







Existen discrepancias al delimitar y reconocer bióticamente la parte central del continente americano, principalmente el área comprendida entre el sur de México y el norte de América del Sur. La delimitación de esta zona se ve dificultada al observar características propias, producto de los patrones de distribución de las especies y en establecer criterios en los que se base su reconocimiento. Este trabajo presenta diferentes argumentos sobre la existencia de esa región por medio de su conocimiento biogeográfico, con el fin de proponer una “Mesoamérica biótica” que pueda ser reconocida y diferenciada de las zonas adyacentes por gradientes bióticos. Se plantean criterios que deben ser considerados y los problemas que se presentan en el entendimiento de la región.
There are differences in the biotic delimitation and recognition of the central part of the Americas, specifically from southern Mexico to northern South America. The delimitation of this area has been difficult when typical characteristics, product of patterns of distribution of the species, and diagnostic criteria are considered in its identification. This study present different arguments about the existence of a “biotic Mesoamerica” trough the biogeographical knowledge, which can help in its recognition and its differences from neighboring areas by biotic gradients. Some criteria have been considered and the problems present in the understanding of the region have been discussed.
En 1943, Paul Kirchhoff acuñó el término Mesoamérica para referirse a un conjunto de pueblos precolombinos que presentaban caracteres culturales en común; incluso los delimitó geográficamente, al norte coincidiendo con los límites tropicales en México y al sur hasta el occidente de Costa Rica (Kirchhoff, 1943, Fig. 1). Resulta interesante observar que estos límites coinciden de manera aproximada con los límites geográficos de la distribución de muchas especies de plantas y animales (Vivó, 1943), por lo que podemos encontrar, al igual que con las culturas prehispánicas, elementos característicos, compartidos o ausentes en esta región. La similitud entre rasgos culturales y bióticos también se ha discutido en regiones geográficas más amplias como Latinoamérica y el Neotrópico; donde, a pesar de la congruencia geográfica entre ambos, es posible asociar cada concepto con características culturales y bióticas respectivamente, estableciendo definiciones para propósitos diferentes (Simpson, 1964). Aunque resulta complicado admitir que exista una unidad biótica que se encuentre limitada por aspectos culturales, es claro que al igual que con las culturas, existen elementos biológicos que difieren de forma significativa de las zonas adyacentes en las que se ubican (Wallace, 1876). Indudablemente existe un factor común a esta congruencia, ya que tanto los límites de las especies como los de las culturas se han visto influenciados por factores ambientales (de Mendizábal, 1946; Simpson, 1964).
.")
Delimitación geográfica de Mesoamérica propuesta por Kirchhoff (1943).
Recientemente, Winker (2011) opinó sobre el término biogeográfico adecuado para definir la región que se encuentra entre América del Norte y del Sur, haciendo referencia a que el término América Media o América Central (Middle America), que incluye desde el sur de los Estados Unidos hasta el istmo del Darién, Panamá, incluyendo las Antillas, con excepción de Trinidad y Tobago (Baird, 1864-1872: 1) “tiene una mayor exactitud geográfica y biogeográfica en su definición” (Winker 2011: 5). Además, propuso que el término debe ser oficialmente reconocido, ya que se utilizó por primera vez en la literatura científica en 1864 (Baird, 1864–1872: 1). Sin embargo, esta afirmación carece de un contexto histórico y un conocimiento biogeográfico completo de la región que permita discernir si es adecuada o no, pues el término America Media también se ha utilizado en un contexto geopolítico más que biológico (Lasserre, 1976). En la literatura biológica no existe consenso sobre la definición biótica de Mesoamérica, pues para algunos autores incluye todo México y Centroamérica (Flores y Gerez, 1994; Arroyo-Cabrales et al., 2007), para otros del sureste de México hasta Panamá (Navarro et al., 2001; Ford, 2005) o desde la península de Yucatán hasta Panamá (Comisión centroamericana de ambiente y desarrollo [Ccad], 2002; Cavers et al., 2003), lo que provoca un concepto vago y sin el rigor necesario para poder establecer una unidad reconocible. Además, los conceptos bióticos de Mesoamérica y América Media no deben ser considerados como sinónimos, pues se refieren a áreas con diferente extensión geográfica, dado que en todas las definiciones anteriores Mesoamérica se encuentra dentro de América Media excluyendo el sur de los Estados Unidos y las Antillas (Winker, 2011; Baird 1864–1872).
