


Las palomas y cotorras pueden resultar perjudiciales para cultivos anuales en algunas regiones de Sudamérica. En este estudio analizamos patrones de uso de hábitat de las palomas torcaza (Zenaida auriculata), manchada (Patagioenas maculosa) y picazuró (P.picazuro) y la cotorra (Myiopsitta monachus) durante la primera emergencia del cultivo de soja en Entre Ríos, Argentina. Se estimó el uso y la disponibilidad de hábitats y se calculó la selección mediante intervalos de confianza de Bonferroni. Ninguna de las 4 especies seleccionó el cultivo de soja. La paloma torcaza seleccionó rastrojos de trigo y colza, las palomas picazuró y manchada seleccionaron hábitats con árboles nativos e introducidos; finalmente, la cotorra común seleccionó hábitats con árboles introducidos. Esto reforzaría la hipótesis de que estas especies solo seleccionaron lotes con cotiledones de soja cuando existe escasez de granos en el ambiente. Asumiendo que esta hipótesis sea verdadera, una alternativa para disminuir daños por aves en soja en emergencia sería disuadirlas de utilizar los lotes de soja, ofreciendo granos para atraerlas a otros sitios y evitar que dañen el cultivo. Esta disuasión podría lograrse haciendo coincidir el espacio y la temporalidad de la siembra de soja con la cosecha de cultivos invernales, o bien ofreciendo granos como atrayentes para las aves.
Pigeons and parrots can damage annual crops in some regions of South America. We analyzed bird damage patterns for Eared Doves (Zenaida auriculata), Spotted-winged Pigeon (Patagioenas maculosa), Picazuro Pigeon (P.picazuro), and Monk Parakeet (Myiopsitta monachus), during the emergence first seedtime in soybean crops in Entre Ríos, Argentina. We estimated habitat use and availability. Habitat selection was estimated by means of Bonferroni confidence intervals. None of the 4 species selected soybean fields. Eared Doves selected wheat and canola stubbles. Picazuro Pigeon and Spotted-winged Pigeon showed strong selection for both native and introduced wooded habitats. Finally Monk Parakeet selected habitats with introduced trees. These results reinforce the idea that these species only selected fields with soybean cotyledons when there is a shortage of grain in the environment. An alternative to reduce bird damage in soybean crops first seedtime could be to offer grains in alternative places to attract them away from the soybean fields. Deterrence could be achieved by coordinating spatio-temporality soybean planting with harvest of winter grain crops, or by offering grains attractive to birds.
La intensa transformación de hábitats naturales vinculados a las actividades antrópicas ha modificado la composición y la abundancia de las comunidades de aves en todo el mundo (Foley et al., 2005; Lemoine, Bauer, Peintinger y Böhning-Gaese, 2007; Murphy, 2003; Vitousek, Mooney, Lubchenco y Melillo, 1997). Una de las principales modificaciones de ambientes naturales por actividades antrópicas es el reemplazo de los mismos por cultivos anuales, un factor determinante de la abundancia de numerosas especies de aves (Filloy y Bellocq, 2006; Lemoine et al., 2007; Murphy, 2003; Toral y Figuerola, 2010). Asimismo, las modificaciones de ambientes naturales por cultivos anuales producen cambios en la biodiversidad, frecuentemente con disminución en la misma (Foley et al., 2005; Vitousek et al., 1997).
Las aves responden de manera diferente a la transformación de la tierra y a la alteración del hábitat original (Filloy y Bellocq, 2007). Las consecuencias posibles y contrastantes son, por un lado, especies que son seriamente amenazadas y, por otro, especies que pueden sobrevivir e incluso prosperar en el nuevo entorno (Filloy y Bellocq, 2006, 2007; Gaston, 1994; Vickery et al., 1999). Estas últimas especies, favorecidas por este nuevo paisaje que les proporciona alimento y refugio, pueden, consecuentemente, transformarse en abundantes y perjudiciales para actividades productivas (Bomford y Sinclair, 2002).
