




La información sobre la composición taxonómica y el estado de conservación de la selva mediana subperennifolia en el estado de Hidalgo es escasa y descriptiva, y las alteraciones provocadas por las actividades humanas están afectando drásticamente su estructura y fisonomía original. Por estas razones, los objetivos del presente estudio fueron: 1) realizar un inventario de la flora arbórea, 2) determinar de manera cuantitativa la estructura de los distintos estadios sucesionales de la selva mediana subperennifolia. El inventario florístico y la caracterización estructural se realizaron en 9 localidades del municipio de Huautla, en la Huasteca hidalguense. En cada localidad se calculó área basal, frecuencia, densidad de individuos (diámetro normal > 10cm) y valor de importancia relativa de cada especie. Se identificaron 92 especies, 76 géneros y 31 familias de árboles. Las familias Fabaceae, Malvaceae y Moraceae fueron las más representativas por su riqueza específica. Las especies con valor de importancia relativa más alto fueron Adelia barbinervis y Cedrela odorata en estadios tempranos de la sucesión, Bursera simaruba y Guazuma ulmifolia en etapas intermedias, y Brosimum alicastrum en estadios maduros. En pocas localidades se encontró vegetación madura y la riqueza arbórea total fue comparable con la registrada en el mismo tipo de vegetación en otras entidades de México.
Information on the taxonomic composition and conservation status of the semi-evergreen tropical forest in the state of Hidalgo is limited and descriptive, and disturbances caused by human activities are drastically affecting its structure and original physiognomy. For these reasons, the objectives of this study were: 1) to conduct an inventory of the tree flora, and 2) to provide quantitative data on the structure of the different semi-evergreen tropical forest successional stages. The floristic inventory and structural characterization were conducted in 9 localities of Huautla municipality, in the “Huasteca hidalguense”. In each locality, the basal area, frequency, density of individuals (normal diameter > 10cm) and relative importance value of each species were calculated. A total of 92 species, 76 genera and 31 families of trees were identified. The families Fabaceae, Malvaceae and Moraceae were the most representative for their species richness. The species with highest relative importance value were Adelia barbinervis and Cedrela odorata in early stages of succession, Bursera simaruba and Guazuma ulmifolia in intermediate stages and Brosimum alicastrum in mature stages. In few localities, mature vegetation was observed, and total tree species richness was comparable with that recorded in the same type of vegetation in other entities of Mexico.
El análisis de la sucesión continúa siendo uno de los temas centrales en ecología, la comprensión de la dinámica temporal de las comunidades vegetales es crucial para predecir las fluctuaciones en la biodiversidad y en los servicios de los ecosistemas como respuesta a los cambios ambientales (Zhou et al., 2014). El término sucesión ecológica se refiere al proceso de regeneración natural que conlleva cambios temporales en la composición taxonómica, la estructura y las funciones de un ecosistema después de una perturbación (Huston y Smith, 1987; Martínez-Ramos y García-Orth, 2007).
La disponibilidad de propágulos, las condiciones bióticas, abióticas y las características del disturbio (extensión, intensidad y frecuencia), determinan la composición florística o trayectoria de regeneración particular durante la sucesión (Guariguata y Ostertag, 2001; Martínez-Ramos y García-Orth, 2007). En las últimas décadas se ha puesto especial atención en el análisis de la regeneración en las selvas tropicales del planeta, por la alarmante disminución del área que ocupan y por el enorme reservorio de diversidad biológica que representan (Cole, Bhagwat y Willi, 2014; Macario, García, Aguirre y Hernández, 1995).
La sucesión secundaria en las selvas tropicales húmedas es un proceso continuo, en la etapa inicial los factores más relevantes son los que rigen el proceso de colonización, hasta llegar a estadios más avanzados en los que la habilidad competitiva de las especies y su tolerancia a las condiciones son las que tienden a regular los patrones de reemplazo de especies, las relaciones de dominancia y los tipos de asociaciones en un sitio (Guariguata y Ostertag, 2001; Wiegleb y Felinks, 2001). Durante los distintos estadios de la sucesión el número de especies dominantes del dosel cambia, en las selvas jóvenes es dominado por pocas especies, pero la relación se va equilibrando (codominancia) a medida que alcanzan la madurez (Finegan, 1996).
En México, la selva mediana subperennifolia (SMS) se distribuye a lo largo de la vertiente del golfo de México, desde la porción sureste de San Luis Potosí y el norte de Veracruz, hasta los estados de Campeche, Quintana Roo y Tabasco; se extiende, además, hasta porciones del norte de Oaxaca y Chiapas por la vertiente del océano Pacífico (Challenger, 1998). El área de distribución más septentrional de la SMS en el continente americano (22° N) se encuentra en la Huasteca: Hidalgo, San Luis Potosí y Veracruz (Luna, 1997), región que en la actualidad se caracteriza por poseer una alta densidad de población humana; por lo que la SMS se encuentra confinada a terrenos no aptos para la agricultura, en sitios con pendientes escarpadas o de difícil acceso y su cobertura es en forma de parches o pequeños fragmentos, inmersos en una matriz de hábitats perturbados (Pennington y Sarukhán, 1998). De acuerdo con algunos autores, la SMS con su fisionomía primaria o prístina, ha desaparecido de la Huasteca y ha sido sustituida por vegetación secundaria (Inegi, 2005; Puig, 1991; Rzedowski, 1978; Trejo, 1998).