Por estos motivos, el presente trabajo pretende dar a conocer una visión general del panorama biogeográfico de la parte central del continente americano, de manera tal que pueda contarse con la información necesaria para establecer los límites, conocer los factores que han jugado un papel en la historia biótica de la región y sobre todo, que sea posible reconocer la existencia biótica de Mesoamérica, si fuera el caso, aun sin Mesoamérica, parafraseando el trabajo de Wilkerson (2000).
Elementos bióticos y sus afinidades en la regiónVarios trabajos biogeográficos se han centrado en la explicación del origen de la biota en México y Centroamérica. Los trabajos de Savage (1966, 1982) con herpetofauna; Halffter (1961, 1964a, 1976, 1978) con insectos y Delgadillo (1979, 1986) con musgos, presentan elementos en común, lo que demuestra historias biogeográficas compartidas caracterizadas por distintas afinidades. Si bien estas propuestas están basadas en la dispersión para de explicar el origen de la biota, es interesante notar que los 3 autores sugieren la existencia de un elemento mesoamericano, de un origen relativamente reciente, que hace evidente la identificación de una parte de la biota restringida a esta zona. Además, coinciden también en la presencia de un elemento sudamericano que hace innegable la dispersión de elementos del sur hacia el norte. Un elemento destacado es el componente proveniente del norte (norteño antiguo [Savage, 1966; 1982] o paleoamericano/Neártico [Halffter, 1964b, 1976]) con mayores afinidades neárticas y que para algunos autores está dado por patrones de dispersión norte-sur (Savage, 1966, 1982), aunque hay quienes sugieren que se trata de un elemento autóctono del norte de México (Flores, 1991; Rzedowski, 1991a). Los elementos antes mencionados fueron sintetizados por Savage (1966, 1982) como explicación biogeográfica para la herpetofauna del sureste de México y Centroamérica, reconociendo 4 patrones que constituyen la herpetofauna mesoamericana: 1) mesoamericano, básicamente compuesto por géneros endémicos de México y Centroamérica; 2) septentrional antiguo, con géneros extratropicales de Europa y América del Norte; 3) sudamericano, géneros sudamericanos llegados recientemente a Mesoamérica; y 4) septentrional reciente, géneros extratropicales de la zona árida de México y América del Norte. A partir de éstos, los procesos que explican la historia biogeográfica están basados en eventos de dispersión seguidos por vicarianza: dispersión de América del Sur en el Cretácico tardío seguida de aislamiento antes de la formación del istmo de Panamá, dispersión norteamericana antes del Eoceno seguida por la fragmentación de las biotas en Norte y Centroamérica en el Oligoceno, y dispersión reciente desde América del Sur con el cierre del istmo de Panamá en el Plioceno (Savage, 1982). Esta explicación ha tenido influencia en las interpretaciones biogeográficas para la diversificación de la herpetofauna mexicana (Flores, 1991) y algunos elementos han sido considerados como parte importante en el reconocimiento de los patrones biogeográficos actuales con mamíferos (Webb, 2006; Ceballos et al., 2010; Ferrusquía-Villafranca et al., 2010).