En Argentina, la expansión e intensificación de la agricultura ha sido evidente en las últimas décadas, siendo la introducción de soja genéticamente modificada el principal determinante de estos cambios (Aizen, Garibaldi y Dondo, 2009; Gavier-Pizarro et al., 2012; Oesterheld, 2008). Estos procesos de expansión e intensificación agrícola han ocurrido en diversas ecorregiones, incluyendo el pastizal Pampeano (región con más de un siglo de uso agrícola), el Espinal (donde campos con uso históricamente ganadero se han convertido a la producción de cultivos) y, últimamente, en la región Chaqueña (Baldi y Paruelo, 2008; Gasparri y Grau, 2009; Gavier-Pizarro et al., 2012; Grau, Aide y Gasparri, 2005; Paruelo, Guerschman y Verón, 2005; Zak, Cabido y Hodgson, 2004). Algunas especies como palomas y cotorras se han adaptado y beneficiado con este cambio en la estructura del paisaje (Allen y O’Connor, 2000; Bucher y Ranvaud, 2006; Filloy y Bellocq, 2006) y pueden producir daños en cultivos y generar conflictos entre diversos grupos de actores sociales respecto a la mejor manera de disminuir los problemas (Canavelli, Aramburú y Zaccagnini, 2012).
El daño ocasionado en los sistemas de producción agrícola varía según las especies de aves que lo ocasionan y su biología, la región, la estación del año y el periodo de la producción en el que se presenta (Bomford y Sinclair, 2002; Bucher, 1998; Amano, Ushiyama, Fujita y Higuchi, 2007; Casuso, Tarragó, Jiménez, Dardanelli y Pérez, 2016; Dardanelli, Calamari, Canavelli y Zaccagnini, 2011; Dardanelli, Canavelli y Zaccagnini, 2011; Simon, 2008). La paloma manchada (Patagioenas maculosa), la paloma picazuró (P.picazuro), la paloma torcaza (Zenaida auriculata) y la cotorra común (Myiopsitta monachus) son consideradas perjudiciales para cultivos anuales en Argentina y otros países de Sudamérica, como Colombia, Perú, Bolivia, Paraguay, Brasil y Uruguay (Bruggers, Rodriguez y Zaccagnini, 1998; Bruggers y Zaccagnini, 1994; Bucher y Bedano, 1976; Bucher y Ranvaud, 2006; Dardanelli, Calamari et al., 2011; Dardanelli, Canavelli et al., 2011; Robles, Jacobsen, Rasmussen, Otazu y Mandujano, 2003). En Argentina, estas aves pueden afectar cultivos como sorgo, girasol y maíz, principalmente en maduración, aunque también algunos de ellos (soja y girasol) en emergencia (Ranvaud et al., 2001; Ranvaud y Bucher, 2006). En la actualidad, el daño en soja en emergencia se manifiesta como un problema relevante, ya que las aves se alimentan de las semillas y los cotiledones de las plántulas de soja recién emergidas, produciendo daños que pueden llevar a los productores a resembrar parte o la totalidad de los lotes afectados (Cândido, Snak, Castaldelli, Brocardo y Model, 2008, Canavelli, Zuil, Bernardos y Zaccagnini, 2011; Dardanelli, Canavelli et al., 2011; Ranvaud y Bucher, 2006; Ranvaud et al., 2001; Rodríguez, Tiscornia y Olivera, 2011).
Existen estudios sobre algunos aspectos de la ecología de estas aves, como su dinámica poblacional, ecología reproductiva, comportamiento y dieta (Bucher, 1998; Bucher, Martin, Martella y Navarro, 1990; Dardanelli, Calamari et al., 2011; Spreyer y Bucher, 1998), pero otros aspectos claves para su manejo, como la abundancia poblacional y el uso de hábitat, no han sido plenamente investigados. El objetivo de este trabajo fue analizar patrones de uso de hábitat de las 4 especies de aves granívoras previamente mencionadas (paloma manchada, paloma picazuró, paloma torcaza y cotorra común) en Entre Ríos durante el periodo de emergencia del cultivo de soja, que en Argentina se conoce como «de primera» (i.e., sembrado sobre rastrojo de cultivos de verano, en ausencia de cultivos durante otoño e invierno). En este periodo, que corresponde a la estación de primavera, en general la oferta de alimento para las especies de aves mencionadas es reducida (Bucher, 1998), debido a que quedan pocas semillas disponibles en los rastrojos de cultivos y también hay una baja densidad de las semillas de especies nativas que estas especies consumen (Calvo-Silvera, 2006; Dardanelli, Calamari et al., 2011). Por esta razón, responder si las aves seleccionan lotes de soja para consumir los cotiledones del cultivo por sobre los demás cultivos y plantas nativas disponibles en el ambiente en el momento en que la soja está emergiendo contribuiría a explicar los daños atribuidos a las mismas y, con base en esta información, orientar la búsqueda de opciones de manejo a escala de paisaje que permitan prevenir o disminuir dichos daños.