En el Inventario Nacional de Gran Visión (SARH, 1992) se estimó que en 1981 la cobertura de la SMS en el estado de Hidalgo era de 126,950ha de vegetación primaria y 33,300ha de vegetación perturbada. Sin embargo, la situación cambió drásticamente en 1992, cuando se estimó que solo 9,800ha eran de vegetación primaria, debido principalmente a la elevada tasa de deforestación, de más de 10,600ha anuales (Flores y Gerez, 1994).
El estudio de la sucesión vegetal depende en gran medida de la disponibilidad y veracidad de registros históricos, o bien orales, por lo que establecer una cronosecuencia histórica de una comunidad vegetal es complejo cuando existe poca información y en etapas avanzadas de más de 50 años de regeneración, debido a que la estructura y la composición de la vegetación cambian gradualmente y a velocidad variable (Carreón-Santos y Valdez-Hernández, 2014). Sin embargo, las etapas más conspicuas de la sucesión pueden definirse de manera indirecta y confiable con base en la composición de especies y las características estructurales, cuando no existe información sobre la edad de los fragmentos (Arroyo-Mora, Sánchez-Azofeifa, Rivard, Calvo y Janzen, 2005; Chávez-Costa, Granados-Sánchez y López-Ríos, 2000; Puig, 1991; Rzedowski, 1978).
El conocimiento sobre los cambios en la composición florística y estructura de la vegetación durante la sucesión en la SMS del país es descriptivo y escaso (Puig y Lacaze, 2004), en cambio, el grado de perturbación y pérdida de su cobertura es bastante elevado. La caracterización cuantitativa es indispensable para entender los cambios de la vegetación en el espacio y en el tiempo (Williams-Linera, 2002). Por ello, en el presente estudio se plantearon como objetivos: 1) conocer la composición y estructura de la SMS y 2) analizar la trayectoria de regeneración de este tipo de vegetación, tomando como modelo la SMS del municipio de Huautla, Hidalgo, con la intención de que los resultados obtenidos puedan servir de referencia en la elaboración e implementación de programas de manejo y conservación de la SMS de México.
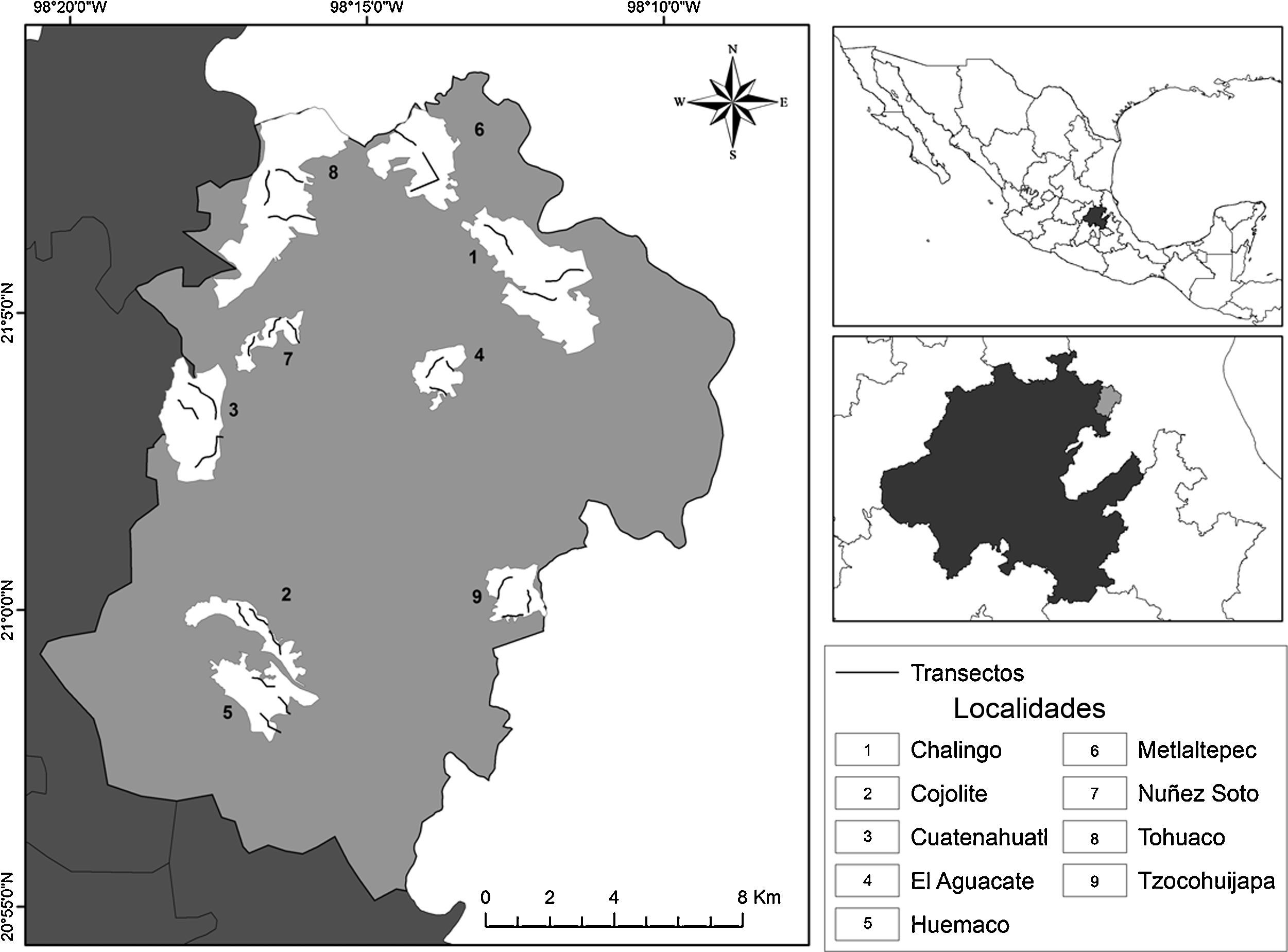
Materiales y métodosEl municipio de Huautla se ubica al noreste del estado de Hidalgo, dentro de la zona ecológica tropical húmeda (Toledo y Ordóñez, 2009), entre los 20°55′ y 21°10′ N, 98°08′ y 98°21′ O (fig. 1); abarca una superficie continental de 292.31km2, lo que equivale al 1.4% de la superficie del estado de Hidalgo. La mayor parte del municipio pertenece a la región fisiográfica de la Sierra Madre Oriental (Huasteca hidalguense) y una pequeña porción a la Llanura Costera del golfo de México (Inegi, 2005).
