



El registro fósil de los peces de agua dulce en México, si bien es escaso, se ha visto incrementado durante los últimos años. Estos fósiles se conocen en 39 localidades, de las cuales 4, ubicadas en el norte del país, son cretácicas y cuentan con restos de cf. †Melvius sp., Amiidae, Lepisosteidae y †Ceratodus sp., grupos que no vuelven a registrarse como fósiles en periodos posteriores. Las demás localidades son cenozoicas (Oligoceno a Pleistoceno) y poseen faunas modernas; se ubican principalmente en la Faja Volcánica Transmexicana y la mayoría son pleistocénicas. La diversidad de peces fósiles incluye 12 familias y por lo menos 26 géneros y 39 especies, y de estas últimas, algunas únicamente son conocidas como fósiles. La ubicación de las localidades cenozoicas con relación a los drenajes e ictiofauna actuales sugiere eventos de cambios en la hidrología de algunas cuencas recientes. Para ampliar el conocimiento sobre este grupo de peces se requiere realizar trabajo prospectivo activo en las zonas menos exploradas, como los bolsones del norte y las cuencas tropicales del sur de México, aplicar técnicas de recuperación y de estudio más finas, así como incorporar las nuevas técnicas para análisis moleculares e isotópicos, entre otros.
The fossil record of Mexican freshwater fishes, although scarce, has increased in recent years. These fossils are known in 39 localities, 4 of which are Cretaceous and carry remains of cf. †Melvius sp., Amiidae, Lepisosteidae and †Ceratodus sp., groups no longer recorded as fossil material in subsequent periods. The remaining localities belong to the Cenozoic Era (Oligocene to Pleistocene) and contain modern faunas; most of them are located in the Mexican Volcanic Belt and most are from Pleistocene deposits. The diversity of fossil fish includes 12 families and at least 26 genera, and 39 species. Only some of the latter are known as fossils. The position of the Cenozoic localities in relation to existing drainage and fish communities suggests events of change in the hydrology for some recent basins. In order to broaden the knowledge on this group of fishes, it is required to perform active prospective work in less explored areas such as Northern endorheic drainages and Southern tropical basins of Mexico, in addition to applying finer recovery and study techniques, as well as incorporating new procedures for, among others, molecular and isotopic analyses.
Los peces de agua dulce viven normalmente en aguas con salinidad no mayor a 5 partes por 1,000 (gramos). De este grupo descartamos a las especies que generalmente viven en aguas marinas, pero que eventualmente pasan algunas temporadas en aguas de menor salinidad. Los peces dulceacuícolas están confinados a cuerpos y corrientes de agua continentales, por lo que su dispersión necesariamente implica la existencia de cambios geográficos en las conexiones entre diferentes sistemas hidrológicos, que se dan tanto en las cabeceras y partes medias de las cuencas como en las partes bajas de las mismas. Por la primera vía (por ejemplo, piratería de afluentes) se dispersan principalmente los peces primarios o dulceacuícolas obligados, que en México son generalmente peces de estirpe neártica, y por la segunda (por ejemplo, desbordamiento con invasión de drenajes) los peces secundarios o dulceacuícolas con una mayor tolerancia a aumentos temporales de la salinidad, que generalmente son peces de origen neotropical. La presente contribución actualiza y analiza la información disponible sobre la paleoictiofauna dulceacuícola mexicana, basada en una investigación documental, y presenta algunas reflexiones sobre la distribución en el pasado de algunos grupos.
Materiales y métodosLas fuentes bibliográficas consideradas fueron libros, capítulos de libros, artículos científicos, tesis, resúmenes de congreso e informes técnicos. La búsqueda bibliográfica partió de las recopilaciones previas relativas a este grupo fósil en México (e.g., Alvarado-Ortega, González-Rodríguez, Blanco-Piñón, Espinosa-Arrubarrena y Ovalles-Damián, 2006; Arroyo-Cabrales et al., 2008; Barrios-Rivera, 1985; Cavender, 1986; González-Rodríguez, Espinosa-Arrubarrena y González-Barba, 2013; Guzmán y Polaco, 2009; Maldonado-Koerdell, 1949; Miller y Smith, 1986; Smith, 1981), con lo cual se recuperó la literatura original revisada por esos autores. Para cubrir los trabajos más recientes y otros no registrados por dichas obras, fueron consultadas diversas bibliotecas y archivos especializados en paleontología y arqueozoología del Cuaternario de México, además de consultar en su totalidad varias revistas que usualmente publican información paleontológica de México (Revista de la Sociedad Mexicana de Historia Natural, Revista de la Sociedad Mexicana de Paleontología, Revista Mexicana de Ciencias Geológicas [antes Revista del Instituto de Geología, UNAM], Journal of Paleontology, Journal of Vertebrate Paleontology) y los resúmenes de las reuniones de la Society of Vertebrate Paleontology y de la Sociedad Mexicana de Paleontología. Las tesis revisadas fueron seleccionadas de las bases de datos que mantienen diversas instituciones académicas (e.g., Universidad Nacional Autónoma de México, Universidad Autónoma Metropolitana e Instituto Politécnico Nacional). La información así recopilada se analizó y se actualizaron los datos de naturaleza taxonómica, geológica y temporal.
Las localidades fueron ubicadas de acuerdo con las provincias morfotectónicas de Ferrusquía-Villafranca (1993). Las redes hidrológicas fueron redibujadas de Espinosa-Pérez, Gaspar-Dillanes y Fuentes-Mata (1993) y NEODAT (2009). La nomenclatura taxonómica biológica fue modernizada de acuerdo con Miller, Minckley y Norris (2005), reteniendo el uso de Chirostoma y Moxostoma por no haber suficiente evidencia para aceptar su reemplazo por otros nombre genéricos, ya sea para separar a las especies involucradas en otro género (en el caso de Moxostoma: Chen y Mayden, 2012; Clementz, Bart y Hurley, 2012) o para reunirlas en otro (en el caso de Chirostoma: Bloom, Piller, Lyons, Mercado-Silva y Medina-Nava, 2009).
ResultadosA la fecha, en México se tienen registradas 39 localidades con fósiles de 12 familias, y, por lo menos, 26 géneros y 39 especies de peces dulceacuícolas; estos restos proceden de localidades del Mesozoico y del Cenozoico.