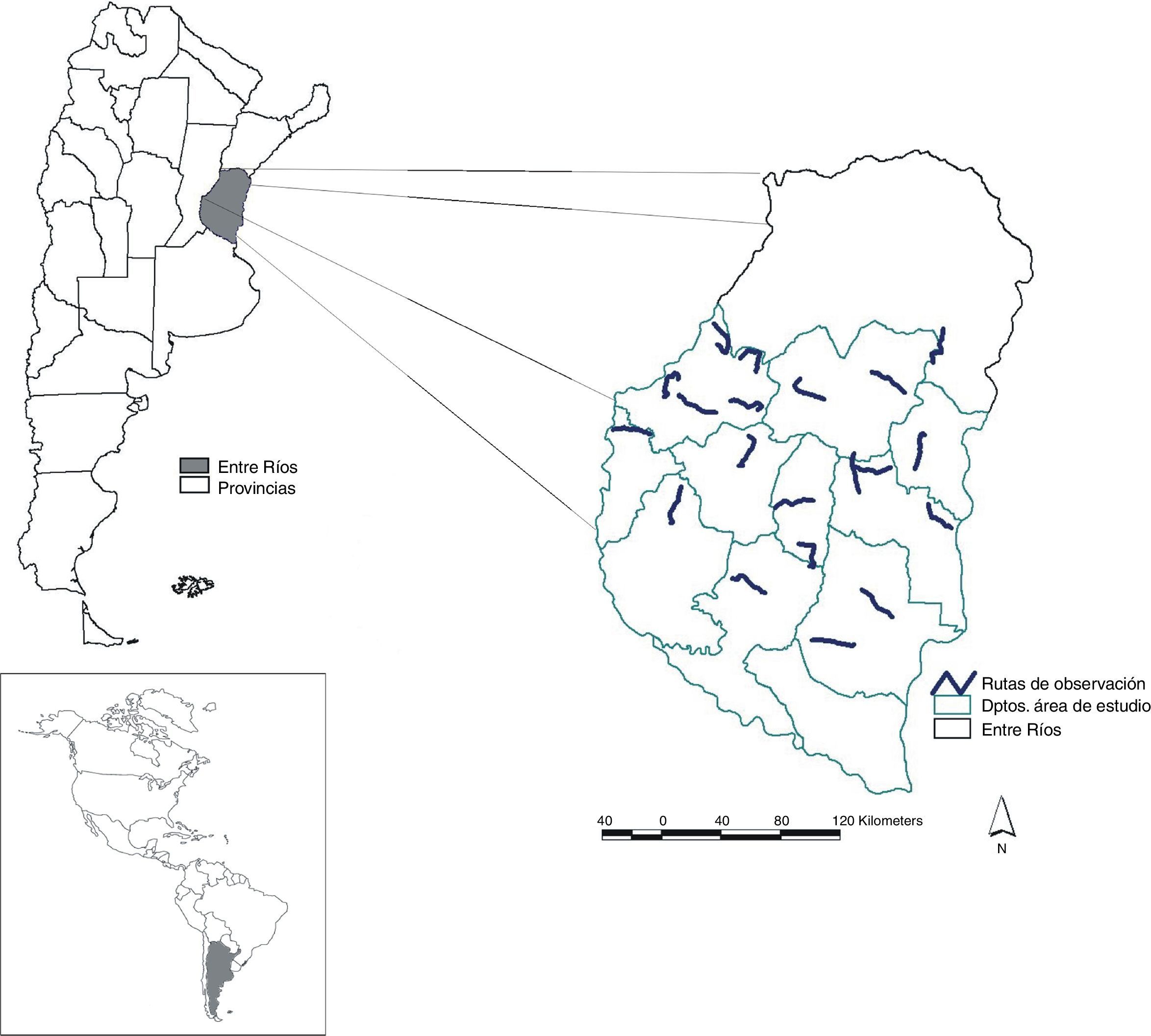
Materiales y métodosEl estudio se realizó en el centro-sur de la provincia de Entre Ríos, Argentina (fig. 1). Originalmente, el área estaba dominada por bosques subtropicales xerófilos bajos y su transición hacia pastizales abiertos, en lo que constituía los bosques subtropicales del Espinal y su ecotono con los pastizales pampeanos (Cabrera, 1976). La fisonomía típica del Espinal está representada por bosques bajos y semiabiertos de tipo xerofíticos, dominados por algarrobos (principalmente Prosopis affinis y P.nigra) (Morello, Matteucci y Rodríguez, 2012). En la actualidad, el área de estudio es una matriz agrícola, con dominancia de cultivo de soja (Glicine max) en verano, algunos campos ganaderos y fragmentos remanentes de bosque del Espinal original (Morello et al., 2012).

