad. Por otra parte, las hembras no presentaron movimientos al azar durante todo el muestreo. Los resultados evidencian que la especie ocupa áreas de mayor tamaño que las planteadas en otros trabajos. La conservación de hábitat para la especie debe de contemplar la interconexión de los cuerpos de agua presentes en el sitio Los Gatos.
An understanding of the spatial ecology of the Coahuilan box turtle is critical for the development of successful habitat conservation strategies for this imperiled species. Regrettably, the spatial ecology of this species is poorly understood. We monitored the daily movements of 7 females Coahuilan box turtles from August to October 2014 and 12 female turtles from April to June 2015, in the wetland Los Gatos, inside of Cuatro Ciénegas valley. We estimated home range size using both minimum convex polygon and 95% fixed Kernel. Core activity areas were estimated by 50% fixed Kernel. Directionality was estimated by Rao's test to evaluate mean directionality of movements (locations) in relationship with the center of activity. Minimum convex polygon home ranges were different for the 2 sampling periods, while 95% fixed Kernel showed no differences between the 2 sampling periods. The 50% fixed Kernel core activity areas also did not show differences between the sampling periods. Moreover, female Terrapene coahuila movements did not make random during either sampling period. Our home range analyses indicate that the species occupies larger areas than estimated by previous authors. Habitat conservation for this species must consider the connectivity of the ponds in the Los Gatos study area.
Terrapene coahuila es una especie endémica de México cuya distribución se restringe al valle de Cuatro Ciénegas, Coahuila. Está catalogada como amenazada según la Norma Oficial Mexicana (Semarnat, 2010) y en peligro de extinción por parte de la Unión para la Conservación de la Naturaleza (Van Dijk, Flores-Villela y Howeth, 2007). La especie ha mostrado un decremento poblacional en los últimos años. Estudios previos mostraron abundancias promedio de 148ind/ha (Brown, 1968), mientras que para el 2008 se estimaron abundancias de 0.66 a 66ind/ha (Howeth, McGaugh y Hendrickson, 2008). Sin embargo, datos más recientes indican que el sitio de mayor abundancia de T. coahuila apenas presenta densidades de 3ind/ha (Salas-Westphal et al., 2011). La reducción en la abundancia de la tortuga es atribuida principalmente al deterioro ambiental de los cuerpos de agua del área de distribución, ocasionado por la extracción de agua del valle de Cuatro Ciénegas, lo que a su vez fomenta la pérdida de hábitat de la tortuga. Bajo este contexto, y al tratarse de una especie de distribución restringida, cuya vida está ligada a los cuerpos de agua, es relevante conocer los requerimientos sobre la ecología espacial y direccionalidad de los movimientos de las hembras de la tortuga de bisagra, con la finalidad de comprender y plantear estrategias de conservación encaminadas a la protección del hábitat requerido por la especie. Por tal motivo, el objetivo de este trabajo fue estimar el ámbito hogareño de las hembras grávidas y no grávidas dentro del sitio llamado Los Gatos, en el valle de Cuatro Ciénegas, así como conocer los promedios de las distancias recorridas y la direccionalidad de los movimientos realizados por las hembras de la especie. Dado el extendido periodo reproductivo de la especie y que la anidación puede ocurrir desde mayo hasta septiembre, el objetivo se planteó para 2 temporadas que contemplan el periodo de copulación y anidación de la especie (agosto-octubre 2014 y abril-junio 2015).