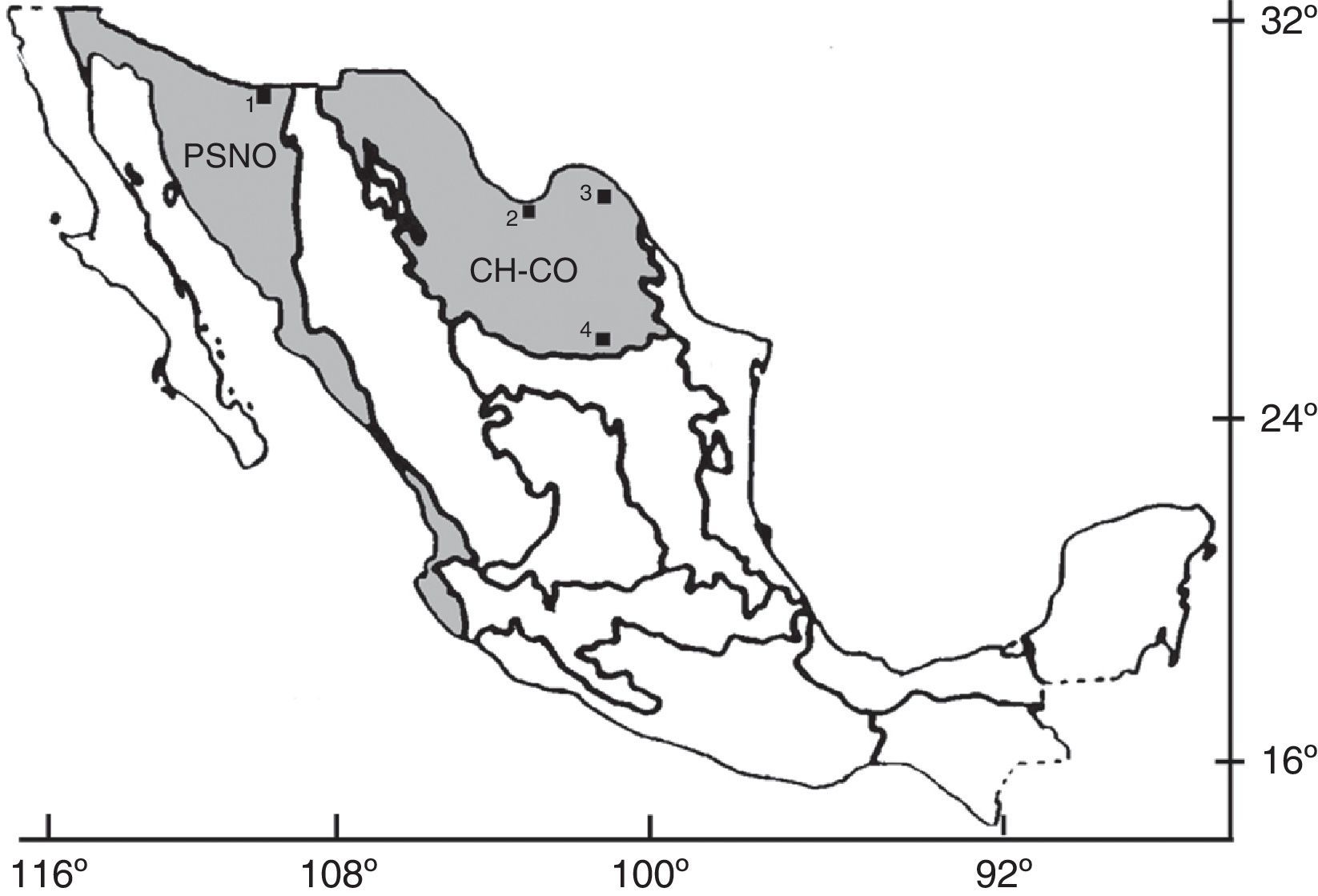
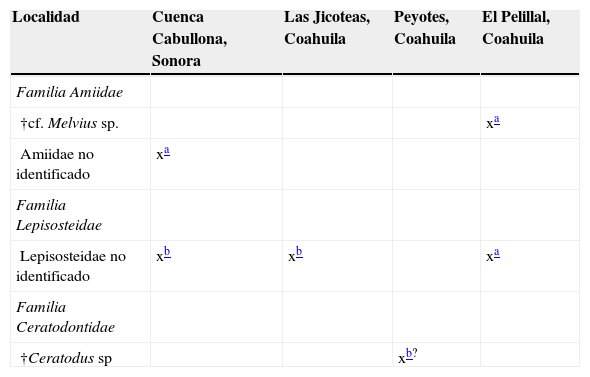
MesozoicoEstas localidades son cretácicas y se encuentran en el norte de México; en ellas se recuperaron restos de 3 familias: Amiidae, Lepisosteidae y Ceratodontidae (fig. 1; tabla 1). La localidad más temprana se ubica en Peyotes, Coahuila, en donde se registró material del pez pulmonado †Ceratodus (Aguilera, 1896, 1907, en Maldonado-Koerdell, 1949), en una caliza apizarrada de temporalidad Turoniana y como fósiles asociados †Inoceramus labiatus, Ostrea lugubris, Anomia sp., †Ptychodus mortoni, †Ptychodus cf. P. whipplei, Trionyx sp. y otros dientes y esqueletos de peces (Böse, 1913; Willis, 1912); posiblemente se trate de un ambiente de transición. En Las Jicoteas, Coahuila (Rivera-Sylva, Frey, Palomino-Sánchez, Guzmán-Gutiérrez y Ortiz-Mendieta, 2009: Formación Aguja, Campaniano) y en la cuenca Cabullona, Sonora (González-León y Lawton, 1995; Lucas, Kues y González-León, 1995: Formación Packard, Cretácico tardío) se recuperaron restos de catán (Lepisosteidae) en depósitos de ambientes transicionales; en este último sitio, en otra formación, se identificó material de un ámido en capas de origen dulceacuícola (González-León y Lawton, 1995; Lucas et al., 1995: Formación Corral de Enmedio, Cretácico tardío). Por otra parte, de los depósitos de ambientes de agua dulce de El Pelillal, Coahuila, proceden restos de catán (Lepisosteidae) y de un ámido (cf. †Melvius sp.) (Rodríguez-de la Rosa y Cevallos-Ferriz, 1998; Formación Cerro del Pueblo, Campaniano). Las especies recientes emparentadas con estos fósiles actualmente habitan ambientes dulceacuícolas y solo algunas especies de catanes habitan esporádicamente aguas salobres y marinas (e.g., Berra, 2007; Grande y Bemis, 1998; Hoese y Moore, 1998; Kells y Carpenter, 2011; Miller et al., 2005).
. Localidades: 1) cuenca Cabullona, Sonora; 2) Las Jicoteas, Coahuila; 3) Peyotes, Coahuila; 4) El Pelillal, Coahuila. Provincias morfotectónicas: PSNO, planicies y sierras del Noroeste; CH-CO, mesetas y cordilleras de Chihuahua y Coahuila (Ferrusquía-Villafranca, 1993).")
Localidades cretácicas con restos de peces fósiles dulceacuícolas. Mapa modificado de Ferrusquía-Villafranca (1993). Localidades: 1) cuenca Cabullona, Sonora; 2) Las Jicoteas, Coahuila; 3) Peyotes, Coahuila; 4) El Pelillal, Coahuila.
Provincias morfotectónicas: PSNO, planicies y sierras del Noroeste; CH-CO, mesetas y cordilleras de Chihuahua y Coahuila (Ferrusquía-Villafranca, 1993).
Ictiofauna dulceacuícola de localidades mesozoicas
Existe un hiato en el registro que abarca casi todo el Paleógeno, para volverse continuo desde el Neógeno al Pleistoceno, con localidades claramente continentales (fluviales y lacustres), con faunas modernas (que aparecen en el Cenozoico) y en sistemas hidrológicos que fueron creándose y evolucionando en el territorio a medida que iba emergiendo, prefigurando muchos de los actuales drenajes.