El muestreo de aves se realizó en noviembre y diciembre de 2012, coincidiendo con la época de emergencia de soja en el área de estudio. Para ello, utilizando un sistema de información geográfica (ArcGis v 9.2) se ubicó una grilla con celdas de 30×30km sobre el área de estudio, de las cuales se seleccionaron 20 celdas de manera sistemática con arranque aleatorio (Canavelli et al., 2004; Zaccagnini, Canavelli, Calamari y Schrag, 2010). En cada una de estas celdas se seleccionó al azar un camino secundario o terciario como ruta de observación de aves de 30km de largo. En cada ruta se localizaron 30 puntos de conteo de aves equidistantes 1km con el primer punto seleccionado al azar. De este modo se muestrearon en total 600 puntos de conteo distribuidos equitativamente en 2 bloques horarios (06:00-11:00 y 15:00-20:00) de cada día.
En cada punto de observación se registraron todos los individuos observados y oídos de paloma picazuró, paloma manchada, torcaza y cotorra común durante 3min en un radio ilimitado de observación. En cada observación se estimó la abundancia de individuos posados en el suelo, detallando el hábitat que estaban usando.
El uso de la tierra se registró en un radio de 200m alrededor del punto de observación, estimando visualmente los porcentajes de cobertura de las siguientes categorías de uso (Schrag, Zaccagnini, Calamari y Canavelli, 2009): 1)soja, 2)maíz, 3)trigo, 4)colza, 5)otros cultivos y pasturas anuales y perennes (e.g., sorgo, girasol, alfalfa, ray grass), 6)bosque o cortina natural o mixta, 7)bosque o cortina artificial (exótica introducida), 8)pastizales naturales (campos y banquinas con especies no cultivadas de gramíneas y/o dicotiledóneas) y 9)otros (e.g., construcciones, caminos, ambientes acuáticos y otros usos de la tierra). La proporción de cobertura de cada categoría de uso de la tierra a nivel de cada ruta se obtuvo de promediar las proporciones de los 30 puntos (Schrag et al., 2009).
Cada ruta fue considerada como unidad de muestreo. Por este motivo, los valores de abundancia relativa de aves (número de aves observadas/punto) se promediaron para contar con un valor medio y desvíos por ruta (Schrag et al., 2009). A fin de evaluar posibles diferencias en el uso de hábitat entre las especies de aves y entre unidades muestrales se consideró la abundancia de individuos utilizando un hábitat (cobertura/uso de la tierra).
Se determinó si una especie estaba seleccionando, evitando o usando un determinado tipo de cobertura de la tierra de manera indistinta mediante intervalos de confianza de selección de Bonferroni (Neu, Byers y Peek, 1974). Para ello, se interpretó que si el uso observado (abundancia media de aves observadas en un tipo de cobertura, con su correspondiente intervalo de confianza) fue mayor al límite superior del intervalo de confianza de disponibilidad de ese tipo de cobertura (expresado como porcentaje de tipo de cobertura de la tierra), las aves seleccionaron dicha cobertura. Si el uso observado fue menor al límite inferior del intervalo de disponibilidad, entonces ese tipo de cobertura fue evitado por las aves. Adicionalmente, si el valor de abundancia observado estuvo comprendido dentro del intervalo de confianza de disponibilidad de cobertura, entonces las aves usaron ese tipo de cobertura de manera indistinta (Cherry, 1996; Montenegro y Acosta, 2008; Neu et al., 1974). Estas relaciones se estimaron utilizando el programa Havistat V2.2 (Montenegro, Acosta, Reimer, 2014).