, estado de Hidalgo (área de color negro), México. Las líneas delgadas que aparecen dentro de cada localidad (áreas de color blanco), representan los transectos en los que se establecieron los 15 puntos de muestreo.")
Localidades de estudio en el municipio de Huautla (área de color gris), estado de Hidalgo (área de color negro), México. Las líneas delgadas que aparecen dentro de cada localidad (áreas de color blanco), representan los transectos en los que se establecieron los 15 puntos de muestreo.
La altitud oscila entre 100 y 800 m, la temperatura entre 22 y 26°C y la precipitación total anual es de 1,400-1,600mm. Se pueden distinguir 3 tipos climáticos dentro del municipio, semicálido húmedo con abundantes lluvias en verano (45% del territorio), semicálido húmedo con lluvias todo el año (42%) y cálido subhúmedo con lluvias en verano (13%) (Inegi, 2005).
La selva mediana subperennifolia ocupa más del 50% de la superficie del municipio (Luna, 1997; Rzedowski, 1978; Trejo, 1998). Sin embargo, en el año 2005 el Inegi estimó que solo 1.77km2 correspondían a SMS preservada y 167.42km2 fueron de vegetación secundaria. La SMS se distribuye en parches de distinto tamaño y estado de sucesión, los fragmentos con vegetación continua más grandes alcanzan tamaños de entre 100 y un poco más de 1,000ha y la mayoría se localizan en las zonas con pendiente más pronunciada. Los fragmentos de menos de 100ha presentan altos nivel de alteración debido al cambio de uso del suelo forestal hacia áreas de cultivo, ganadería y zonas urbanas, principalmente (obs. pers.).
El presente estudio se realizó en 9 fragmentos (localidades) con cobertura arbórea continua mayor a 100ha, con la finalidad de obtener datos representativos de los cambios en la composición, fisionomía, diversidad arbórea y estructura de la SMS en condiciones «prístinas o naturales», en diferentes etapas de la sucesión en la región de la Huasteca hidalguense (límites políticos del municipio de Huautla).
En cada localidad se recolectaron ejemplares de especies de árboles presentes en 3 transectos de 1,500 m de largo cada uno. La recolección se realizó en la temporada de lluvias y de secas de los años 2013 y 2014. Los ejemplares fueron identificados mediante diferentes claves taxonómicas (v. gr. Flora de Veracruz, Flora del Bajío y regiones adyacentes, principalmente; Gentry, 1993; Pennington y Sarukhán, 1998). En el caso del género Ficus se utilizó la clave publicada por Ibarra-Manríquez, Cornejo-Tenorio, González-Castañeda, Piedra-Malagón y Luna (2012). Los ejemplares con estructuras reproductivas en buen estado, se depositaron en la colección del herbario HGOM de la Universidad Autónoma del Estado de Hidalgo.