El resultado de la mezcla de los elementos bióticos mencionados, aunado a factores geológicos y ambientales, ha producido una elevada riqueza de especies y una alta tasa de diferenciación in situ, dando como resultado una cantidad importante de endemismos (Rzedowski, 1991a; Flores-Villela y Navarro, 1993; Flores y Gerez, 1994; Peterson y Navarro, 2000) que forman parte del elemento que es característico entre América del Norte y del Sur. Se estima que en la región comprendida por México y Centroamérica existen unas 2 500 especies de vertebrados endémicos (Flores y Gerez, 1994), aproximadamente el 64% de las especies de fanerógamas para México y norte de Centroamérica (Rzedowski, 1991b), y grupos como copépodos presentan hasta un 41% de especies endémicas tan solo en la península de Yucatán (Suárez-Morales y Reid, 2003).
Estas altas concentraciones de especies endémicas se encuentran asociadas principalmente con zonas montanas (Gutiérrez, 2009), dominadas por bosques densos con valores altos de humedad (Watson y Peterson, 1999), representadas por bosques de encinos, coníferas, mesófilos (Rzedowski, 1978) y matorrales subalpinos (Gutiérrez, 2009), y en las tierras bajas las áreas dominadas por bosques tropicales caducifolios y subcaducifolios (Rzedowski, 1978; Ríos-Muñoz y Navarro-Sigüenza, 2012). En estos últimos, es posible encontrar que hasta el 60% de las plantas que componen este tipo de vegetación tienen una distribución restringida sólo a México (Lott y Atkinson, 2010), y en el caso de los vertebrados terrestres, más del 50% de las aves de la zona tropical de México y Centroamérica (Ríos-Muñoz y Navarro-Sigüenza, 2012) y una tercera parte de la mastofauna y herpetofauna de México (García, 2006).
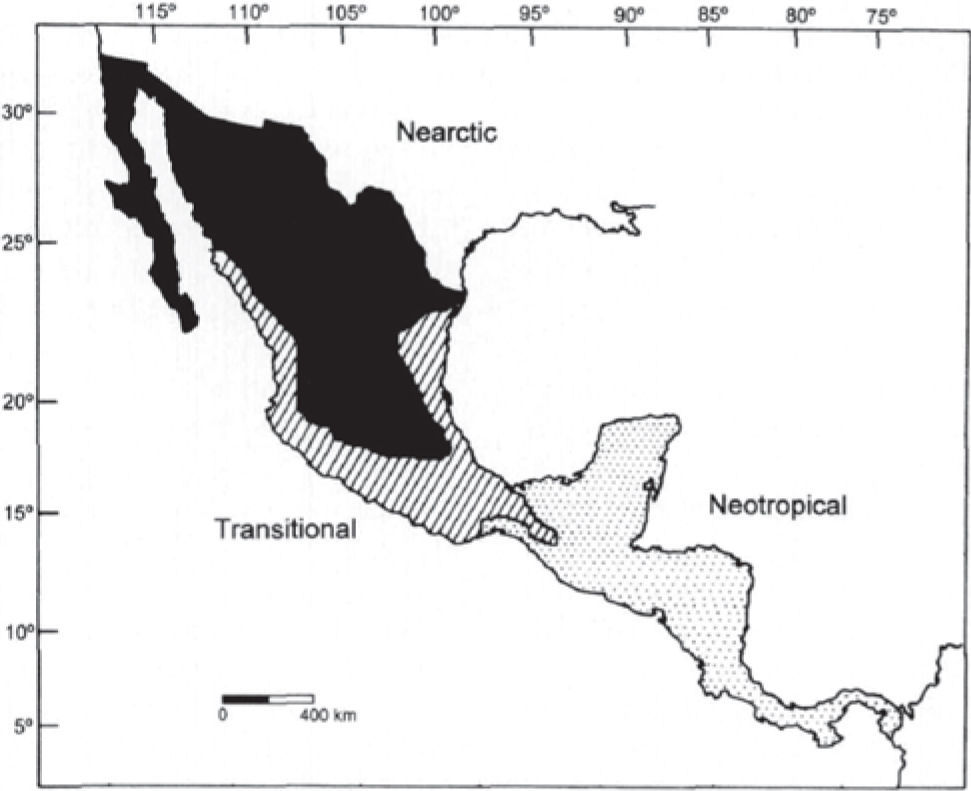
Identificación de la región como zona de transiciónAunque pudiera establecerse el límite latitudinal de los trópicos como la delimitación entre las regiones Neártica y Neotropical en el centro de México, es difícil trazar una única línea, ya que depende de los grupos taxonómicos que se utilicen como forma de evaluación (Escalante et al., 1998). Por esta razón, varios autores han reconocido la existencia de una zona de transición más que un límite, donde es posible encontrar una mezcla de elementos bióticos que pueden pertenecer a ambas regiones (Townsend, 1895; Vivó, 1943; Halffter, 1964a, b, 1976, 1978, 1987; Ortega y Arita, 1998; Morrone, 2004a).