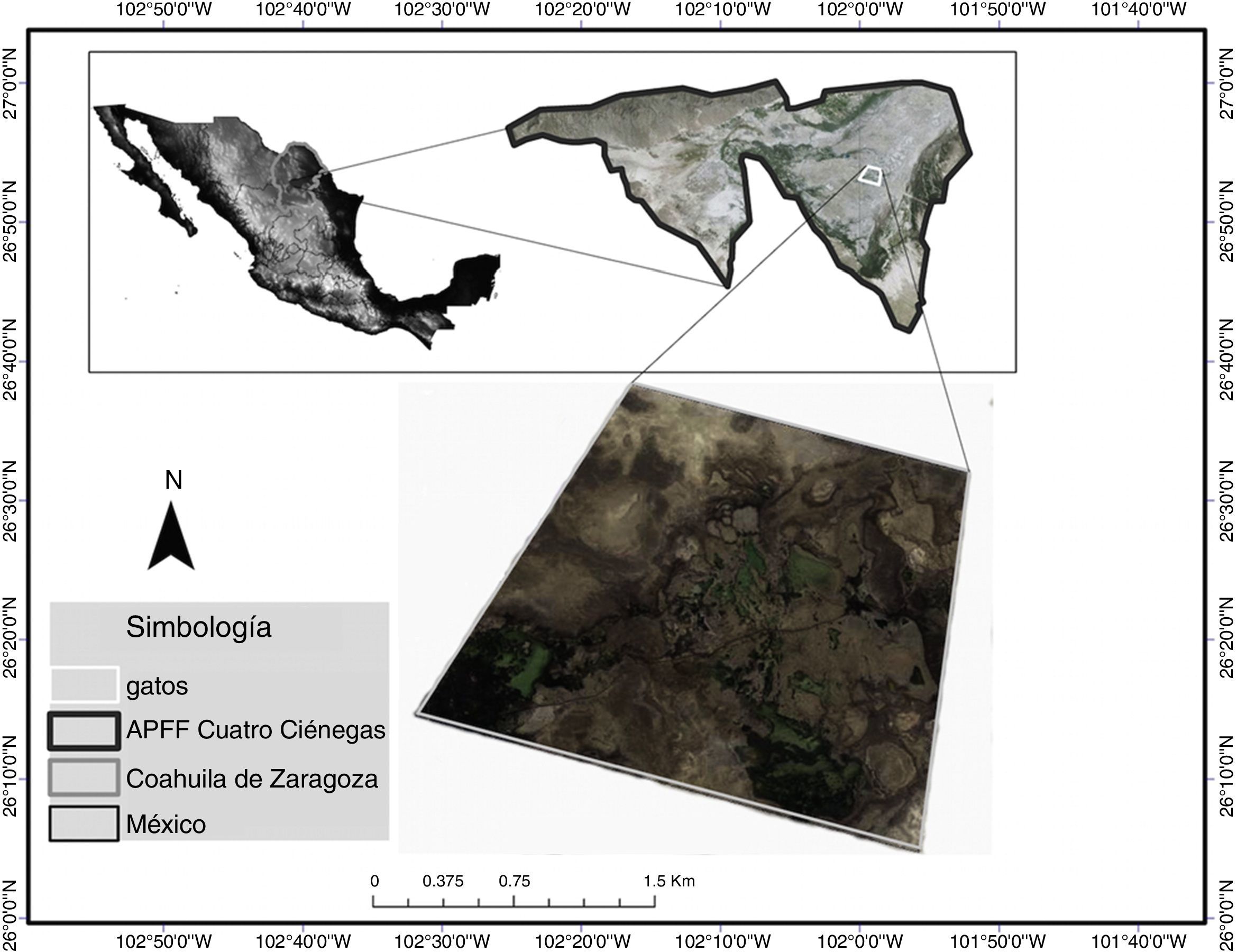
Materiales y métodosEl sitio Los Gatos cuenta con una extensión aproximada de 4.1km2 (fig. 1). Es uno de los sitios con mayor abundancia de tortugas (Salas-Westphal et al., 2011) dentro del Área de Protección de Flora y Fauna de Cuatro Ciénegas. Tiene una altitud promedio de 700m snm. El clima es seco a semicálido, con una precipitación anual de 100 a 400mm. La temperatura dentro del valle oscila entre los 12° y los 30°C (INE, 1999).

El trabajo de campo se llevó a cabo en 2 estaciones, de agosto a octubre de 2014 y de abril a junio de 2015. Ambas estaciones coinciden con el periodo de copulación y oviposición de la especie. La literatura identifica 2 periodos en los que ocurre la copulación (abril-mayo y agosto-septiembre), mientras que la oviposición ocurre principalmente en mayo (primavera), aunque la especie tiene el potencial de realizar hasta 3 puestas al año, por lo que durante gran parte del año es posible observar hembras grávidas y no grávidas. Debido a lo anterior, en este trabajo se consideraron 2 estaciones donde fue más probable reconocer hembras en distintas fases reproductoras (Brown, 1968; Cerda y Waugh, 1992). Durante cada estación se realizó una búsqueda directa de hembras considerando el dimorfismo sexual descrito para la especie y para otros emídidos (Brown, 1968; Dodd, 2001). Se georreferenció la posición de cada individuo (Salas-Westphal et al., 2011) y se tomaron medidas de largo y ancho de caparazón y plastrón, así como el peso (Castañeda-Gaytán et al., 2012). Se consideraron únicamente las hembras con una longitud de caparazón superior a 120mm. A las hembras encontradas durante el primer periodo de muestreo se les realizó palpación manual en búsqueda de huevos calcificados (Rueda-Almonacid et al., 2007). Durante el segundo periodo de muestreo, además de la palpación se realizó una ecografía a cada ejemplar para la identificación de folículos y/o huevos con un equipo de ultrasonido marca SonoScape modelo A6 con una frecuencia de 7.5 a 10Mhz (Costa et al., 2009; Schildger et al., 1994). Cada ultrasonido realizado fue fotografiado para conocer el estado reproductivo de los individuos. Los huevos encontrados se midieron para deducir si estaban próximos a su puesta. Posteriormente, se colocaron radiotransmisores tipo VHF marca Wildlife Materials, modelo SORP-023, con un peso de 10g (menor al 8% del peso total) y una dimensión de 27×15×10mm. En el primer periodo de muestreo se colocaron 7 radiotransmisores, mientras que en el segundo periodo de muestreo se colocaron 12. De esos, 10 fueron hembras con radiotransmisores VHF y 2 fueron hembras con radiotransmisores con GPS marca Telenax con un peso de 35g. Todas las hembras pesaron más de 400g y tuvieron presencia de huevos en el oviducto de dimensiones próximas al desove. Los radiotransmisores se colocaron a un costado del caparazón con la antena libre y se aseguraron con resina epóxica marca Devcon. Para recibir la señal de los transmisores, cada tortuga fue rastreada diariamente de 30 a 45 días para el primer muestreo y de 16 a 46 días durante el segundo. La búsqueda se realizó con un escáner de frecuencias modelo RX-TLNX y una antena Yagi de 3 elementos (Telenax, Playa del Carmen, México). Las coordenadas se registraron con un GPS modelo eTrex 30 de la marca Garmin (Garmin Inc., Olathe, Kansas, EUA) en el sistema Universal Transversa Mercator.