Cuando la vegetación es escasa o las condiciones del terreno son inaccesibles (paisajes forestales escarpados o perturbados), el uso de parcelas de muestreo para estimar la densidad u otros atributos estructurales no es factible (Hijbeek et al., 2013; Kissaa y Sheilb, 2012). En el presente estudio, las localidades de muestreo presentaron una alta heterogeneidad ambiental: topografía abrupta, presencia de pendientes elevadas y pronunciadas y disturbio elevado, por lo que se eligió la técnica de punto cuadrante central (Cottam y Curtis, 1956) para realizar el muestreo, debido a que no involucra el uso de un área específica (parcelas de muestreo) y considera solo la distancia entre árboles y su altura, para así simplificar el trabajo en campo sin afectar la confiabilidad de los resultados (Forbes y Craig, 2013; Franco et al., 1985; Macario et al., 1995; Mitchell, 2007; Zhu y Zhang, 2009). Esta técnica asume que las especies clave siguen una distribución espacial aleatoria, por lo que no es recomendable cuando la distribución de especies de árboles es uniforme o agregada (Hijbeek et al., 2013; Zhu y Zhang, 2009).
De acuerdo con Korning, Thomsen y Øllgaard (1991), la técnica de punto cuadrante central aplicada a lo largo de transectos proporciona valores confiables de la riqueza de especies, del promedio de la altura y de la densidad de árboles de una región completa, a diferencia de las técnicas de área, que solo reflejan la estructura y composición de la vegetación dentro de cada parcela. Se ha estimado que el número ideal de puntos de muestreo en la TPCC para obtener valores confiables de la densidad de individuos en distintos tipos de vegetación debe ser de al menos 15 (Zhu y Zhang, 2009). Considerando lo anterior, en el presente estudio se seleccionaron al azar 15 puntos diferentes dentro de cada localidad (5 por cada uno de los transectos de 1,500 m, utilizados para la recolección de especies), para medir la altura y el diámetro de los tallos ≥ a 10cm a 1.30cm de altura (diámetro normal [DN]) de todas las especies con forma de vida arbórea. Los puntos de muestreo se colocaron al menos a 100 m de distancia de áreas abiertas, para reducir lo más posible el efecto de borde (Gutiérrez et al., 2009). En los árboles con contrafuertes, el DN se midió 5cm por arriba de esta estructura y en los de tallos múltiples se midieron y sumaron todos los diámetros (Godínez-Ibarra y López-Mata, 2002; Padilla-Velarde, Cuevas-Guzmán, Ibarra-Manríquez y Moreno-Gómez, 2006). En general, se observó que en cada uno de los puntos de muestreo la distribución de las especies más comunes fue aproximadamente al azar. Además, se estimó la completitud del inventario florístico por medio de los índices no paramétricos Chao2 y jackknife (Colwell, 2016) y los valores obtenidos se compararon con los de riqueza de especies en el total de los puntos de muestreo, como una referencia cualitativa de la eficiencia en el muestreo.
La estructura de la vegetación se estimó con base en los valores relativos de área basal (ABR), densidad (DR) y frecuencia (FR) de las especies. El valor de importancia relativa (VIR) de cada especie se calculó con la fórmula VIR = (ABR + DR + FR) / 3 (Matteucci y Colma, 1982). Con los datos de la densidad de las especies obtenidos en las 9 localidades de muestreo, se estimó el índice de Shannon (H’) para las SMS del municipio de Huautla, Hidalgo, con la finalidad de obtener un valor de diversidad susceptible de comparar con el de otras regiones de México con el mismo tipo de vegetación; el cálculo se realizó en el programa PAST 3.0 (Hammer, Harper y Ryan, 2001).
En el presente estudio se utilizó el análisis de agrupamiento (cluster analysis, por sus siglas en inglés), para clasificar a las comunidades vegetales en categorías o grupos, con base en el valor de importancia relativa de las especies. La agrupación fue jerárquica, politética, aglomerativa, utilizando como método de unión el promedio entre grupos y como medida de semejanza el índice de Sorensen. Los análisis se realizaron con el programa de cálculo PC-ORD versión 4.34 (McCune y Mefford, 1999). Para definir los grupos formados en el cluster analysis se eligió un nivel de corte estándar en el que se consideró el 50% de la información remanente (McCune y Grace, 2002).
Además, se realizó un análisis de correspondencia canónica, con datos del VIR de las especies de cada localidad (matriz de datos 1) y con los siguientes valores estructurales promedio: densidad de tallos (árboles/ha), DN, altura del dosel y VIR de las especies pioneras (matriz de datos 2). La elección de las variables estructurales y de las especies pioneras fue con base en conocimiento ecológico previo (v. gr. García-Licona, Esparza-Olguín y Martínez-Romero, 2014; Pennington y Sarukhán, 1998; Puig, 1991; Rzedowski, 1978; Sánchez, Islebe y Hernández, 2007). La dominancia de especies pioneras se consideró como un indicador de disturbio reciente, y por ende de un estadio de sucesión inicial (Arroyo-Rodríguez, Aguirre, Benítez-Malvido y Mandujano, 2007).
La estructura vertical y horizontal de la vegetación en cada localidad se representó por medio de diagramas de perfil semirrealista (Richards, 1952), elaborados con base en la altura promedio de los individuos, los valores estructurales de las especies y las observaciones en campo sobre la fisionomía de la vegetación de las localidades. Se realizó un diagrama de perfil por cada estado sucesional identificado, tomando como referencia la o las localidades más representativas.