Las zonas de transición se localizan en los límites de las regiones biogeográficas y representan una mezcla de elementos bióticos que es promovida por cambios históricos y ecológicos (Morrone; 2004a). De manera general, se pueden encontrar varias propuestas que cubren los aspectos generales en la descripción de una zona de transición; entre ellas se encuentran las de Vivó (1943), basadas en la distribución de la flora y posteriormente contrastadas con la fauna que presenta la Zona de Transición Mexicana-Centroamericana, que tiene una extensión geográfica coincidente con la Mesoamérica de Kirchhoff (1943) y otra zona de contacto antillana que incluye parte de las Antillas Mayores y el sur de la Florida (Fig. 2). Otra propuesta, la más conocida y difundida, fue realizada por Halffter (1964a, b, 1976, 1978, 1987) llamada Zona de Transición Mexicana (ZTM), que está descrita a partir de la entomofauna, principalmente la asociada a zonas montanas y se extiende desde el suroeste de Estados Unidos hasta Nicaragua, aunque nunca muestra un mapa para apreciar la delimitación completa (Fig. 3). Una tercera, ha sido establecida por Ortega y Arita (1998), quienes con base en la distribución de los murciélagos han establecido que existe una zona transicional que comprende zonas montanas y bajas, extendiéndose desde Sonora y Tamaulipas (en los extremos este y oeste), abarcando la Faja Volcánica Transmexicana (en el centro) hasta las tierras altas de Oaxaca y Chiapas (Fig. 4). La evidencia de que la región comprendida entre América del Norte y del Sur es una zona de transición que ha sidoreconocida también a través de taxones acuáticos como copépodos (Suárez-Morales, 2003; Suárez-Morales et al., 2004a) y peces (Miller, 1966), aunque no establecen una delimitación geográfica. Todas las propuestas coinciden en que se trata de una zona compleja, donde existe una diversidad alta, que es más bien una característica inusual que la identifica (Morrone, 2004a) y geográficamente todas incluyen parte de las zonas tropicales de México y Centroamérica.
, entre las zonas Neártica y Neotropical, con base en su flora.")
Zona de transición propuesta por Vivó (1943), entre las zonas Neártica y Neotropical, con base en su flora.
, con base en la entomofauna y sus sistemas orográficos: I, Sierra Madre Occidental, II, Sierra Madre Oriental; III, Altiplano Mexicano; IV, Eje Volcánico Transmexicano; V, Cuenca del Balsas; VI, Sierra Madre del Sur; VII, Sierra Madre de Chiapas; VIII, Macizo Central de Chiapas; IX y X, Núcleo Centroamericano.")
Zona de Transición Mexicana propuesta por Halffter (1987), con base en la entomofauna y sus sistemas orográficos: I, Sierra Madre Occidental, II, Sierra Madre Oriental; III, Altiplano Mexicano; IV, Eje Volcánico Transmexicano; V, Cuenca del Balsas; VI, Sierra Madre del Sur; VII, Sierra Madre de Chiapas; VIII, Macizo Central de Chiapas; IX y X, Núcleo Centroamericano.
, con base en la distribución de murciélagos.")
Zona de transición propuesta por Ortega y Arita (1998), con base en la distribución de murciélagos.