Las localizaciones de cada individuo se analizaron en el programa Biotas 2.0 (ESS, 2004), compatible con Windows 7. En el programa antes mencionado se estimó el ámbito hogareño por el polígono mínimo convexo y el método de Kernel fijo al 95% (Refsnider, Strickland y Janzen, 2012). Se calcularon las áreas de mayor actividad de las hembras (áreas núcleo) mediante el método de Kernel fijo al 50%. A su vez, se estimó la distancia entre localizaciones obtenidas cada 24h y se calculó el centro de actividad de cada individuo por medio de la media armónica; posteriormente, se realizó un segundo análisis para conocer los ángulos de dirección con respecto de la media armónica. Finalmente, se elaboraron los mapas mediante el programa ArcMap 10.1 (ESRI, 2012).
Para realizar las comparaciones de ámbito hogareño entre las hembras grávidas y las no grávidas se calcularon los promedios para cada periodo de muestreo; de la misma manera, se realizó el promedio de las distancias diarias recorridas por las hembras de cada condición reproductiva en cada periodo de muestreo, y con esos datos se llevó a cabo una prueba de normalidad de Shapiro-Wilk. Se realizaron las comparaciones de medias entre muestreos y estados reproductivos por medio de pruebas t de Student para muestras independientes no pareadas una vez que se probó la normalidad y la homogeneidad de la varianza de los datos (Zar, 1999). Estas comparaciones se realizaron para los polígonos de hembras grávidas vs. no grávidas de manera intra e interestacional. El análisis estadístico se llevó a cabo en el programa SPSS 17.0 (SPSS, 2008). El análisis de direccionalidad de movimiento se llevó a cabo en el programa Oriana 4 (Plummer, 2014) para Windows, utilizando la prueba de Rao para establecer si los movimientos con respecto a la media armónica fueron aleatorios o dirigidos hacia un punto. Los mapas de ámbito hogareño se generaron en el programa ArcMap 10.1 (ESRI, 2012) y las imágenes del terreno se tomaron de mapas disponibles en Global Mapper versión 16 (BMG, 2015). Todos los análisis se hicieron con una α=0.05.
ResultadosSe obtuvieron un total de 288 localizaciones correspondientes a 7 hembras durante agosto a octubre (2014) (92 coordenadas corresponden a 2 hembras grávidas y 196 corresponden a 5 hembras no grávidas). Para el segundo periodo de muestreo se registraron 388 localizaciones para 12 hembras (252 localizaciones corresponden a 8 hembras grávidas y 136 a 4 hembras no grávidas). El número de radiolocalizaciones obtenidas por individuo osciló entre 16 y 45 (promedio de 34).
El ámbito hogareño estimado por el polígono mínimo convexo para el periodo de agosto a octubre fue de 31.43±20.63ha en promedio para las 7 hembras monitorizadas, con una media de 37.05±28.82ha para hembras grávidas y de 29.18±20.22ha para hembras no grávidas (fig. 2). No hubo diferencias significativas entre las 2 condiciones reproductivas (t=0.42, g. l.=5, p=0.69). Para el periodo de abril a junio del 2015 el ámbito hogareño promedio de 12 hembras fue de 12.05±11.55ha, con un promedio de 14.07±12.23ha para hembras grávidas y de 8.03±10.41ha para las no grávidas (fig. 3). No se observaron diferencias significativas (t=0.84, g. l.=10, p=0.42) entre los 2 grupos de hembras.


El ámbito hogareño estimado para el periodo de agosto a octubre fue más extenso en comparación con el periodo de abril a junio (2015) (t=2.64, g. l.=17, p=0.01). Con el método de Kernel fijo al 95% se estimó un área promedio de 6.16±11.14ha para todas las hembras durante el periodo de agosto a octubre; las hembras grávidas promediaron 1.40±0.96ha, mientras que las hembras no grávidas promediaron 8.06±13.05ha (fig. 4). En las estimaciones obtenidas por este método no se observaron diferencias significativas entre ambas categorías (t=0.68, g. l.=5, p=0.52). Para el periodo de abril a junio (2015) las hembras presentaron un ámbito hogareño promedio de 0.95±0.50ha; 1.02±0.25ha para hembras grávidas y 0.82±0.87ha para hembras no grávidas, sin presentar diferencias significativas entre ambos grupos (t=0.63, g. l.=10, p=0.53). Los promedios de ámbito hogareño globales entre periodos tampoco presentaron diferencias significativas (t=1.64, g. l.=17, p=0.11; fig. 5).