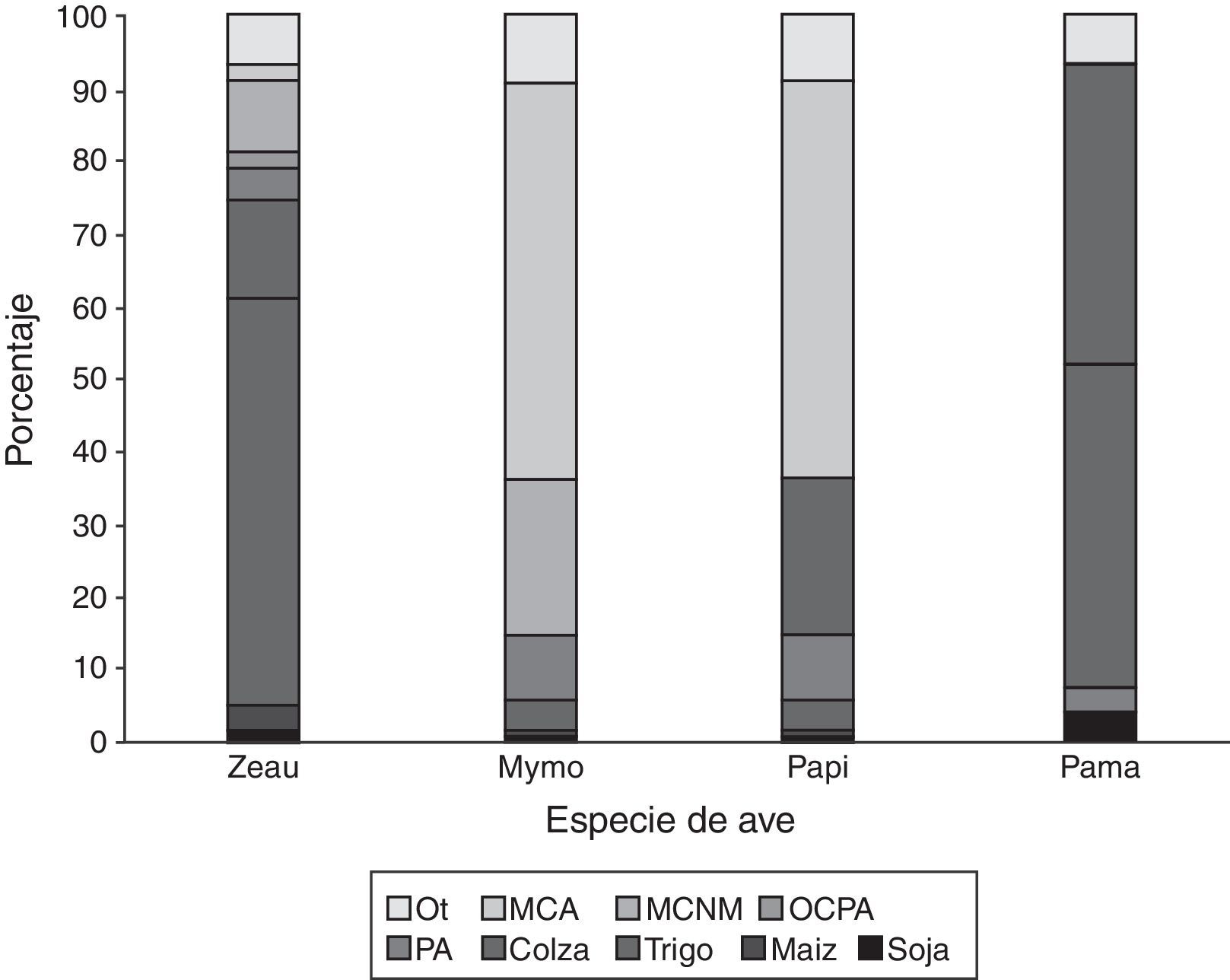
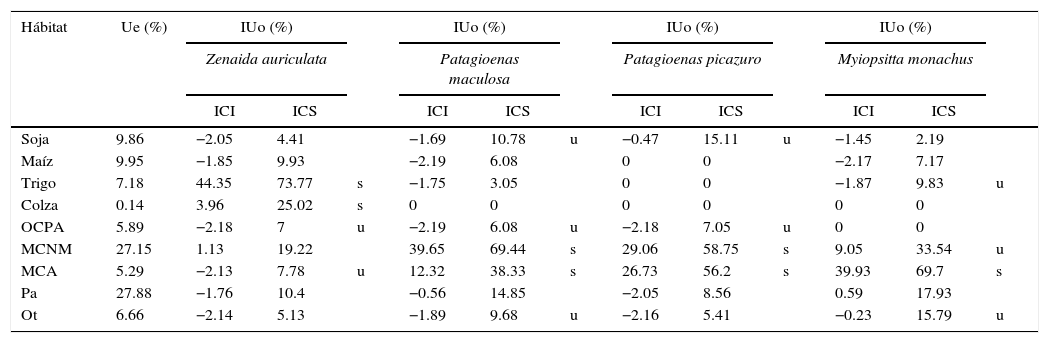
ResultadosLa mayoría (70%) de los individuos de paloma torcaza observados estaban ocupando los rastrojos de cultivos de trigo y colza recientemente cosechados (fig. 2). En cambio, las otras 2 especies de paloma (manchada y picazuró) y la cotorra común mostraron patrones de uso de hábitat diferentes, con la mayoría de los individuos (entre el 76 y el 83%) presentes en bosques nativos y exóticos (fig. 2). Comparando el uso de los distintos tipos de cobertura en relación con la disponibilidad de los mismos, se encontró que la paloma torcaza seleccionó los rastrojos de cultivos de trigo y colza y usó otros cultivos y pasturas anuales en proporción a su disponibilidad (tabla 1). En el caso de las palomas manchada y picazuró, ambas seleccionaron vegetación leñosa, tanto nativa y mixta como exótica, y usaron el cultivo de soja y otros cultivos y pasturas anuales en proporción a su disponibilidad (tabla 1). Finalmente, la cotorra común seleccionó la vegetación leñosa exótica y usó los bosques nativos y mixtos, los rastrojos de trigo y otros tipos de cobertura, como construcciones y caminos, en proporción a su disponibilidad (tabla 1).
 de individuos de cada especie de ave en cada tipo de cobertura. Especies de aves: Mymo: Myiopsitta monachus; Pama: Patagioenas maculosa; Papi: Patagioenas picazuro; Zeau: Zenaida auriculata. Hábitats: MCA: bosque y cortina artificial; MCNM: bosque y cortina natural o mixta; OCPA: otro cultivo o pastura anual o perenne; Ot: otros (e.g., construcciones, caminos, ambientes acuáticos y otros usos de la tierra); Pa: pastizales.")
Uso de hábitat medido en porcentaje (%) de individuos de cada especie de ave en cada tipo de cobertura.
Especies de aves: Mymo: Myiopsitta monachus; Pama: Patagioenas maculosa; Papi: Patagioenas picazuro; Zeau: Zenaida auriculata.