Reconocimiento del área como unidad biogeográfica. Se han reconocido las diferencias bióticas entre las regiones Neártica y Neotropical y se ha establecido su límite en el centro de México, sur de Florida y las Antillas (Morrone, 2001; Morrone y Escalante, 2009). Algunos autores han identificado el norte del Neotrópico como un área reconocible y la han propuesto como una o varias zonas biogeográficas. Por ejemplo, Wallace (1876) reconoció la región Tropical Norteamericana o región Mexicana (que incluye las vertientes oriental y occidental de México hasta el Darién, Panamá) y una región Antillana (restringida a las Antillas con excepción de Trinidad y Tobago, Fig. 5). Por otro lado, Morrone (2001, 2006) reconoce estos elementos en una subregión Caribeña en la que se encuentran los dominios Mesoamericano, Antillano y del noroeste de América del Sur (Fig. 6), algo muy similar a lo propuesto recientemente por Holt et al. (2013).
, en las que se numeran las subregiones 2, Brasileña; 3, Mexicana y 4, Antillana.")
Extracto de la región Neotropical propuesta por Wallace (1876), en las que se numeran las subregiones 2, Brasileña; 3, Mexicana y 4, Antillana.
 en la región Caribe. Zona de Transición Mexicana: 1, Sierra Madre Occidental; 2, Sierra Madre Oriental; 3, Eje Volcánico Transmexicano; 4, Cuenca del Balsas; 5, Sierra Madre del Sur. Dominio Mesoamericano: 6, Costa Pacífica Mexicana; 7, Golfo de México; 8, Chiapas;, 9, Este de Centroamérica; 10, Oeste del Istmo de Panamá. Dominio Antillano: 11, Península de Yucatán; 12, Bahamas; 13, Cuba; 14, Islas Caimán; 15, Jamaica; 16, Española; 17, Puerto Rico; 18, Antillas Menores. Dominio Noroeste de América del Sur: 19, Chocó; 20, Maracaibo; 21, Costa de Venezuela; 22, Trinidad y Tobago; 23, Magdalena; 24, Llanos venezolanos; 25, Cauca, 26, Islas Galápagos; 27, Occidente de Ecuador; 28. Ecuador árido; 29., Tumbes-Piura (Modificada de Morrone, 2001).")
Provincias biogeográficas propuestas por Morrone (2001, 2006) en la región Caribe. Zona de Transición Mexicana: 1, Sierra Madre Occidental; 2, Sierra Madre Oriental; 3, Eje Volcánico Transmexicano; 4, Cuenca del Balsas; 5, Sierra Madre del Sur. Dominio Mesoamericano: 6, Costa Pacífica Mexicana; 7, Golfo de México; 8, Chiapas;, 9, Este de Centroamérica; 10, Oeste del Istmo de Panamá. Dominio Antillano: 11, Península de Yucatán; 12, Bahamas; 13, Cuba; 14, Islas Caimán; 15, Jamaica; 16, Española; 17, Puerto Rico; 18, Antillas Menores. Dominio Noroeste de América del Sur: 19, Chocó; 20, Maracaibo; 21, Costa de Venezuela; 22, Trinidad y Tobago; 23, Magdalena; 24, Llanos venezolanos; 25, Cauca, 26, Islas Galápagos; 27, Occidente de Ecuador; 28. Ecuador árido; 29., Tumbes-Piura (Modificada de Morrone, 2001).
Es importante señalar que hay quienes piensan que las Antillas no deben ser consideradas del todo como una mezcla de las biotas de América del Norte y del Sur, pues los taxones que ahí habitan han derivado completamente de América del Sur (Savage, 1974) o como zona de transición, no posee las mismas proporciones que se presentan en la parte continental (Vivó, 1943). Sin embargo, existen modelos biogeográficos en donde se ha considerado la composición biótica de las Antillas estrechamente relacionada con la biota centroamericana (Rosen, 1975), e incluso con el norte de América del Sur (Croizat, 1976). Esto puede ser relacionado directamente con los procesos geológicos de la formación de las Antillas y Centroamérica, los que sugieren un origen Pacífico de la placa Caribe (Pindell et al., 2006).