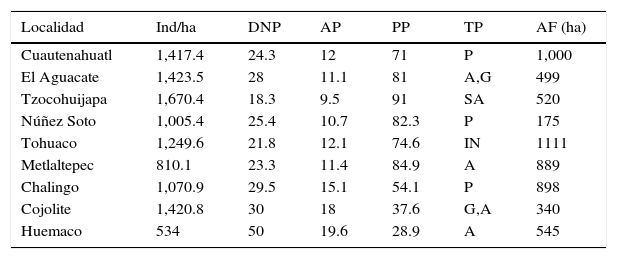
ResultadosSe identificaron 92 especies de árboles (véase Anexo), pertenecientes a 75 géneros y 32 familias. La familia Fabaceae fue la más representativa en número de especies y de individuos, sin embargo, su importancia estructural fue baja en las localidades analizadas. Las familias Malvaceae y Moraceae presentaron alta riqueza de especies, de individuos e importancia estructural. Se identificaron solo 2 especies de Burseraceae, ambas con abundancia e importancia estructural elevada. Las especies con mayor abundancia en la SMS del municipio de Huautla fueron: Adelia barbinervis, Brosimum aliscastrum, Bursera simaruba, Guazuma ulmifolia y Protium copal (tablas 1-4).
Características generales de las 9 localidades de muestreo analizadas.
| Localidad | Ind/ha | DNP | AP | PP | TP | AF (ha) |
|---|---|---|---|---|---|---|
| Cuautenahuatl | 1,417.4 | 24.3 | 12 | 71 | P | 1,000 |
| El Aguacate | 1,423.5 | 28 | 11.1 | 81 | A,G | 499 |
| Tzocohuijapa | 1,670.4 | 18.3 | 9.5 | 91 | SA | 520 |
| Núñez Soto | 1,005.4 | 25.4 | 10.7 | 82.3 | P | 175 |
| Tohuaco | 1,249.6 | 21.8 | 12.1 | 74.6 | IN | 1111 |
| Metlaltepec | 810.1 | 23.3 | 11.4 | 84.9 | A | 889 |
| Chalingo | 1,070.9 | 29.5 | 15.1 | 54.1 | P | 898 |
| Cojolite | 1,420.8 | 30 | 18 | 37.6 | G,A | 340 |
| Huemaco | 534 | 50 | 19.6 | 28.9 | A | 545 |
A: agricultura; AF: área del fragmento; AP: altura promedio; DNP: diámetro normal promedio; G: ganadería; IN: incendio; P: plantaciones (naranja, café o plátano); PP: porcentaje de especies pioneras; SA: sistema agroforestal; TP: tipo de perturbación.
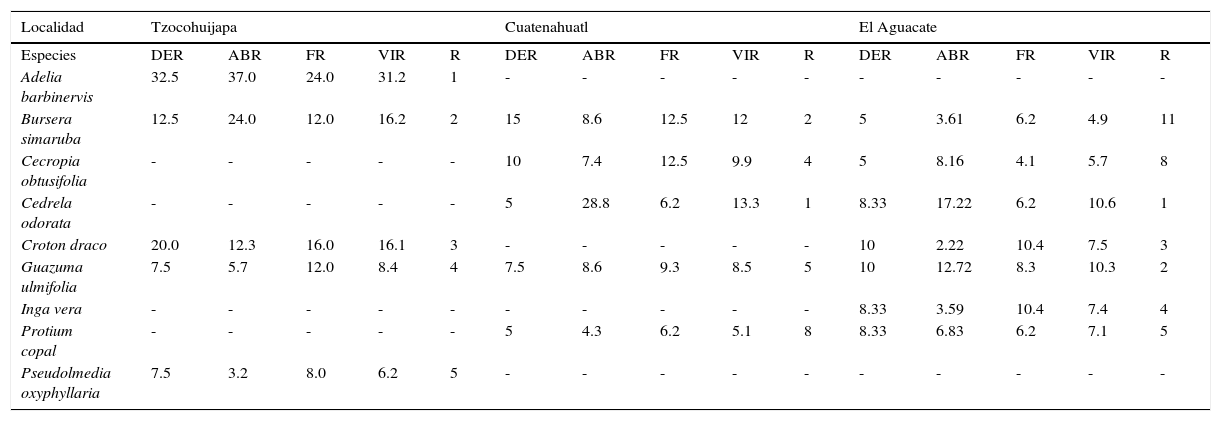
Valores estructurales de las especies de la SMS inicial o juvenil.