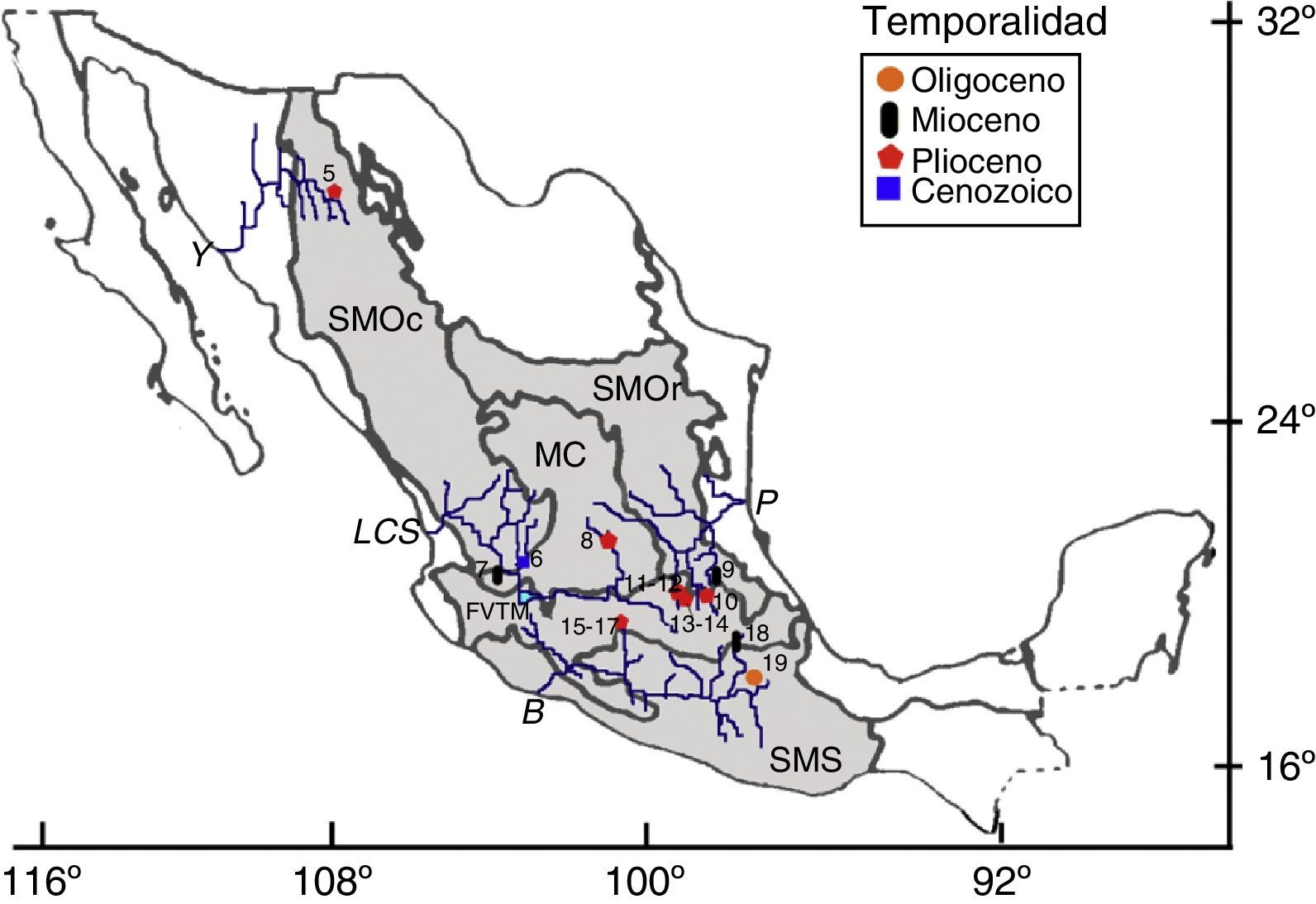
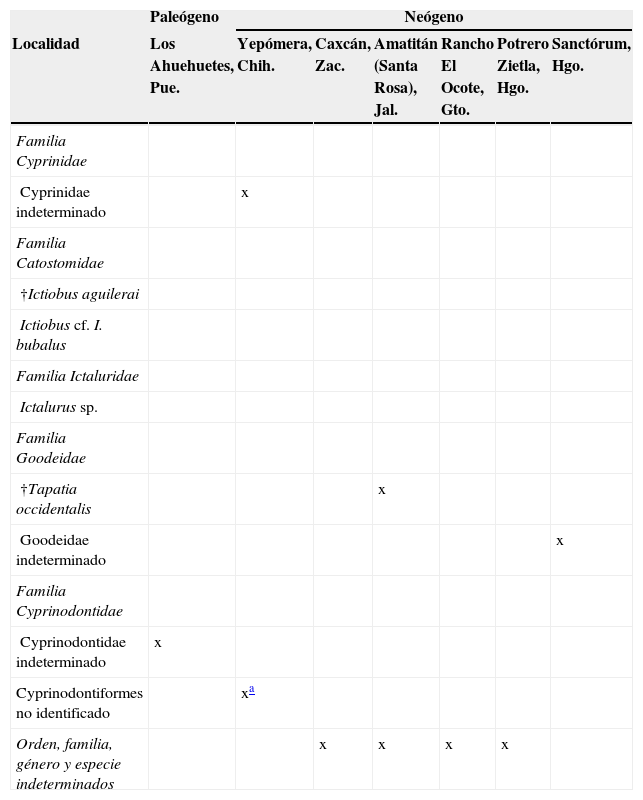
PaleógenoLa localidad Los Ahuehuetes está ubicada en la subcuenca del río Atoyac, dentro de la provincia Sierra Madre del Sur (fig. 2), y tiene una edad oligocénica (Beraldi-Campesi, Cevallos-Ferriz, Centeno-García, Arenas-Abad y Fernández, 2006). Los restos de peces fueron identificados como ciprinodóntidos (Espinosa-Pérez, González-Rodríguez y Pleasanis, 1991; tabla 2), aunque esa familia no se conoce actualmente en ese drenaje (Miller et al., 2005); son peces de origen marino (Ross y Matthews, 2014) y por ello también se les denomina vicarios.
. Localidades: 5) Yepómera, Chihuahua; 6) Caxcán, Zacatecas; 7) Amatitán (Santa Rosa), Jalisco; 8) Rancho El Ocote, Guanajuato; 9) Potrero Zietla, Hidalgo; 10) Sanctorum, Hidalgo; 11) La Cementera, Hidalgo; 12) La Viga de Tula, Hidalgo; 13) El Hoyo, Hidalgo; 14) Teacalco, Hidalgo; 15) El Pirul, Michoacán; 16) cañada de los Cuatro Vientos, Michoacán; 17) Las Encinillas, Michoacán; 18) Panotla, Tlaxcala; 19) Los Ahuehuetes, Puebla. Provincias morfotectónicas: SMOc, Sierra Madre Occidental; SMOr, Sierra Madre Oriental; MC, Meseta Central; Faja Volcánica Transmexicana; SMS, Sierra Madre del Sur (Ferrusquía-Villafranca, 1993). Drenajes: Y, río Yaqui; LSC, Sistema Lerma-Chapala-Santiago; B, río Balsas; P, Pánuco (Espinosa-Pérez et al., 1993; NEODAT, 2009).")
Localidades paleógenas y neógenas con restos de peces fósiles dulceacuícolas. Mapa modificado de Ferrusquía-Villafranca (1993). Localidades: 5) Yepómera, Chihuahua; 6) Caxcán, Zacatecas; 7) Amatitán (Santa Rosa), Jalisco; 8) Rancho El Ocote, Guanajuato; 9) Potrero Zietla, Hidalgo; 10) Sanctorum, Hidalgo; 11) La Cementera, Hidalgo; 12) La Viga de Tula, Hidalgo; 13) El Hoyo, Hidalgo; 14) Teacalco, Hidalgo; 15) El Pirul, Michoacán; 16) cañada de los Cuatro Vientos, Michoacán; 17) Las Encinillas, Michoacán; 18) Panotla, Tlaxcala; 19) Los Ahuehuetes, Puebla.
Provincias morfotectónicas: SMOc, Sierra Madre Occidental; SMOr, Sierra Madre Oriental; MC, Meseta Central; Faja Volcánica Transmexicana; SMS, Sierra Madre del Sur (Ferrusquía-Villafranca, 1993).
Drenajes: Y, río Yaqui; LSC, Sistema Lerma-Chapala-Santiago; B, río Balsas; P, Pánuco (Espinosa-Pérez et al., 1993; NEODAT, 2009).