Hasta el momento ningún trabajo ha establecido las relaciones entre los elementos que se encuentran al norte del Neotrópico. Sin embargo, es posible identificar patrones diferenciales entre los dominios (sensuMorrone, 2006) de acuerdo con su composición biótica, definiéndolos como elementos independientes estrechamente relacionados (Morrone, 2001).
Debido a que las explicaciones biogeográficas han sido adecuadas únicamente a ciertos grupos taxonómicos, al igual que la descripción de provincias biogeográficas; p. ej. plantas (Rzedowski, 1978), peces (Miller, 1966), herpetofauna (Smith, 1940, 1949), mamíferos (Goldman y Moore, 1945; Ryan 1953), no ha sido posible obtener un patrón general aplicable a la biota de la región. Por lo tanto, el desarrollo de modelos de consenso para el establecimiento de provincias biogeográficas (Stuart, 1964; Cabrera y Willink, 1980; Morrone, 2001, 2002, 2006) y el desarrollo de métodos que identifiquen la homología biogeográfica (Morrone, 2004b), han creado un mejor entendimiento del área intentando establecer sus límites en los que varios trabajos han destacado la complejidad biogeográfica de la zona.
Homología y entendimiento de las relaciones biogeográficasEstablecer jerárquicamente las relaciones entre las áreas de endemismo es parte del reconocimiento de patrones de homología biogeográfica primaria (Morrone, 2004b). De esta forma se han reconocido 5 provincias en la Zona de Transición Mexicana (Sierra Madre Occidental, Sierra Madre Oriental, Eje Volcánico Transmexicano, Cuenca del Balsas y Sierra Madre del Sur), 5 provincias para el dominio Mesoamericano (Costa Pacífica Mexicana, Golfo de México, Chiapas, este de Centroamérica, Oeste del Istmo de Panamá) y la Península de Yucatán, como la única área continental agrupada dentro del dominio Antillano (Morrone, 2001, 2002; Fig. 6). Sin embargo, el reconocimiento de las relaciones entre las áreas está dado por homología secundaria (Morrone, 2004b), la cual se establece a partir de patrones dados por vicarianza, lo que permite reconocer no sólo los componentes bióticos, sino contar con métodos que permitan la reconstrucción de las relaciones entre las áreas de endemismo con base en las filogenias de los taxones que las habitan (Humphries y Parenti, 1999).
Los trabajos con base en este fundamento tuvieron especial interés en la parte norte del Neotrópico (Rosen, 1978; Liebherr, 1991, 1994; Marshall y Liebherr, 2000; Flores-Villela y Goyenechea, 2001), ya que incluso el primer ejemplo del establecimiento de procesos vicariantes y métodos en biogeografía cladística se realizó en esta zona (Rosen, 1978; Platnick, 1981). Además, se reconocieron los patrones mesoamericanos y se describieron las relaciones entre las áreas de distribución. Sin embargo, no existe un consenso entre las relaciones que guardan las diferentes áreas (Morrone, 2001), por lo que se considera que sigue en una etapa descriptiva.
Uno de los problemas han sido las relaciones biogeográficas de la península de Yucatán, la cual se ha relacionado directamente con las Antillas (Morrone, 2001; Suárez-Morales y Reid, 2003) o con la parte continental (Morrone et al., 1999; Marshall y Liebher, 2000; Ramírez-Barahona et al, 2009), aunque no se ha considerado si esta aparente incongruencia es producto de que se estén analizando 2 historias biogeográficas producidas por una diferencia temporal, una asociándola con las Antillas (Suárez-Morales y Reid, 2003) por su relación geológica desde el Paleoceno y una más reciente que se estableció a partir del cierre del istmo Centroamericano a finales del Plioceno.