Hábitats: MCA: bosque y cortina artificial; MCNM: bosque y cortina natural o mixta; OCPA: otro cultivo o pastura anual o perenne; Ot: otros (e.g., construcciones, caminos, ambientes acuáticos y otros usos de la tierra); Pa: pastizales.
Análisis de uso de hábitat a partir de los intervalos de confianza de Bonferroni (Cherry, 1996), con un nivel de significancia de 0.05.
| Hábitat | Ue (%) | IUo (%) | IUo (%) | IUo (%) | IUo (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Zenaida auriculata | Patagioenas maculosa | Patagioenas picazuro | Myiopsitta monachus | ||||||||||
| ICI | ICS | ICI | ICS | ICI | ICS | ICI | ICS | ||||||
| Soja | 9.86 | −2.05 | 4.41 | −1.69 | 10.78 | u | −0.47 | 15.11 | u | −1.45 | 2.19 | ||
| Maíz | 9.95 | −1.85 | 9.93 | −2.19 | 6.08 | 0 | 0 | −2.17 | 7.17 | ||||
| Trigo | 7.18 | 44.35 | 73.77 | s | −1.75 | 3.05 | 0 | 0 | −1.87 | 9.83 | u | ||
| Colza | 0.14 | 3.96 | 25.02 | s | 0 | 0 | 0 | 0 | 0 | 0 | |||
| OCPA | 5.89 | −2.18 | 7 | u | −2.19 | 6.08 | u | −2.18 | 7.05 | u | 0 | 0 | |
| MCNM | 27.15 | 1.13 | 19.22 | 39.65 | 69.44 | s | 29.06 | 58.75 | s | 9.05 | 33.54 | u | |
| MCA | 5.29 | −2.13 | 7.78 | u | 12.32 | 38.33 | s | 26.73 | 56.2 | s | 39.93 | 69.7 | s |
| Pa | 27.88 | −1.76 | 10.4 | −0.56 | 14.85 | −2.05 | 8.56 | 0.59 | 17.93 | ||||
| Ot | 6.66 | −2.14 | 5.13 | −1.89 | 9.68 | u | −2.16 | 5.41 | −0.23 | 15.79 | u | ||
ICI: intervalo de confianza inferior del uso observado; ICS: intervalo de confianza superior del uso observado; IUo: intervalo de confianza de uso observado; MCA: bosque y cortina artificial; MCNM: bosque y cortina natural o mixta; OCPA: otro cultivo o pastura anual; Ot: otros (i.e., construcciones, caminos, ambientes acuáticos y otros usos de la tierra); Pa: pastizales; s: selecciona (ICI>Ue); u: Usa (ICI<Ue<ICS); Ue: uso esperado.