Ictiofauna dulceacuícola de localidades paleógenas y neógenas
| Paleógeno | Neógeno | ||||||
|---|---|---|---|---|---|---|---|
| Localidad | Los Ahuehuetes, Pue. | Yepómera, Chih. | Caxcán, Zac. | Amatitán (Santa Rosa), Jal. | Rancho El Ocote, Gto. | Potrero Zietla, Hgo. | Sanctórum, Hgo. |
| Familia Cyprinidae | |||||||
| Cyprinidae indeterminado | x | ||||||
| Familia Catostomidae | |||||||
| †Ictiobus aguilerai | |||||||
| Ictiobus cf. I. bubalus | |||||||
| Familia Ictaluridae | |||||||
| Ictalurus sp. | |||||||
| Familia Goodeidae | |||||||
| †Tapatia occidentalis | x | ||||||
| Goodeidae indeterminado | x | ||||||
| Familia Cyprinodontidae | |||||||
| Cyprinodontidae indeterminado | x | ||||||
| Cyprinodontiformes no identificado | xa | ||||||
| Orden, familia, género y especie indeterminados | x | x | x | x | |||
| Neógeno | ||||||||
|---|---|---|---|---|---|---|---|---|
| Localidad | La Cementera, Hgo. | La Viga de Tula, Hgo. | El Hoyo, Hgo. | Teacalco, Hgo. | El Pirul, Mich. | Cañada de los Cuatro Vientos, Mich. | Las Encinillas, Mich. | Panotla, Tlax. |
| Familia Cyprinidae | ||||||||
| Cyprinidae indeterminado | ||||||||
| Familia Catostomidae | ||||||||
| †Ictiobus aguilerai | x | x | x | x | ||||
| Ictiobus cf. I. bubalus | x | |||||||
| Familia Ictaluridae | ||||||||
| Ictalurus sp. | x | x | x | |||||
| Familia Goodeidae | ||||||||
| †Tapatia occidentalis | ||||||||
| Goodeidae indeterminado | ||||||||
| Familia Cyprinodontidae | ||||||||
| Cyprinodontidae indeterminado | ||||||||
| Cyprinodontiformes no identificado | x | |||||||
| Orden, familia, género y especie indeterminados | x | x | ||||||
Las localidades miocénicas se ubican en puntos opuestos de la parte media del país, ya que una se encuentra en Jalisco y las otras en Tlaxcala y en Hidalgo (fig. 2). Se registran en conjunto 3 especies, cada una representante de distintas familias (tabla 2). Amatitán, Jalisco, está asociada al sistema Lerma-Chapala-Santiago y a la provincia Mesa Central, y es la localidad tipo del goodeido más antiguo conocido y a la vez extinto (†Tapatia occidentalis: Álvarez y Arriola-Longoria, 1972; Guzmán, Stinnesbeck, Robles-Camacho y Polaco, 1998); este pez muestra un mosaico de estructuras nuevas (e.g., gonopodio) y ancestrales, y estas últimas desaparecen en las formas recientes de la familia, pero se mantienen en otras familias del orden Cyprinodontiformes (e.g., forma del proceso coronoides del ángulo articular) (Guzmán, 2010). Panotla, Tlaxcala, es una localidad de edad miocénica ubicada en la parte alta (subcuenca del Atoyac) de la cuenca del Balsas, dentro de la provincia Sierra Madre del Sur (Castañeda-Posadas, Calvillo-Canadell y Cevallos-Ferriz, 2009); ahí se han identificado restos de Ictiobus cf. I. bubalus e Ictalurus sp. (Castañeda-Posadas, 2004). Por otro lado, en Potrero de Zietla, Hidalgo, dentro de la cuenca del río Pánuco y la parte sur de la provincia Sierra Madre Oriental, se conocen restos de un pez indeterminado (Castillo-Cerón, Cabral-Perdomo y Carranza-Castañeda, 1996).
Existen 10 localidades pliocénicas, 9 de las cuales se ubican en la parte media de México (fig. 2), generalmente en las partes altas de las cuencas hidrológicas más extensas del país (Pánuco, Balsas y Lerma-Chapala-Santiago); en ellas se han recuperado restos fósiles de catostómidos, ictalúridos, goodeidos, un ciprinodontiforme y otras formas no identificadas (tabla 2). Del Rancho El Ocote en la Meseta Central, y asociado a la subcuenca del río de La Laja, procede un pequeño conjunto de restos de peces no identificados (Carranza-Castañeda y Ferrusquía-Villafranca, 1978). Hidalgo presenta varias localidades, todas asociadas a la cuenca del río Pánuco, aunque de 2 diferentes provincias y diferente contenido fósil: en Sanctorum, en la provincia Sierra Madre Oriental y en el río Moctezuma, se conservaron, en capas laminares de limolita, restos de goodeidos en relación anatómica, esto es, conservando la posición natural en el esqueleto (Becerra-Martínez, Guzmán y Velasco de León, 2002; Becerra-Martínez, 2003; Rodríguez-Becerra, Velasco de León y Salgado-Ugarte, 2002; Rodríguez-Becerra, 2004); en las otras 4 localidades, ubicadas en la provincia Faja Volcánica Transmexicana y relacionadas al drenaje del río Tula, se recuperaron restos de una especie fósil extinta (†Ictiobus aguileraiAlvarado-Ortega, Carranza-Castañeda y Álvarez-Reyes, 2006) y de bagres del género Ictalurus (Alvarado-Ortega y Carranza-Castañeda, 2002a, 2002b, 2003; Castillo-Cerón et al., 1996). Tres afloramientos más (El Pirul, Cañada de los Cuatro Vientos y Las Encinillas) están asociados a la subcuenca del lago de Cuitzeo del río Lerma, en Michoacán (López-García, 2008), en donde se localizaron algunos peces articulados del orden Cyprinodontiformes.
La única localidad pliocénica más allá del área central del país ya referida es Yepómera, en Chihuahua, ubicada hacia el norte de la provincia Sierra Madre Occidental, en la subcuenca del río Papigóchic de la cuenca del río Yaqui (fig. 2); en este sitio se identificaron restos de ciprínidos y ciprinodontiformes (Smith, 1981; tabla 2). La temporalidad de los materiales de Caxcán, Zacatecas, no ha sido establecida (fig. 2; tabla 2), pero por el tipo de preservación (capas laminares con ejemplares articulados de estirpes modernas), se estimó que corresponden por lo menos al Neógeno. Caxcán se ubica en la subcuenca del río Juchipila al sur de la provincia Sierra Madre Occidental, y los restos todavía no están identificados (R. Guzmán-Gutiérrez, comunicación personal, 2009).