Otro problema que se presenta es la definición de la región hacia su límite sur. Aunque se ha propuesto que hay una continuidad biogeográfica hasta la región del istmo de Panamá (Morrone, 2001) y no sólo hasta los límites de las zonas de transición propuestas (Vivó, 1943; Halffter, 1987; Ortega y Arita, 1998), no existe un consenso que permita definir un límite como sucede en la parte norte, el cual lo marca la zona tropical (Wallace, 1876; Vivó, 1943; Halffter, 1987; Morrone, 2006).
Hacia el reconocimiento biótico entre México y CentroaméricaLa identificación de los patrones biogeográficos y ecológicos podría ser, sin duda, uno de los criterios para la delimitación geográfica de la región, ya que pueden presentarse límites marcados por gradientes ecológicos que podrían llevar a un reconocimiento biogeográfico incluyente con respecto a la historia y la ecología, debido a que se ha demostrado que no son excluyentes (Kreft y Jetz, 2010). El nombre “Mesoamérica” por sí solo puede resultar carente de contexto biológico. Sin embargo, el definir un concepto a nivel biótico sería útil para denotar una región rica y de una historia biológica compleja, incluso llegando a ser propuesta como una zona importante para la protección de la biodiversidad a nivel mundial (Myers et al., 2000). Por esta razón, es necesario definir una “Mesoamérica biótica” que pueda ser diferenciable de la “Mesoamérica antropológica” y de los distintos criterios utilizados para definirla y delimitarla (Kirchhoff, 1943; Romero-Contreras y Ávila-Ramos, 1999).
Sin duda, para poder llegar a una definición biótica es necesario considerar los aspectos discutidos previamente. Considerar las relaciones bióticas no es tarea sencilla, la alta tasa de dispersión es un proceso innegable dentro de la configuración biótica del área, el cual modificó considerablemente la estructura biótica desde el cierre del istmo Centroamericano (Stehli y Webb, 1985; Webb, 2006) y ha producido diferencias entre la historia geológica y los patrones generales de distribución de la biota actual (Coates y Obando, 1996; Coates, 1997; Morrone, 2001). Además, existen factores temporales que no han sido incluidos en las propuestas biogeográficas que complican el establecimiento de límites biogeográficos, debido a que pueden modificarse con el tiempo (Zunino y Zullini, 2003). También la presencia de especies con áreas de distribución restringida propias de la región (Williams et al., 2010) demuestra procesos de especiación in situ producidos por vicarianza que podrían ser considerados como recientes (Navarro et al., 2001) y que deben ser incluidos.
Considerando todos estos elementos podría reconocerse una “Mesoamérica biótica” en la que no existen límites fijos, sino gradientes de intercambio con zonas aledañas; al norte, en la división Neártico – Neotrópico a nivel continental (Wallace, 1876; Cabrera y Willink, 1980; Morrone, 2001), al este, la península de Yucatán donde sin duda existe una mezcla de elementos tanto continentales como antillanos (Suárez-Morales y Reid, 2003; Morrone, 2001; Vázquez-Miranda et al., 2007; Ramírez-Barahona et al., 2009) y al sur, desde la depresión de Nicaragua hasta el noroeste de América del Sur (Wallace, 1876; Vivó, 1943; Halffter, 1987; Morrone, 2001, 2006; Holt et al., 2013) que incluye las propuestas que han sido planteadas y que podrían ser integradas e interpretadas como consenso de la información biótica que ahí se encuentra. A través del reconocimiento de la región es posible uniformizar criterios y poder concentrar esfuerzos en continuar con el conocimiento en esta importante y amenazada región (Myers et al., 2000), la cual es sin duda, una de las regiones más polémicas y aún poco conocidas del mundo.
A J. J. Morrone, D. V. Espinosa-Martínez y a 2 revisores anónimos por sus importantes comentarios en versiones previas del manuscrito. A O. Flores-Villela, D. V. Espinosa-Martínez y N. Cortés-Rodríguez por su ayuda para localizar parte de la bibliografía utilizada en el presente trabajo. Al Conacyt y al Posgrado en Ciencias Biológicas de la UNAM, por la beca de estudios de posgrado.