| Localidad | Tzocohuijapa | Cuatenahuatl | El Aguacate | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Especies | DER | ABR | FR | VIR | R | DER | ABR | FR | VIR | R | DER | ABR | FR | VIR | R |
| Adelia barbinervis | 32.5 | 37.0 | 24.0 | 31.2 | 1 | - | - | - | - | - | - | - | - | - | - |
| Bursera simaruba | 12.5 | 24.0 | 12.0 | 16.2 | 2 | 15 | 8.6 | 12.5 | 12 | 2 | 5 | 3.61 | 6.2 | 4.9 | 11 |
| Cecropia obtusifolia | - | - | - | - | - | 10 | 7.4 | 12.5 | 9.9 | 4 | 5 | 8.16 | 4.1 | 5.7 | 8 |
| Cedrela odorata | - | - | - | - | - | 5 | 28.8 | 6.2 | 13.3 | 1 | 8.33 | 17.22 | 6.2 | 10.6 | 1 |
| Croton draco | 20.0 | 12.3 | 16.0 | 16.1 | 3 | - | - | - | - | - | 10 | 2.22 | 10.4 | 7.5 | 3 |
| Guazuma ulmifolia | 7.5 | 5.7 | 12.0 | 8.4 | 4 | 7.5 | 8.6 | 9.3 | 8.5 | 5 | 10 | 12.72 | 8.3 | 10.3 | 2 |
| Inga vera | - | - | - | - | - | - | - | - | - | - | 8.33 | 3.59 | 10.4 | 7.4 | 4 |
| Protium copal | - | - | - | - | - | 5 | 4.3 | 6.2 | 5.1 | 8 | 8.33 | 6.83 | 6.2 | 7.1 | 5 |
| Pseudolmedia oxyphyllaria | 7.5 | 3.2 | 8.0 | 6.2 | 5 | - | - | - | - | - | - | - | - | - | - |
ABR: área basal relativa; DER: densidad relativa; FR: frecuencia relativa; R: posición con respecto al VIR; VIR: valor de importancia relativa.
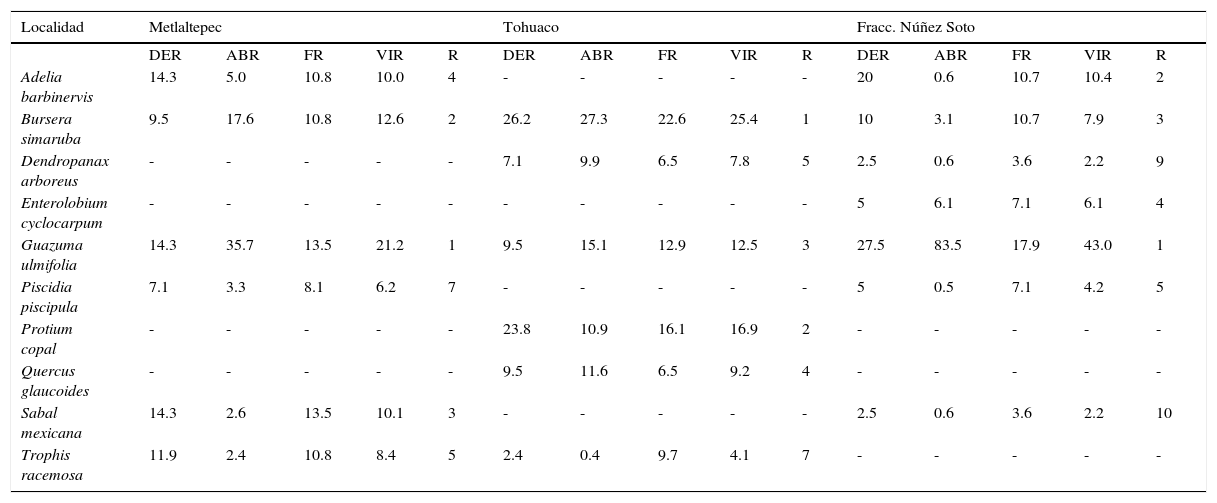
Valores estructurales de las especies de la SMS en estadio intermedio.
| Localidad | Metlaltepec | Tohuaco | Fracc. Núñez Soto | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DER | ABR | FR | VIR | R | DER | ABR | FR | VIR | R | DER | ABR | FR | VIR | R | |
| Adelia barbinervis | 14.3 | 5.0 | 10.8 | 10.0 | 4 | - | - | - | - | - | 20 | 0.6 | 10.7 | 10.4 | 2 |
| Bursera simaruba | 9.5 | 17.6 | 10.8 | 12.6 | 2 | 26.2 | 27.3 | 22.6 | 25.4 | 1 | 10 | 3.1 | 10.7 | 7.9 | 3 |
| Dendropanax arboreus | - | - | - | - | - | 7.1 | 9.9 | 6.5 | 7.8 | 5 | 2.5 | 0.6 | 3.6 | 2.2 | 9 |
| Enterolobium cyclocarpum | - | - | - | - | - | - | - | - | - | - | 5 | 6.1 | 7.1 | 6.1 | 4 |
| Guazuma ulmifolia | 14.3 | 35.7 | 13.5 | 21.2 | 1 | 9.5 | 15.1 | 12.9 | 12.5 | 3 | 27.5 | 83.5 | 17.9 | 43.0 | 1 |
| Piscidia piscipula | 7.1 | 3.3 | 8.1 | 6.2 | 7 | - | - | - | - | - | 5 | 0.5 | 7.1 | 4.2 | 5 |
| Protium copal | - | - | - | - | - | 23.8 | 10.9 | 16.1 | 16.9 | 2 | - | - | - | - | - |
| Quercus glaucoides | - | - | - | - | - | 9.5 | 11.6 | 6.5 | 9.2 | 4 | - | - | - | - | - |
| Sabal mexicana | 14.3 | 2.6 | 13.5 | 10.1 | 3 | - | - | - | - | - | 2.5 | 0.6 | 3.6 | 2.2 | 10 |
| Trophis racemosa | 11.9 | 2.4 | 10.8 | 8.4 | 5 | 2.4 | 0.4 | 9.7 | 4.1 | 7 | - | - | - | - | - |
ABR: área basal relativa; DER: densidad relativa; FR: frecuencia relativa; R: posición con respecto al VIR; VIR: valor de importancia relativa.