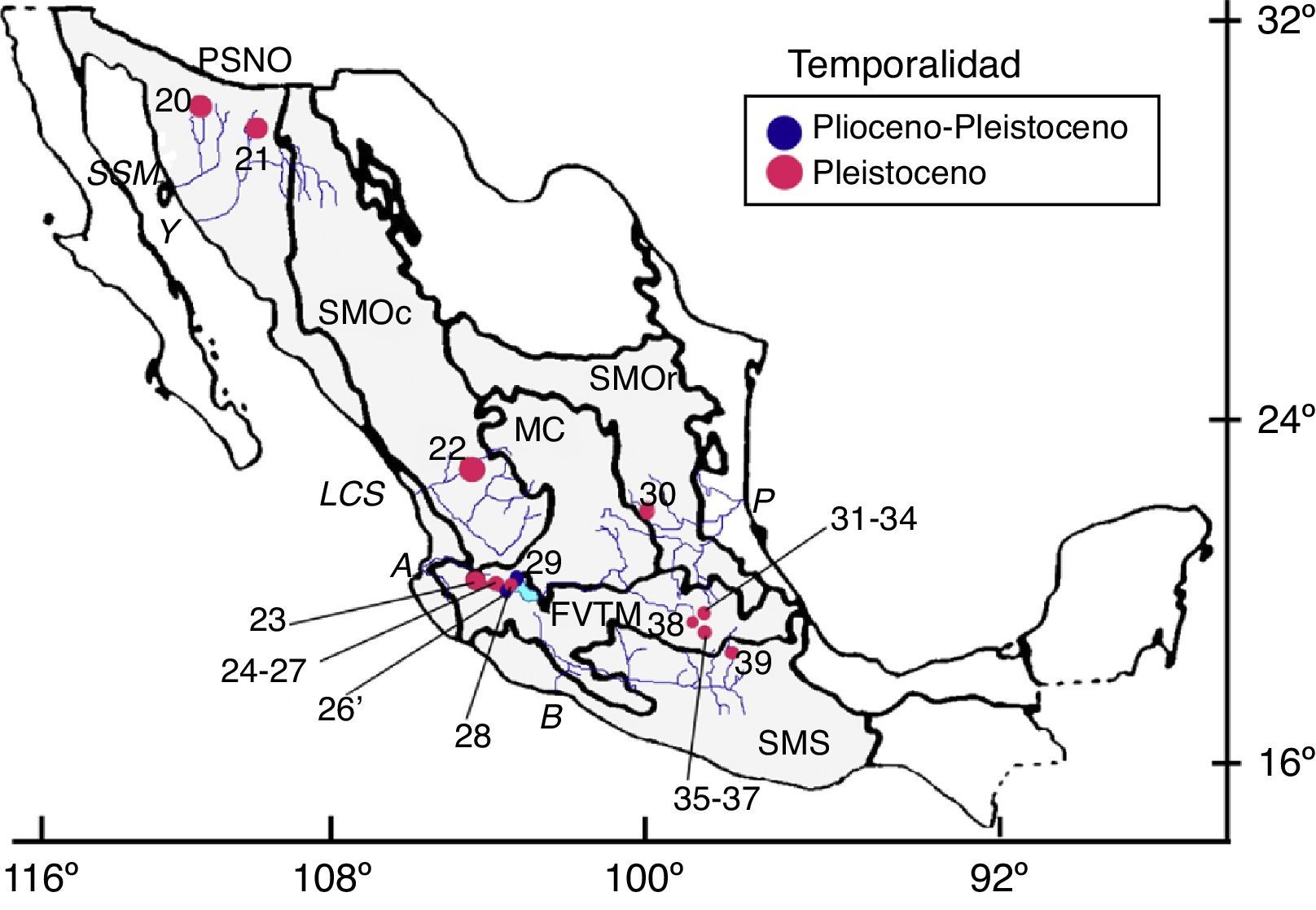
CuaternarioPleistoceno. Es la temporalidad con el mayor número de localidades, provincias morfotectónicas y diversidad registradas (fig. 3; tabla 3; 20 localidades, 6 provincias y 32 especies). Hacia el norte, en Sonora, hay 2 localidades en 2 drenajes diferentes de la provincia Planicies y Sierras del Noroeste. En el río Santo Domingo, en el Rancho La Brisca, se reportaron ciprínidos (Agosia chrysogaster), catostómidos (Catostomus wigginsi) y pecílidos (Poeciliopsis) (Van Devender, Rea y Smith, 1985); en Terapa, junto al río Moctezuma, se identificaron ciprínidos (A. chrysogaster), catostómidos e ictalúridos (Mead et al., 2006). Son faunas que actualmente también se registran en los drenajes mencionados (Miller et al., 2005). Más al sur, en la provincia Sierra Madre Occidental y con relación al río Chapalagana, se registró un ciprínido en Los Tanques, Zacatecas (Guzmán y Polaco, 2000); del otro lado del país, en la provincia Sierra Madre Oriental, se recuperaron restos de un ictalúrido en la Laguna de la Media Luna, San Luis Potosí, la cual pertenece al sistema del río Pánuco (Hernández-Junquera, 1977) (fig. 3; tabla 3).
. Localidades: 20) Rancho La Brisca, Sonora; 21) Terapa, Sonora; 22) Los Tanques, Zacatecas; 23) Ameca, Jalisco; 24) Laguna de Atotonilco El Bajo, Jalisco; 25) Laguna de San Marcos, Jalisco; 26) Laguna de Zacoalco, Jalisco; 26’) lagos de Chapala y Zacoalco, Jalisco; 27) Jocotepec, Jalisco; 28) Ajijic, Jalisco; 29) Formación Chapala, Jalisco; 30) Laguna de la Media Luna, San Luis Potosí; 31) San Miguel Tocuila, Estado de México; 32) Huatepec, Estado de México; 33) Santa Lucía, Estado de México; 34) Santa Isabel Iztapan, Estado de México; 35) Tlapacoya, Estado de México; 36) Chimalhuacán, Estado de México; 37) Santa Martha Acatitla, Estado de México; 38) Reforma, Distrito Federal; 39) Valsequillo, Puebla. Provincias morfotectónicas: PSNO, planicies y sierras del Noroeste; SMOc, Sierra Madre Occidental; SMOr, Sierra Madre Oriental; MC, Meseta Central; FVTM, Faja Volcánica Transmexicana; SMS, Sierra Madre del Sur (Ferrusquía-Villafranca, 1993). Drenajes: SSM, río Sonora-San Miguel; Y, río Yaqui; LSC, sistema Lerma-Chapala-Santiago; A, Ameca; B, río Balsas; P, Pánuco (Espinosa-Pérez et al., 1993; NEODAT, 2009).")
Localidades plio-pleistocénicas y pleistocénicas con restos de peces fósiles dulceacuícolas. Mapa modificado de Ferrusquía-Villafranca (1993). Localidades: 20) Rancho La Brisca, Sonora; 21) Terapa, Sonora; 22) Los Tanques, Zacatecas; 23) Ameca, Jalisco; 24) Laguna de Atotonilco El Bajo, Jalisco; 25) Laguna de San Marcos, Jalisco; 26) Laguna de Zacoalco, Jalisco; 26’) lagos de Chapala y Zacoalco, Jalisco; 27) Jocotepec, Jalisco; 28) Ajijic, Jalisco; 29) Formación Chapala, Jalisco; 30) Laguna de la Media Luna, San Luis Potosí; 31) San Miguel Tocuila, Estado de México; 32) Huatepec, Estado de México; 33) Santa Lucía, Estado de México; 34) Santa Isabel Iztapan, Estado de México; 35) Tlapacoya, Estado de México; 36) Chimalhuacán, Estado de México; 37) Santa Martha Acatitla, Estado de México; 38) Reforma, Distrito Federal; 39) Valsequillo, Puebla.
Provincias morfotectónicas: PSNO, planicies y sierras del Noroeste; SMOc, Sierra Madre Occidental; SMOr, Sierra Madre Oriental; MC, Meseta Central; FVTM, Faja Volcánica Transmexicana; SMS, Sierra Madre del Sur (Ferrusquía-Villafranca, 1993).
Drenajes: SSM, río Sonora-San Miguel; Y, río Yaqui; LSC, sistema Lerma-Chapala-Santiago; A, Ameca; B, río Balsas; P, Pánuco (Espinosa-Pérez et al., 1993; NEODAT, 2009).