Ninguna de las 4 especies estudiadas seleccionó los lotes con cultivo de soja en emergencia. La paloma torcaza seleccionó rastrojos de trigo y colza, las palomas picazuró y manchada mostraron marcada selección por hábitats dominados por árboles tanto nativos como introducidos, y finalmente la cotorra común seleccionó hábitats dominados por árboles introducidos.
La selección de la paloma torcaza por hábitats con rastrojos de trigo y colza podría estar relacionada con sus hábitos de alimentación, ya que esta especie prefiere recoger granos secos del suelo, y una buena parte de la dieta de la especie se basa en semillas cultivadas que recoge de los rastrojos (Baptista, Trail, y Horblit, 1997; Cândido et al., 2008; Dardanelli, Calamari et al., 2011; Fontoura y Orsi, 2013; Goodwin, 1983; Murton, Bucher, Nores, Gomez y Reartes, 1974; Ranvaud et al., 2001). Respecto a esta selección, estudios previos sostienen que el enorme crecimiento poblacional que ha experimentado esta especie se debe principalmente a su capacidad para aprovechar la gran disponibilidad de semillas que permanecen en los rastrojos durante los meses en que debería, en condiciones naturales, sufrir mayor mortandad por escasez de alimento (Bucher, 1990; Bucher y Bedano, 1976). En épocas en que las semillas de los rastrojos están ausentes o poco accesibles para las palomas, como por ejemplo durante el verano, las torcazas prefieren los bosques y evitan los cultivos anuales (Canavelli, Zaccagnini y Calamari, 2006), debido probablemente a que esta especie recurre a semillas de plantas nativas ante la escasez de alimento en los cultivos (Bucher, 1998; Calvo-Silvera, 2006; Dardanelli, Calamari et al., 2011).
La selección de las palomas picazuró y manchada de bosques tanto nativos como introducidos ha sido observada anteriormente para la misma área y en verano (Canavelli et al., 2006). En primavera y verano esta selección podría estar relacionada, por un lado, con la selección que hacen estas especies de los árboles como sitios de nidificación y refugio (Baptista et al., 1997; Dardanelli, Calamari et al., 2011). Por otro lado, esta selección podría estar vinculada a la mayor plasticidad que presentan estas palomas en su comportamiento de alimentación, ya que no solo consumen semillas, sino también frutos y brotes de hojas que obtienen tanto del suelo como sobre la vegetación a distintas alturas (Baptista et al., 1997; Dardanelli, Calamari et al., 2011; Goodwin, 1983). No obstante, se destaca que ambas especies usaron los cultivos de soja en proporción a su disponibilidad en el ambiente. Dado que existe información que demuestra que ambas especies consumen soja en emergencia, tanto en etapa de cotiledón como en las etapas vegetativas siguientes (Bruggers et al., 1998; Bucher, 1990; Dardanelli, Calamari et al., 2011), esto podría implicar que las mismas tengan cierta incidencia sobre los daños del cultivo en la emergencia.
La selección de la cotorra común de boques con árboles introducidos tanto en la primavera (este estudio) como en el verano (Canavelli et al., 2006) estaría relacionada con la marcada selección de sitios altos para nidificar, particularmente árboles del género Eucalyptus (Bruggers et al., 1998; Bucher y Aramburú, 2014; Martín y Bucher, 1993; Spreyer y Bucher, 1998). Esta asociación de la cotorra común con los árboles exóticos se daría durante todo el año, dado que utilizan este tipo de vegetación para nidificar en la época reproductiva y como dormideros durante el periodo no reproductivo (Forshaw, 1989; Martín y Bucher, 1993). La baja asociación de la cotorra común con cultivos de grano puede estar relacionado con el periodo en que se realizó el muestreo, dado que muchos de los frutos de los cultivos que esta especie consume, como por ejemplo maíz, sorgo y girasol, no estaban disponibles para ser consumidos. Además, hay que resaltar que, a diferencia de las palomas estudiadas, la cotorra común no dependería de cultivos de granos, ya que su dieta es más amplia e incluye frutas, brotes y semillas silvestres (Spreyer y Bucher, 1998).