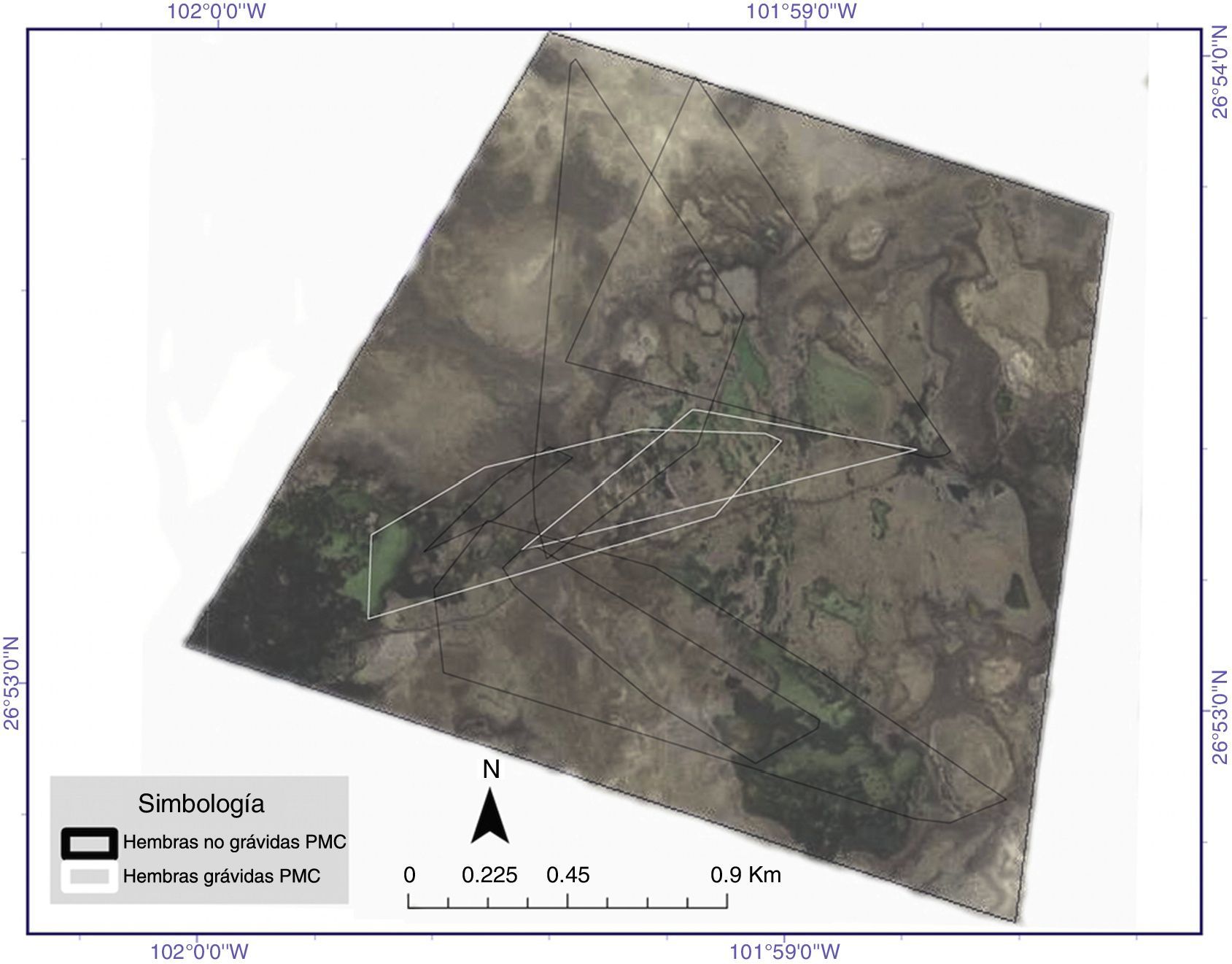
 con zonas consideradas núcleo o de mayor intensidad de uso (al 50%) por el método de Kernel fijo obtenidos para los diferentes individuos de Terrapene coahuila estudiados. En la leyenda del interior de la imagen se aprecian los individuos y su condición reproductora. Periodo de agosto a octubre de 2014.")
Áreas de actividad (al 95%) con zonas consideradas núcleo o de mayor intensidad de uso (al 50%) por el método de Kernel fijo obtenidos para los diferentes individuos de Terrapene coahuila estudiados. En la leyenda del interior de la imagen se aprecian los individuos y su condición reproductora. Periodo de agosto a octubre de 2014.
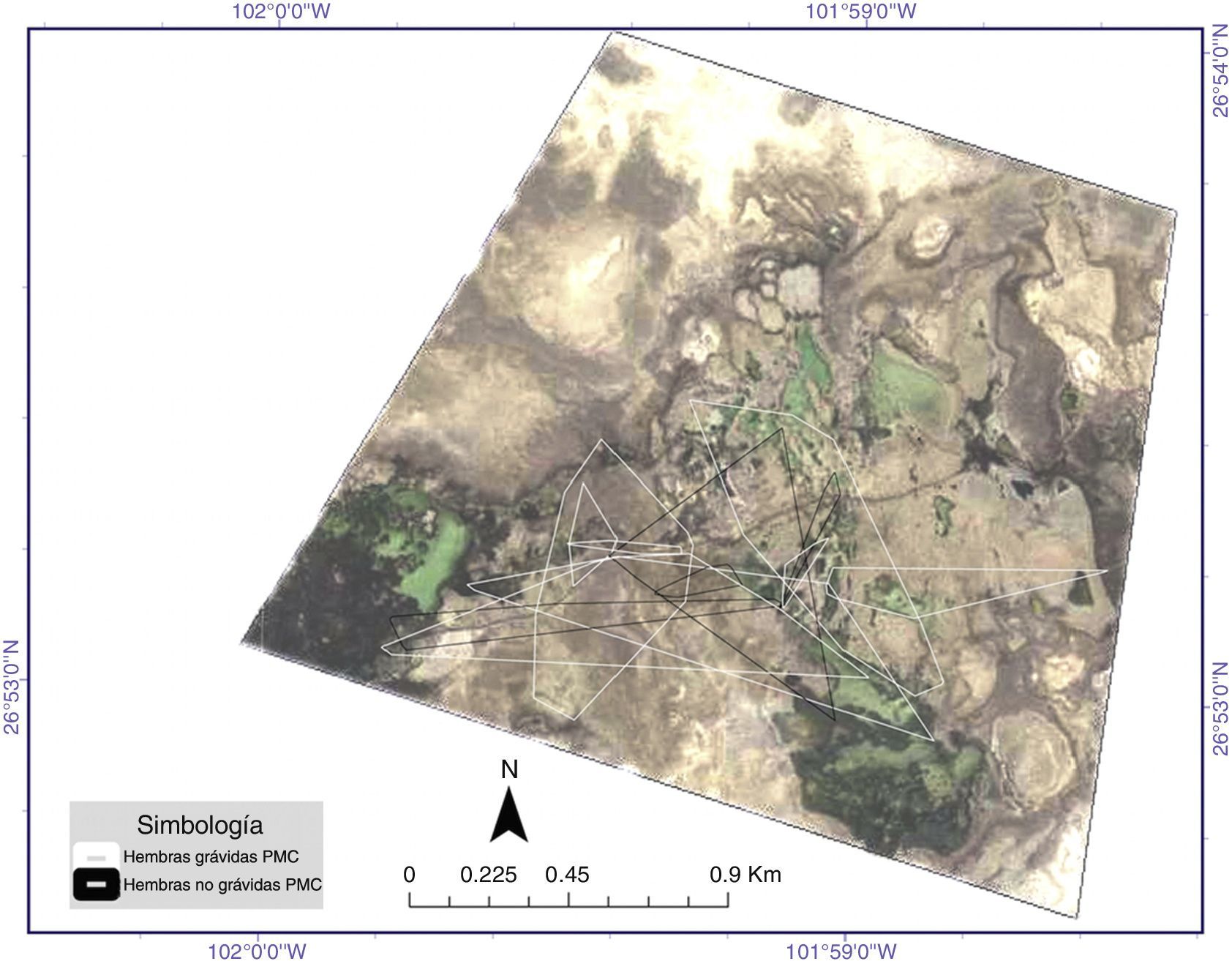
 con zonas consideradas núcleo o de mayor intensidad de uso (al 50%) por el método de Kernel fijo obtenidos para los diferentes individuos de Terrapene coahuila estudiados. En la leyenda del interior de la imagen se aprecian los individuos y su condición reproductora. Periodo de abril a junio de 2015.")