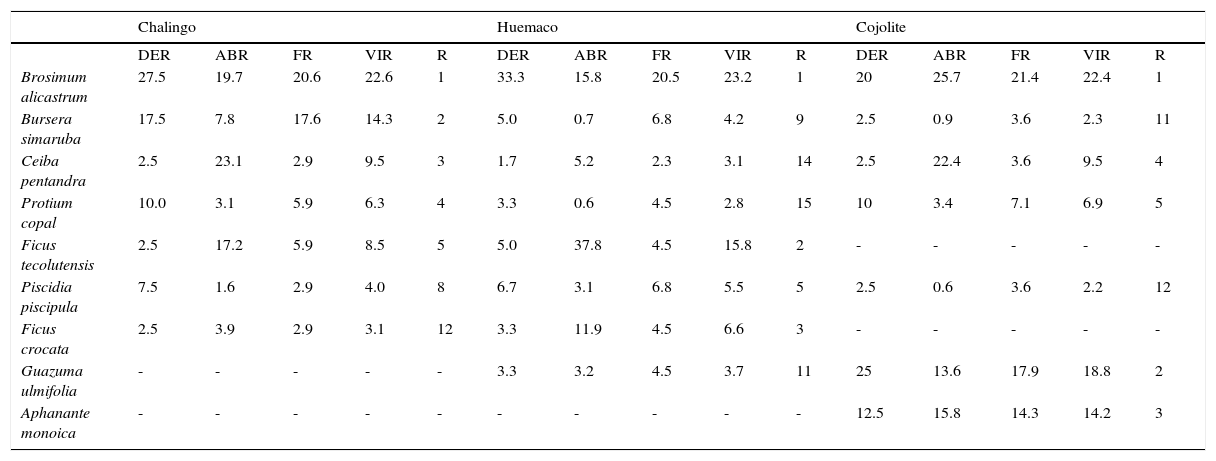
Valores estructurales de las especies de la SMS en estadio maduro.
| Chalingo | Huemaco | Cojolite | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DER | ABR | FR | VIR | R | DER | ABR | FR | VIR | R | DER | ABR | FR | VIR | R | |
| Brosimum alicastrum | 27.5 | 19.7 | 20.6 | 22.6 | 1 | 33.3 | 15.8 | 20.5 | 23.2 | 1 | 20 | 25.7 | 21.4 | 22.4 | 1 |
| Bursera simaruba | 17.5 | 7.8 | 17.6 | 14.3 | 2 | 5.0 | 0.7 | 6.8 | 4.2 | 9 | 2.5 | 0.9 | 3.6 | 2.3 | 11 |
| Ceiba pentandra | 2.5 | 23.1 | 2.9 | 9.5 | 3 | 1.7 | 5.2 | 2.3 | 3.1 | 14 | 2.5 | 22.4 | 3.6 | 9.5 | 4 |
| Protium copal | 10.0 | 3.1 | 5.9 | 6.3 | 4 | 3.3 | 0.6 | 4.5 | 2.8 | 15 | 10 | 3.4 | 7.1 | 6.9 | 5 |
| Ficus tecolutensis | 2.5 | 17.2 | 5.9 | 8.5 | 5 | 5.0 | 37.8 | 4.5 | 15.8 | 2 | - | - | - | - | - |
| Piscidia piscipula | 7.5 | 1.6 | 2.9 | 4.0 | 8 | 6.7 | 3.1 | 6.8 | 5.5 | 5 | 2.5 | 0.6 | 3.6 | 2.2 | 12 |
| Ficus crocata | 2.5 | 3.9 | 2.9 | 3.1 | 12 | 3.3 | 11.9 | 4.5 | 6.6 | 3 | - | - | - | - | - |
| Guazuma ulmifolia | - | - | - | - | - | 3.3 | 3.2 | 4.5 | 3.7 | 11 | 25 | 13.6 | 17.9 | 18.8 | 2 |
| Aphanante monoica | - | - | - | - | - | - | - | - | - | - | 12.5 | 15.8 | 14.3 | 14.2 | 3 |
ABR: área basal relativa; DER: densidad relativa; FR: frecuencia relativa; R: posición con respecto al VIR; VIR: valor de importancia relativa.