Ictiofauna dulceacuícola de localidades plio-pleistocénicas y pleistocénicas
| Localidad | Rancho La Brisca, Son. | Terapa, Son. | Los Tanques, Zac. | Ameca, Jal. | Laguna de Atotonilco el Bajo, Jal. | Laguna de San Marcos, Jal. | Laguna de Zacoalco, Jal. | Lagos de Chapala y Zacoalco, Jal. | Jocotepec, Jal. | Ajijic, Jal. | Formación Chapala, Jal. |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Familia Cyprinidae | |||||||||||
| Agosia chrysogaster | x | x | |||||||||
| Algansea popoche | x | x | |||||||||
| Algansea tincella | x | ||||||||||
| †Algansea sp. nov. 1 (A. milleri) | x | x | |||||||||
| †Algansea sp. nov. 2 (A. paleomexicana) | |||||||||||
| Algansea sp. | |||||||||||
| Aztecula sallaeia | |||||||||||
| Evarra sp. | |||||||||||
| Notropis sp. | |||||||||||
| Yuriria alta | x | ||||||||||
| †Yuriria sp. nov. (Y. elliana) | x | x | x | x | |||||||
| Cyprinidae indeterminado | x | x | x | x | |||||||
| Familia Catostomidae | |||||||||||
| Catostomus wigginsi | x | ||||||||||
| †Moxostoma sp. nov. (M. ammophilum) | x | ||||||||||
| Moxostoma sp. (grupo robustum) | x | ||||||||||
| Catostomidae indeterminado | x | ||||||||||
| Familia Ictaluridae | |||||||||||
| Ictalurus dugesi | x | ||||||||||
| †Ictalurus spodius | x | x | x | x | |||||||
| Ictalurus sp. | |||||||||||
| Ictaluridae indeterminado | x | ||||||||||
| Siluriformesc indeterminado | x | ||||||||||
| Familia Salmonidae | |||||||||||
| †Rhabdofario sp. nov. (R. rugosus) | x | ||||||||||
| †Onchorhynchusdaustralis | x | ||||||||||
| Onchorhynchus sp. | x | ||||||||||
| Salmonidae indeterminado | x | ||||||||||
| Familia Atherinopsidae | |||||||||||
| Chirostoma chapalae | x | ||||||||||
| Chirostoma consocium | x | ||||||||||
| Chirostoma humboldtianum | |||||||||||
| Chirostoma jordani | |||||||||||
| Chirostoma lucius | x | ||||||||||
| Chirostoma cf. C. lucius | x | ||||||||||
| Chirostoma promelas | x | ||||||||||
| Chirostoma cf. C. promelas | x | ||||||||||
| Chirostoma sphyraena | x | ||||||||||
| Chirostoma sp. | x | x | x | ||||||||
| Familia Poeciliidae | |||||||||||
| Poeciliopsis sp. | x | ||||||||||
| Familia Goodeidae | |||||||||||
| Allophorus robustus | x | ||||||||||
| Ameca splendens | x | ||||||||||
| Chapalichthys encaustus | x | ||||||||||
| Girardinichthys viviparus | |||||||||||
| Goodea cf. G. atripinnis | x | ||||||||||
| Xenotoca sp. | x | ||||||||||
| Goodeidae indeterminado | x | x | |||||||||
| Familia Centrarchidae | |||||||||||
| †Micropterus relictus | x | x | x | x | |||||||
| Orden, familia, género y especie indeterminados |
| Localidad | Laguna de la Media Luna, S. L. P. | San Miguel Tocuila, Edo. Méx. | Huatepec, Edo. Méx. | Santa Lucía, Edo. Méx. | Santa Isabel Iztapan, Edo. Mex. | Tlapacoya, Edo. Méx. | Chimalhuacán, Edo. Méx. | Santa Martha Acatitla, Edo. Méx. | Reforma, D. F. | Valsequillo, Pue. |
|---|---|---|---|---|---|---|---|---|---|---|
| Familia Cyprinidae | ||||||||||
| Agosia chrysogaster | ||||||||||
| Algansea popoche | ||||||||||
| Algansea tincella | x | x | ||||||||
| †Algansea sp. nov. 1 (A. milleri) | ||||||||||
| †Algansea sp. nov. 2 (A. paleomexicana) | x | |||||||||
| Algansea sp. | x | x | ||||||||
| Aztecula sallaeia | x | |||||||||
| Evarra sp. | x | x | ||||||||
| Notropis sp. | x | |||||||||
| Yuriria alta | ||||||||||
| †Yuriria sp. nov. (Y. elliana) | ||||||||||
| Cyprinidae indeterminado | x | x | xb | |||||||
| Familia Catostomidae | ||||||||||
| Catostomus wigginsi | ||||||||||
| †Moxostoma sp. nov. (M. ammophilum) | ||||||||||
| Moxostoma sp. (grupo robustum) | ||||||||||
| Catostomidae indeterminado | ||||||||||
| Familia Ictaluridae | ||||||||||
| Ictalurus dugesi | ||||||||||
| †Ictalurus spodius | ||||||||||
| Ictalurus sp. | ||||||||||
| Ictaluridae indeterminado | x | |||||||||
| Siluriformesc indeterminado | ||||||||||
| Familia Salmonidae | ||||||||||
| †Rhabdofario sp. nov. (R. rugosus) | ||||||||||
| †Onchorhynchusdaustralis | ||||||||||
| Onchorhynchus sp. | ||||||||||
| Salmonidae indeterminado | ||||||||||
| Familia Atherinopsidae | ||||||||||
| Chirostoma chapalae | ||||||||||
| Chirostoma consocium | ||||||||||
| Chirostoma humboldtianum | xe | x | ||||||||
| Chirostoma jordani | x | |||||||||
| Chirostoma lucius | ||||||||||
| Chirostoma cf. C. lucius | ||||||||||
| Chirostoma promelas | ||||||||||
| Chirostoma cf. C. promelas | ||||||||||
| Chirostoma sphyraena | ||||||||||
| Chirostoma sp. | x | |||||||||
| Familia Poeciliidae | ||||||||||
| Poeciliopsis sp. | ||||||||||
| Familia Goodeidae | ||||||||||
| Allophorus robustus | ||||||||||
| Ameca splendens | ||||||||||
| Chapalichthys encaustus | ||||||||||
| Girardinichthys viviparus | x | |||||||||
| Goodea cf. G. atripinnis | ||||||||||
| Xenotoca sp. | ||||||||||
| Goodeidae indeterminado | x | |||||||||
| Familia Centrarchidae | ||||||||||
| †Micropterus relictus | ||||||||||
| Orden, familia, género y especie indeterminados | x | x |
En la Faja Volcánica Transmexicana se pueden reconocer 2 conjuntos de localidades, asociadas a los cuerpos lacustres principales del sistema Lerma-Chapala Santiago (fig. 3). Uno de los conjuntos se presenta al oriente, en la cuenca de México (Álvarez y Moncayo, 1976; Álvarez y Polaco, 1977; Block-Iturriaga, 1963; Bradbury, 1971; Morett-Alatorre y Arroyo-Cabrales, 2001; Niederberger-Betton, 1987; Polaco, 1980, 1987; Valentín-Maldonado, 1987) y el otro hacia el poniente en el área lacustre entre Zacoalco, San Marcos y Chapala (fig. 3; Álvarez, 1966; Álvarez del Villar, 1974; Cavender y Miller, 1982; Smith, Cavender y Miller, 1975; Smith, 1980, 1987) y la cuenca aledaña del río Ameca (Brown, 1912). La fauna identificada en ambas zonas comprende varias especies de aterinópsidos, goodeidos y ciprínidos, más ictalúridos en el conjunto poniente, que en lo general corresponden a familias, géneros y especies que se presentan en 2 regiones, aunque en su momento se consideró que entre los restos de catostómidos y ciprínidos podrían haber especies extintas no descritas: †Algansea sp. nov. 1 (A. milleri), †Algansea sp. nov. 2 (A. paleomexicana), †Yuriria sp. nov. (Y. elliana); †Moxostoma sp. nov. (M. ammophilum). En la zona Zacoalco-San Marcos-Chapala también se identificaron restos de especies extintas de las familias Salmonidae y Centrarchidae (†Rhabdofario sp. nov., †Onchorhynchus australis y †Micropterus relictus), familias cuyos actuales límites de distribución sureños están al norte o al poniente del Sistema Lerma-Chapala-Santiago (Miller y Smith, 1986); también se describió una especie extinta de bagre (†Ictalurus spodiusSmith, 1987), aunque otros investigadores previamente habían considerado que el lago solo ha sido habitado por una especie, la actual Ictalurus dugesi (Álvarez, 1966). El último sitio, Valsequillo, se encuentra en el estado de Puebla en la subcuenca del río Atoyac y en la provincia Sierra Madre del Sur. Ahí se han identificado restos de 3 géneros de ciprínidos: Algansea, Evarra y Notropis (Miller et al., 2005; Smith, 1981), aunque ninguno se distribuye actualmente en ese drenaje (Miller et al., 2005).