Áreas de actividad (al 95%) con zonas consideradas núcleo o de mayor intensidad de uso (al 50%) por el método de Kernel fijo obtenidos para los diferentes individuos de Terrapene coahuila estudiados. En la leyenda del interior de la imagen se aprecian los individuos y su condición reproductora. Periodo de abril a junio de 2015.
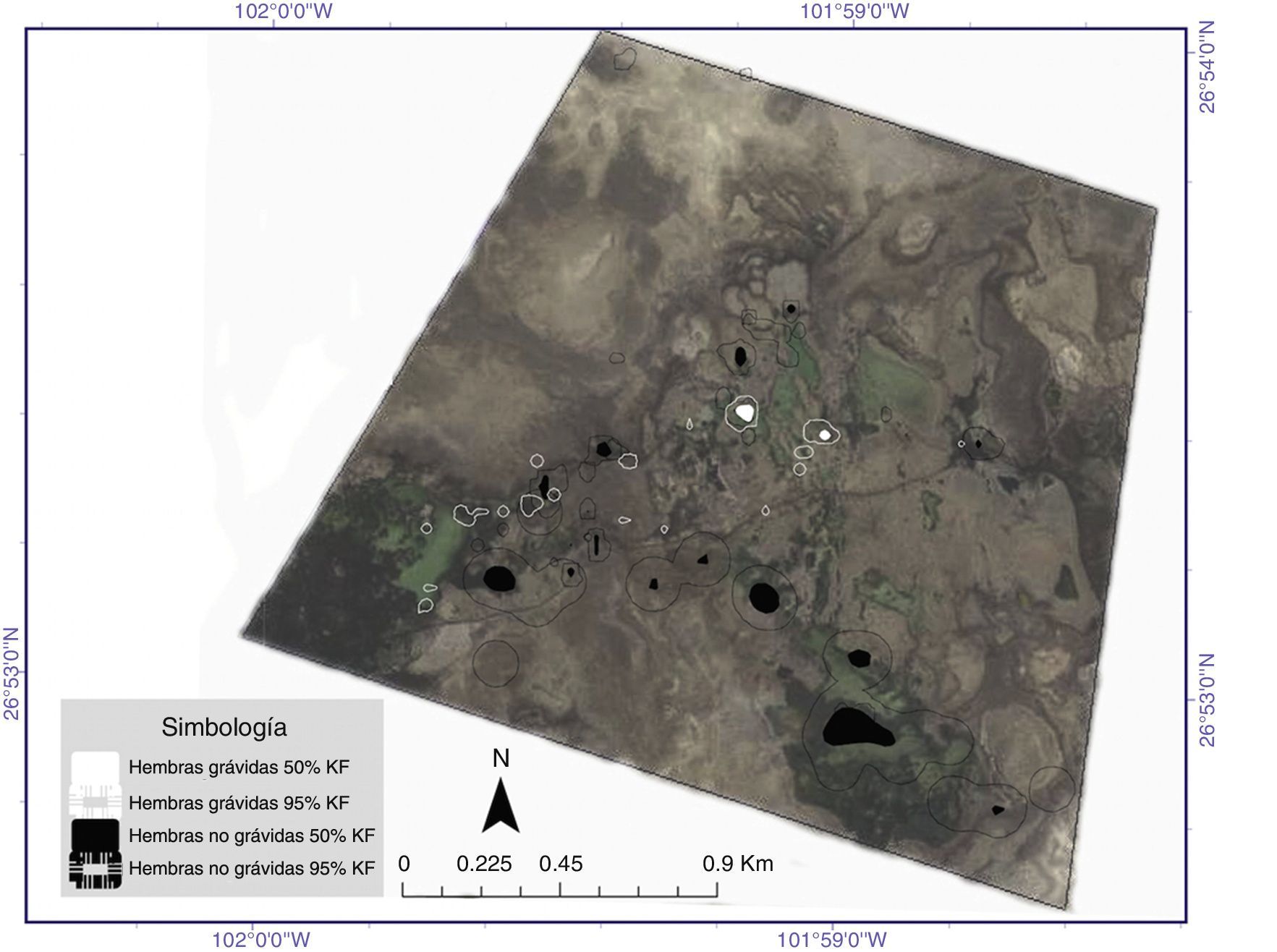
El cálculo de las áreas núcleo al 50% mediante el método de Kernel estableció superficies de 0.55±1.06ha en promedio para las hembras durante el periodo de agosto-octubre. De forma independiente, las superficies indicaron áreas núcleo de 0.10±0.07ha para hembras grávidas y de 0.73±1.25ha para las hembras no grávidas, sin diferencias significativas (t=−0.67, g. l.=5, p=0.53). Para el segundo periodo de muestreo se obtuvo un área promedio de 0.06±0.06ha; 0.04±0.02ha corresponde a hembras grávidas y 0.09±0.10ha a las no grávidas. No se detectaron diferencias entre ambas categorías (t=1.28, g. l.=10, p=0.22). No se observó ninguna diferencia significativa entre los periodos de muestreo para las superficies de las áreas núcleo de las hembras en conjunto (t=1.17, g. l.=17, p=0.25).