El uso y la selección de un hábitat de alimentación, y el consecuente daño ocasionado en un determinado cultivo, varían de acuerdo con las especies de aves, su biología, la región, la estación y en qué periodo de la producción se presentan (Simon, 2008). En este estudio, la emergencia de la soja coincidió con la cosecha de los cultivos de trigo y colza que, de esta manera, estaban disponibles para ser consumidos por las aves. Estos cultivos fueron seleccionados por la paloma torcaza (mucho más abundante que las otras especies) probablemente porque esta especie prefiere alimentarse de semillas en el suelo (Baptista et al., 1997; Calvo-Silvera, 2006; Dardanelli, Calamari et al., 2011; Ranvaud et al., 2001). Esto reforzaría la idea de que esta especie solo recurriría al consumo de cotiledones de soja cuando existe escasez de granos en el ambiente, y brindaría soporte para recomendar estrategias de manejo, como la disuasión (véase más adelante).
Una característica adicional de la biología de las especies que condicionaría el uso de hábitats es la capacidad de desplazamiento diario para buscar fuentes de alimento (Bucher, 1998; Bucher et al., 1990; Canavelli, Branch, Cavallero, González y Zaccagnini, 2014; Martella, Martín, y Navarro, 1998; Spreyer y Bucher, 1998). Mientras que la paloma torcaza puede explorar un gran rango de área desplazándose más de 100km/día (y hasta 500km anualmente) para aprovechar las fuentes de alimento (Bucher, 1990), la cotorra común posee un área de acción diaria de entre 3 y 5km desde sus nidos cuando se está reproduciendo, y hasta 24km fuera de la época reproductiva (Bucher et al., 1990; Spreyer y Bucher, 1998). Esta notable diferencia en sus capacidades de movimiento puede explicar por qué la paloma torcaza parece estar menos condicionada (al menos en la escala del presente estudio) por la presencia de parches de bosque que las demás especies, a pesar de que la misma necesita de esos parches para nidificar y refugiarse (Bucher, 1990; Bucher y Ranvaud, 2006; Murton et al., 1974). De esta manera, la utilización de los distintos hábitats donde se alimentan las especies estudiadas variaría de acuerdo con las adaptaciones de cada una y a la oferta de alimento presente a lo largo del ciclo anual.
Finalmente, es importante resaltar que ante la oferta de otro tipo de alimento, como granos de trigo y colza en los rastrojos, ninguna de las aves estudiadas seleccionó los lotes de soja en emergencia. De esto se desprende que una alternativa de manejo para disminuir los daños por aves en soja en emergencia sería la disuasión (es decir, ofrecer granos a las aves en sitios diferentes a los lotes), para atraerlas a otros sitios y evitar que dañen la soja en emergencia. La disuasión podría lograrse haciendo coincidir espacio-temporalmente la siembra de soja con la cosecha de cultivos invernales como trigo y colza, que actuarían como divergentes (Amano et al., 2007; Bomford y Sinclair, 2002), o bien ofreciendo granos como alimento alternativo para las aves (Amano et al., 2007; Canavelli et al., 2011). De hecho, esta última medida ha sido mencionada como una medida eficaz para alejar a las aves de los lotes de cultivo que se desean protegerse del daño por aves en momentos críticos del cultivo y de esta manera disminuir los posibles daños que pueden llevar en situaciones extremas a la resiembra de los lotes (Amano et al., 2007; Bomford y Sinclair, 2002). Este tipo de manejo no implica ninguna media activa de repelencia o control letal de aves, las cuales podrían tener consecuencias no deseables sobre el resto de las especies silvestres y el medio ambiente, además de implicar costos extra.
Este estudio fue financiado con fondos del Instituto Nacional de Tecnología Agropecuaria (INTA), Argentina. Agradecemos a Ezequiel Acosta y Darío Bersano (EEA Paraná del INTA) por su asistencia en el trabajo de campo.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.