DiscusiónComparado con otros vertebrados terrestres mexicanos (Arroyo-Cabrales, Polaco y Johnson, 2007: 770 localidades con 274 especies; Chávez-Galván, Guzmán y Polaco, 2013: 79 localidades con 62 especies), con el número de especies de peces dulceacuícolas mexicanas recientes (Miller et al., 2005: unas 500) o con los peces de agua dulce fósiles de Estados Unidos (Cavender, 1986: 39 familias), el registro fósil de este grupo de peces en México es escaso y muy fragmentario, aunque cuenta con una mejor representación para las temporalidades más recientes que para las más antiguas, tanto en número de sitios como en la biodiversidad. Ello se debe a que los hallazgos de estos restos son generalmente incidentales al estar excavando restos de megafauna o de plantas (Cavender, 1986; Guzmán y Polaco, 2009), pero también, entre otras razones, a la falta de la revisión sistemática de muestras de sedimento en búsqueda de restos de microvertebrados y a la aplicación de técnicas que destruyen la mayoría de los esqueletos orgánicos, como cuando se trabajan columnas para palinomorfos, lo que impide detectar estructuras pequeñas pertenecientes a los peces.
Quizá por esa escasez de datos, en el registro fósil mexicano se observa una separación de las faunas antiguas y las modernas, contrario a lo observado en Sudamérica, en donde hubo copresencia y sustitución (López-Fernández y Albert, 2011). Así, los 3 linajes ancestrales (Lepisosteidae, Ceratodontidae y Amiidae; e.g., Cavender, 1986; Paleobiology Database, 2014), cuyo origen puede considerarse en Pangea (aunque a los ámidos y a los lepisosteidos también se les clasifica como elementos laurasianos: Ross y Matthews, 2014), están presentes únicamente en las localidades del Cretácico y ausentes de los sitios cenozoicos; por su parte, las formas modernas tienen su registro únicamente en el Cenozoico. La ausencia de información para el Paleógeno y para el área norte de México no permite observar para el caso de las familias Amiidae y Ceratodontidae (sensu lato), ahora extirpadas del país, si su reemplazamiento fue gradual o repentino.
Los fósiles más antiguos de los ámidos están en depósitos de ambientes tanto de agua dulce como marinos, pero las formas más tardías del Cretácico tardío se consideran dulceacuícolas, lo cual incluye al género Melvius; la mayoría de los registros de esta familia están localizados hacia los antiguos terrenos de Laurasia, con casos fuera de esa zona especialmente en el Cretácico tardío (Burr y Bennett, 2014; Grande y Bemis, 1998; Martinelli et al., 2012; O’Leary et al., 2012; Paleobiology Database, 2014). Los lepisosteidos se registran del Cretácico tardío al Cuaternario, principalmente en depósitos de agua dulce, aunque varios fósiles más tempranos del orden Lepisosteiformes proceden de yacimientos marinos-estuarinos; en el Cretácico su distribución abarcó zonas de Gondwana y Laurasia, pero la mayoría de los sitios posteriores están en terrenos laurasianos (Echelle y Grande, 2014; Paleobiology Database, 2014).
En el caso del orden Dipnoi, que agrupa a todos los peces pulmonados, los fósiles más antiguos ocurren en rocas, en su mayoría marinas del Devónico temprano; en el Devónico medio y tardío aparecen principalmente en depósitos de agua dulce (Graham, 1997), y a partir del Carbonífero tardío, prácticamente solo en rocas continentales (Long, 1995). Los registros más antiguos (Triásico) de Ceratodus también abarcan partes de Laurasia y Gondwana, disminuyendo en Laurasia, en especial en el Cretácico, hasta aparecer esporádicamente solo en terrenos gondwánicos (Paleobiology Database, 2014). La presencia del pez pulmonado en el Turoniano de Coahuila podría deberse a la incursión de una forma gondwaniana en esa región de Laurasia, o ser un remanente de las formas laurasianas y, en este caso, representar la localidad más sureña. El material que no ha sido localizado (Alvarado-Ortega, González-Rodríguez, Blanco-Piñón, Espinosa-Arrubarrena y Ovalles-Damián, 2006) requiere revisarse, ya que el actual género Ceratodus se considera que está conformado por un conjunto de formas con morfología craneal y dental dispares que han llevado a subdividir al género (Pardo, Huttenlocker, Small y Gorman, 2010); de igual forma, se requiere precisar la capa y el ambiente de depósito de este fósil.
También es recomendable revisar los restos de lepisosteidos, pues recientemente se han definido nuevos caracteres en las escamas que han permitido separar a varios miembros en una familia hermana (ver Echelle y Grande, 2014). Asimismo, los estudios geológicos recientes, si bien han permitido conocer mejor la evolución paleogeográfica de México en el Cretácico, los límites de las líneas de costa, los ambientes de transición contiguos y la temporalidad asociada, todavía deben detallarse, ya que hay propuestas ligeramente diferentes sobre la relación continente-mar en ese periodo de tiempo, que involucran las regiones donde se asientan las localidades cretácicas de nuestro interés (e.g., Goldhammer, 1999; Stinnesbeck y Frey, 2014).
En el Mioceno, la presencia de los catostómidos y los ictalúridos, familias de estirpe neártica con presencia en Norteamérica desde edades geológicas previas (Cavender, 1986; Miller y Smith, 1986; Paleobiology Database, 2014), evidencian el arribo y el establecimiento de esos peces en la parte centro-este del país, probablemente vía el río Pánuco, en donde todavía se distribuyen especies de esas familias. Sin embargo, la localidad de Panotla pertenece a otra cuenca hidrológica (Balsas), en la que no se distribuyen los catostómidos modernos. En el Plioceno los catostómidos y los ictalúridos han sido recuperados, asociados al río Tula dentro de la misma cuenca del río Pánuco. La exploración de afloramientos continentales de mayor antigüedad en el área noreste de México, en busca de nuevas localidades y nuevos materiales fósiles, aportaría más datos sobre la ruta de dispersión seguida por estas 2 familias. Por otro lado, el registro más antiguo del género Ictiobus en México es de la misma edad (Mioceno) que el material de Estados Unidos (Cavender, 1986; Paleobiology Database, 2014), lo que sugiere que la aparición de Ictiobus en el continente es más antigua al Mioceno.
En el caso de la familia Goodeidae, en el Mioceno de Amatitán se ubica el registro fósil más antiguo del grupo, pero la evidencia molecular calibrada con la edad de los eventos geológicos sugiere que la subfamilia Goodeinae, a la que pertenecen todas las especies mexicanas, tiene su origen en el Mioceno medio en otra área más al norte, en la parte alta del río Mezquital (Domínguez-Domínguez et al., 2010). El hallazgo de goodeidos en el Plioceno de Sanctorum sugiere que la migración de la familia desde el occidente hacia el oriente de México, a través del río Lerma y de una conexión antigua entre el río Pánuco y el río Lerma, habría ocurrido durante el Plioceno-Mioceno.