Las hembras de T. coahuila obtuvieron un promedio de distancia diaria mínima recorrida de 109.04±58.57m para el primer periodo de muestreo; las hembras grávidas recorrieron 106.68±49.27m y las no grávidas 109.98±67.34m en promedio, sin mostrar diferencias entre ambas categorías reproductivas (t=0.06, g. l.=5, p=0.95). Para el segundo periodo de muestreo se obtuvo un promedio de distancia diaria recorrida de 75.26±40.69m para las hembras; las hembras grávidas se desplazaron en promedio 76.59±40.12m, mientras que el promedio de las no grávidas fue de 72.60±47.99m. No hubo diferencias significativas entre categorías reproductoras (t=0.15, g. l.=10, p=0.88). Por otra parte, tampoco se encontraron diferencias en los promedios de distancias diarias recorridas en ambos periodos de muestreo (t=1.48, g. l.=17, p=0.15).
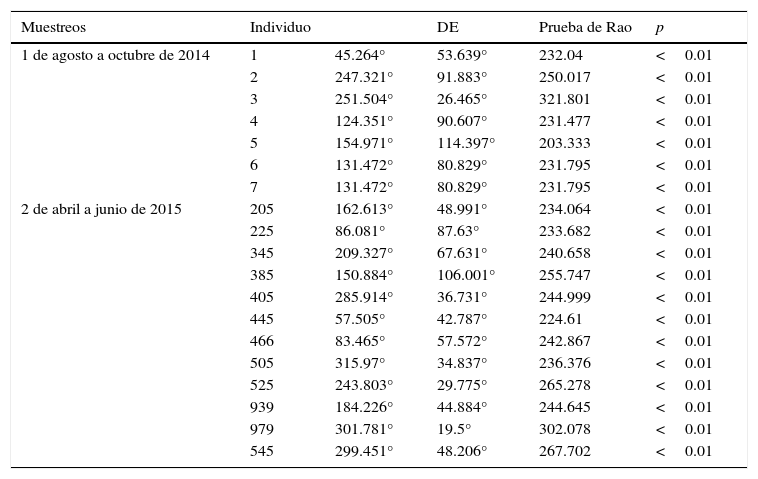
Las pruebas de direccionalidad de Rao evidencian que ninguno de los 19 individuos de T. coahuila estudiados durante ambos periodos de muestreo presentó movimientos al azar (p<0.01, tabla 1). Los ejemplares de T. coahuila mantuvieron la dirección a zonas de humedal, con cuerpos de agua y vegetación cercana.
Resultados de la prueba de Rao (media de los ángulos para cada individuo durante ambos periodos de muestreo, así como su desviación estándar).
| Muestreos | Individuo | DE | Prueba de Rao | p | |
|---|---|---|---|---|---|
| 1 de agosto a octubre de 2014 | 1 | 45.264° | 53.639° | 232.04 | <0.01 |
| 2 | 247.321° | 91.883° | 250.017 | <0.01 | |
| 3 | 251.504° | 26.465° | 321.801 | <0.01 | |
| 4 | 124.351° | 90.607° | 231.477 | <0.01 | |
| 5 | 154.971° | 114.397° | 203.333 | <0.01 | |
| 6 | 131.472° | 80.829° | 231.795 | <0.01 | |
| 7 | 131.472° | 80.829° | 231.795 | <0.01 | |
| 2 de abril a junio de 2015 | 205 | 162.613° | 48.991° | 234.064 | <0.01 |
| 225 | 86.081° | 87.63° | 233.682 | <0.01 | |
| 345 | 209.327° | 67.631° | 240.658 | <0.01 | |
| 385 | 150.884° | 106.001° | 255.747 | <0.01 | |
| 405 | 285.914° | 36.731° | 244.999 | <0.01 | |
| 445 | 57.505° | 42.787° | 224.61 | <0.01 | |
| 466 | 83.465° | 57.572° | 242.867 | <0.01 | |
| 505 | 315.97° | 34.837° | 236.376 | <0.01 | |
| 525 | 243.803° | 29.775° | 265.278 | <0.01 | |
| 939 | 184.226° | 44.884° | 244.645 | <0.01 | |
| 979 | 301.781° | 19.5° | 302.078 | <0.01 | |
| 545 | 299.451° | 48.206° | 267.702 | <0.01 |
T. coahuila no presentó diferencias en el ámbito hogareño de hembras grávidas frente a no grávidas, a diferencia de la especie Emydoidea blandingii (Innes, Babbitt y Kanter, 2008; Millar y Blouin-Demers, 2011), en donde se han reportado diferencias atribuidas a los desplazamientos de las hembras grávidas durante la búsqueda de sitios para anidación. Sin embargo, el hecho de que no se observen diferencias en el ámbito hogareño ni en la distancia promedio desplazada por hembras grávidas y no grávidas en T. coahuila puede deberse parcialmente a 2 factores: 1) a la alta variación observada en el ámbito hogareño, que puede a su vez responder a preferencias individuales asociadas a la distribución heterogénea e irregular de los recursos necesarios para los individuos (Curtin, 1997; Kapfer, Muñoz, Groves y Kirk, 2013); las diferencias tanto en la distribución como en la configuración del hábitat han sido observadas en especies como Clemmys guttata y Chrisemys picta (Bowne, Bowers y Hines, 2006; Milam y Melvin, 2001), lo que resulta en áreas de actividad muy variables entre individuos y entre estaciones; 2) a que los sitios de anidación de las hembras grávidas posiblemente se ubican en la periferia de las pozas (considerando 10 observaciones de nidos depredados a distancias menores de 10m del cuerpo de agua), lo que probablemente influye de manera considerable en la extensión de sus áreas de actividad hacia el exterior de los humedales. En este sentido, los datos evidencian movimientos diarios regulares que variaron entre estaciones de 109m (agosto-octubre) y 72m (abril-junio) en promedio y que posiblemente dependen de requerimientos individuales asociados con la condición del humedal en el que se encuentran (nivel de agua, vegetación circundante a las pozas). Baker (2009) destaca para Terrapene carolina que sus distancias diarias recorridas pueden variar significativamente entre sexos, entre clases reproductivas y entre periodos específicos del año. Sin embargo, para T. coahuila no es posible determinar si la actividad restringida a los humedales anula estas variaciones (sexo, clase reproductiva, periodos del año) o si depende directamente de la configuración y la extensión de las pozas. Por otra parte, la tendencia en algunas especies a mantener áreas de actividad relativamente pequeñas se asocia a un ambiente productivo con suficiente disponibilidad de alimento (Rowe, 1987); sin embargo, en el caso de las hembras de T. coahuila, su extensión espacial evidencia una amplia necesidad de superficie que puede deberse a diferentes factores, como los siguientes: distribución heterogénea de los recursos asociados a los cuerpos de agua, los cuales sufren una dinámica de crecimiento y reducción constante que depende de los fenómenos meteorológicos globales. Estas fluctuaciones pueden generar variaciones en la calidad y la disponibilidad de alimento y/u otros requerimientos asociados directamente con los humedales (por ejemplo, microclima, cubierta de vegetación acuática o periférica). Debido a esto, el hecho de observar ámbitos hogareños tan extensos en comparación con los de especies del género como T. ornata y T. carolina (Bernstein y Richtsmeier, 2007; Kapfer et al., 2013) puede indicar el uso de humedales distantes en la búsqueda de recursos específicos; esta diferencia en extensión de ámbitos hogareños entre especies del mismo género puede ser debida a los hábitats que ocupan las especies. Esta observación se enfatiza al considerar que las estimaciones por el método de Kernel al 50% muestran múltiples áreas núcleo, localizadas en diferentes humedales o bien en diferentes puntos de un mismo humedal.
Se ha planteado para algunas especies del género Terrapene que la densidad de tortugas puede asociarse negativamente con el ámbito hogareño (Stickel, 1989). Los datos de Brown (1968) para T. coahuila y los del presente estudio se ajustan a esta tendencia, en donde los ámbitos hogareños del primero (514.7m2 según la conversión de un radio de 12.8m y en geometría circular) se asociaron con una densidad de aproximadamente 148ind/ ha, mientras que en el sitio de estudio del presente trabajo se ha estimado una abundancia de 3ind/ha (Salas-Westphal et al., 2011), aunque las diferencias de ámbito hogareño entre sexos o hembras grávidas y no grávidas han sido atribuidas a estrategias de búsqueda de pareja o sitios para anidación en diversas especies (Kapfer et al., 2013). La ausencia de dichas diferencias en el presente estudio puede deberse al método y al tamaño de la muestra (Bernstein y Richtsmeier, 2007), o bien a la posibilidad de que la mayoría de los movimientos se restringen hacia los cuerpos de agua y entre los cuerpos de agua (Howeth y Brown, 2011), y a que la búsqueda de sitios de anidación puede no contribuir con la estimación diferencial del ámbito hogareño de hembras grávidas frente a no grávidas.