Otras familias neárticas evidencian la migración hacia el sur del país por otras rutas. La familia Salmonidae se distribuye actualmente en el noroeste de México, mientras que la familia Centrarchidae se localiza de manera natural en la cuenca del río Bravo o Grande del Norte (Miller y Smith, 1986; Miller et al., 2005). En el Pleistoceno, el límite sureño de ambas llegaba al Lago de Chapala, en donde se registran especies ahora extintas y solo conocidas para esa zona; el arribo de los centrárquidos probablemente sucedió por conexiones con la meseta central, quizá por las mismas corrientes utilizadas por los goodeinos, mientras que los salmónidos lo habrían realizado por esa misma área o por conexiones de la región occidental. La ausencia actual de la familia Salmonidae en Chapala también sugiere cambios en la calidad del agua, temperatura, oxigenación, transparencia y vegetación riparia. La distribución de otras familias también fue ligeramente mayor a la presente, como es el caso del ciprinodóntido de Los Ahuehuetes, Puebla y de los géneros Evarra, Algansea y Notropis, mencionados por Miller et al. (2005) para Valsequillo. El ciprinodóntido es uno de los pocos fósiles conocidos de la familia en el mundo (Paleobiology Database, 2014) y es más antiguo que el material miocénico de Estados Unidos (Cavender, 1986; Ross y Matthews, 2014); asimismo, requiere de revisión para verificar que no corresponda a alguno de los antiguos miembros de los Cyprinodontidae, ahora ubicados en otras familias.
El tema de extinción de especies es difícil de evaluar para los taxones fósiles más antiguos, ya que el nivel de identificación es general (orden, familia, género, cf.), excepto para Ceratodus y Melvius, géneros que están extintos a nivel mundial. El grado de identificación es más preciso en los fósiles del Pleistoceno, ya que la mayoría de los taxones están determinados a nivel de especie. Cabe hacer notar que, para los restos pleistocénicos del área de Zacoalco-Chapala, inicialmente se propuso que correspondían a las especies que ahora se distribuyen ahí (Smith et al., 1975), pero posteriormente esas identificaciones fueron reconsideradas por los mismos investigadores y fueron propuestas nuevas especies de ciprínidos y catostómidos (Smith, 1980), aunque estas últimas carecen de validez, ya que no fueron publicadas formalmente. Una situación semejante se presentó con los restos de Algansea de la cuenca de México, que son más grandes y robustos que los de la actual Algansea tincella, lo cual puede deberse a que pertenecen a una especie diferente o a que, por cambios ecológicos, A. tincella hubiera reducido con el tiempo su talla, ya que los peces actuales tienen menor tamaño (Block-Iturriaga, 1963). Sin embargo, los estudios subsecuentes de restos de otros sitios de la cuenca no proporcionaron evidencia suficiente para suponer la existencia de una especie fósil diferente a A. tincella (e.g., Álvarez y Moncayo, 1976). En estos casos, debido a que la talla, la forma corporal y la merística de los peces son caracteres influidos por factores epigenéticos, como la cantidad de individuos habitando simultáneamente, el tamaño del tirante de agua, la velocidad de la corriente y la temperatura del agua (e.g., Jacquemin, Martin y Pyron, 2013; Yamahira, Lankford Jr. y Conover, 2006; Woods, 2010), se vuelve importante evaluar si las diferencias osteológicas y morfométricas de los fósiles podrían ser consecuencia de la mayor talla alcanzada por los organismos en el pasado, o de un fenómeno de especiación y de posterior extinción; si fuese el primer caso, es posible que disminuya el número de especies pleistocénicas que realmente están extintas.
Si bien en los últimos años ha habido un incremento en el hallazgo de localidades con restos fósiles de peces de agua dulce en México y, asimismo, ha habido inferencias de tipo paleoecológico, paleogeográfico y evolutivo para algunas especies y algunos sitios (e.g., Álvarez y Moncayo, 1976; Block-Iturriaga, 1963; Bradbury, 1971; Guzmán et al., 1998; Smith et al., 1975; Smith, 1980; Van Devender et al., 1985), todavía faltan cubrir más áreas y temporalidades para establecer esa información a escala regional. Profundizar en esos temas desde el punto de vista paleontológico requiere buscar los restos de manera sistemática, no incidental, lo cual puede ser una tarea ardua con resultados confiables, pero probablemente escasos. Entre las regiones a trabajar se encuentran varias áreas asociadas al río Bravo, las cuales actualmente funcionan como cuencas endorreicas, ya que a través de esos drenajes se estableció comunicación e intercambio de la fauna moderna hacia otras cuencas (Smith y Miller, 1986), y prácticamente no existe información paleontológica de esa región. La ictiofauna tropical fósil no está documentada, por lo que también habría que incorporar la búsqueda en esas regiones. También es necesario generar información osteológica de las especies de peces y de su variación morfológica para poder evaluar correctamente las diferencias encontradas, especialmente cuando se trata de especies de amplia distribución y sujetas a diferentes condiciones ambientales.
Finalmente, un aspecto complementario reciente son las nuevas tecnologías que se han desarrollado, tales como el estudio de isótopos estables para determinar paleodietas, paleoclimas, migraciones, historias de vida y fechamientos, así como abordar, desde las herramientas tradicionales, estudios de anillos de crecimiento para conocer la dinámica poblacional; y de tafonomía para entender la formación del fósil y del depósito (e.g., Casselman, 1983; Iglesias-Martín y Soler-Gijón, 1999; Kocsis, Ős, Vennemann, Trueman y Palmer, 2009; Wang et al., 2008). En particular, las técnicas con isótopos estables de C, O y N dependen del colágeno y de otras biomoléculas conservadas en el fósil y que un organismo animal elabora a partir de la ingesta de agua y de alimento y, a su vez, la relación isotópica que en esas moléculas hay de los átomos (δ18O, δ13C y δ15N) depende de las vía de fijación del CO2 atmosférico de los organismos fotosintetizadores, de la temperatura ambiental y del nivel trófico y amplitud del nicho del consumidor; estas técnicas han sido instrumentadas desde hace varias décadas, especialmente a restos subfósiles y recientes, por lo que existe una amplia literatura al respecto tanto de los aspectos teóricos como de su aplicación (e.g., Aurioles-Gamboa, Rodríguez-Pérez, Sánchez-Velasco y Lavín, 2013; Carré, Bentaleb, Fontugne y Lavallée, 2005; Craig, 1954; Clementz, 2012; Davis, 1987; Dawson y Siegwolf, 2007; Flanagan, Ehleringer y Pataki, 2005; Gonfiantini, 1978; Hobson y Wassenaar, 2008; Kennett y Voorhies, 1996; Moreno, Zangrando, Tessone, Castro y Panarello, 2011; Pérez-Crespo, Alva-Valdivia, Arroyo-Cabrales y Polaco, 2010; Shackleton, 1973); su uso hacia los restos fósiles de peces en México, especialmente de los menos antiguos y con mayor probabilidad de conservar biomoléculas, podría complementar información sobre la dinámica de las poblaciones fósiles, el paleoclima al que estuvieron sujetas, las dietas y los hábitos de migración local y regional, tal y como se ha estado obteniendo para la mastofauna fósil (e.g., Pérez-Crespo et al., 2009; Pérez-Crespo, Arroyo-Cabrales et al., 2010). Un ejemplo de ello sería el análisis de los restos de Algansea y de sedimentos de la cuenca de México, que podría dar respuestas sobre las condiciones ecológicas del lago y de los alrededores, sobre su dieta y sobre la fidelidad o no a un área particular del lago.
A los revisores anónimos de este texto, por las observaciones que sirvieron para mejorar el presente estudio. A Gloria Tapia, su apoyo en la edición final de las figuras.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.