La diferencia en la estimación de las áreas de actividad de los individuos evaluados por Brown (1968) y la del presente estudio depende, en primera medida, de los métodos de estimación utilizados (método circular vs. polígono mínimo convexo y Kernel), así como del número de relocalizaciones obtenidas por individuo. En este caso, las estimaciones obtenidas a partir de un seguimiento diario durante un promedio de 34 días consecutivos apuntan a una muestra comparable a la de otros estudios (Baker, 2009; Bernstein y Richtsmeier, 2007).
Por otra parte, la discrepancia en cuanto a los datos entre ambos estudios es debida a la disposición de recursos en el sitio de muestreo de Brown, ya que su sitio de estudio constaba de 13 pozas estables durante el año, mientras que en nuestro sitio de estudio las pozas tienen un nivel de agua variable durante las estaciones del año. Estimaciones previas del ámbito hogareño de T. coahuila permiten observar ámbitos de menor tamaño (Salas-Westphal et al., 2011) que los encontrado en el presente trabajo. Tales diferencias se pueden atribuir a la falta de consideración de la condición reproductiva y a que en este trabajo solo se tomaron en cuenta hembras. Al mismo tiempo, y a diferencia de los estudios previos, la consideración de forma independiente de las hembras grávidas y las no grávidas agrega la posibilidad de descartar el efecto de dicha condición en expresiones de ámbito hogareño diferenciales o la influencia de otros factores no considerados en este estudio.
Lo anterior evidencia que la especie tiene requerimientos espaciales altamente variables con múltiples factores involucrados. Sin embargo, es necesario un estudio a largo plazo que involucre diferentes grupos de edad y sexo para poder determinar los patrones de actividad de la especie. Sin embargo, las variaciones encontradas dentro de este trabajo pueden relacionarse con eventos meteorológicos que parecen mostrar una fuerte influencia en la dinámica espacial de la especie.
Castañeda-Gaytán et al. (2012) reportaron ámbitos hogareños para hembras de 3.8ha para los meses de febrero a marzo del 2011, de 3.32ha para los meses de noviembre a diciembre del 2011 y de 4.83ha durante los meses de agosto a octubre del 2012 (los que resultan ser menores que los encontrados durante este trabajo: 31.43±20.63ha para el primer muestreo y 12.05±11.55ha para el segundo muestreo). Lo mismo se observó mediante el método de Kernel, con un ámbito hogareño de 0.7ha para hembras de febrero a marzo y de 0.23ha para el periodo de noviembre a diciembre del 2011 (Castañeda-Gaytán et al., 2012). Las diferencias observadas en dichas comparaciones se asocian con las condiciones ambientales, pues durante el año 2010, el paso del huracán Alex generó lluvias intensas (200 a 500mm en 24h) en Coahuila (Hernández-Unzón y Bravo-Lujano, 2010), recargando en el área de estudio numerosos cuerpos de agua de Cuatro Ciénegas. Estos cuerpos de agua mantuvieron niveles de agua estables durante el año 2011, pero para el año 2014 (un año de sequía para el país y, en particular, para Coahuila) algunos humedales habían desaparecido y otros se habían reducido hasta el punto de aislarse completamente, contribuyendo con una mayor heterogeneidad del hábitat para la especie.
Las variaciones interanuales también se aprecian dentro del mismo año (Becerra-López, datos sin publicar) y, en este caso, representan un factor con potencial de influir en el ámbito hogareño de las hembras de T. coahuila durante las 2 estaciones evaluadas. Se observó fluctuación en el nivel del agua de las pozas a lo largo del tiempo de estudio; sin embargo, los datos de los piezómetros colocados para la monitorización de agua por parte de Conanp no muestran esta variación, lo que puede deberse a que estos aparatos están colocados en un humedal estable dentro del sitio Los Gatos, mientras que las hembras estudiadas mantuvieron movimientos en la periferia de este humedal.
Los requerimientos espaciales, así como el reconocimiento de los diferentes factores vinculados con el ámbito hogareño de las hembras de T. coahuila, reflejan una dinámica compleja y posiblemente asociada con la temporalidad de las pozas. Al mismo tiempo, las estimaciones de este estudio muestran la necesidad de conservar amplias extensiones de humedales con interconectividad y con estabilidad espaciotemporal para favorecer la permanencia de la especie.
Agradecemos al Área de Protección de Flora y Fauna de Cuatro Ciénegas, Coahuila, por su invaluable ayuda en la realización de este trabajo, en especial a Juan Carlos Ibarra y a Gustavo Chavez; a su vez, agradecemos a Conanp por los proyectos PROMOBI 2012, 2013 y 2014, que ayudaron a la realización de este proyecto. Por otra parte, agradecemos la beca otorgada por Conacyt a Alejandra Cueto, y al North American Box Turtle Conservation Committee por el premio Lucille Stickell 2014. A todas las personas que nos apoyaron directa e indirectamente en la realización de este proyecto. Un agradecimiento al equipo de trabajo del Laboratorio de Herpetología de la FCB-UJED.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.