Las 9 localidades analizadas se agruparon en 3 estadios sucesionales, los cuales se denominaron selva secundaria inicial o juvenil, intermedia y madura, clasificados de acuerdo con su fisionomía y valores estructurales (tabla 1). A continuación se describe detalladamente la composición florística y estructura de la vegetación en cada estadio de la sucesión identificado, y se incluye un diagrama de perfil para visualizar la estratificación horizontal y vertical de los fragmentos de selva.
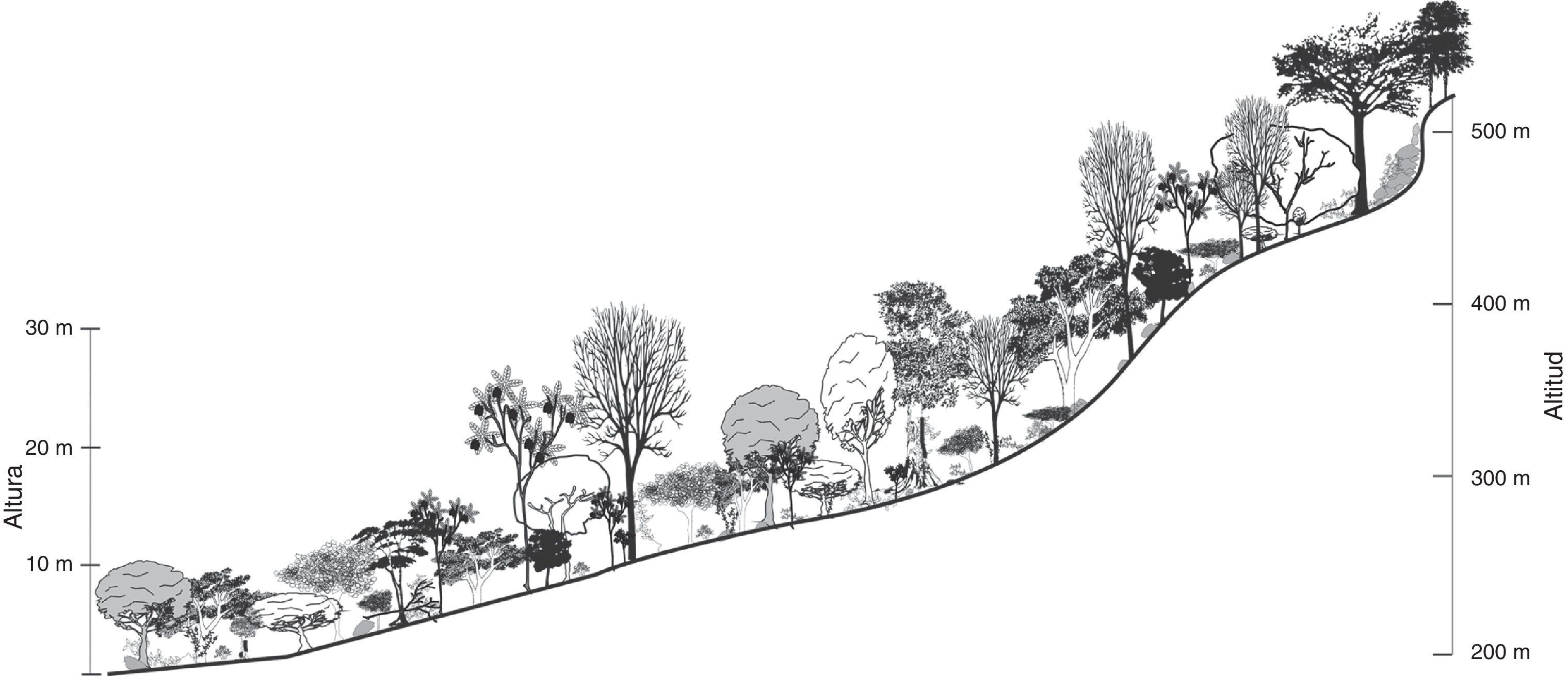
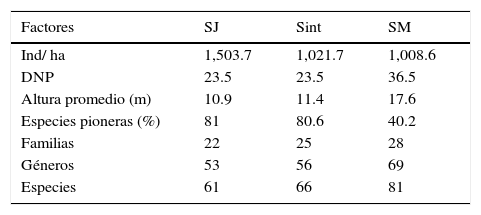
Selva secundaria inicial o juvenilSe observó en las localidades de Cuatenahuatl, El Aguacate y Tzocohuijapa, en sitios donde la perturbación de la vegetación fue elevada por la existencia de plantaciones forestales, actividades ganaderas, terrenos agrícolas y tala de árboles; la altitud en las 3 localidades fue similar, fluctuó entre 200 y 530 m; los valores estructurales y la composición de especies se muestran en la tabla 2. Las familias con mayor número de especies fueron Fabaceae (12), Malvaceae (8) y Moraceae (6). Las especies con mayor VIR en estas localidades se caracterizaron por su alta densidad de tallos (1,503.7 individuos/ha en promedio), 10.9 m de altura promedio y DN pequeños, de 23.5cm en promedio (tabla 2), por lo que los VIR están más relacionados con la densidad y frecuencia relativa.
En cuanto a la estructura vertical de la vegetación, se distinguió un estrato emergente compuesto por pocos individuos de más de 20 m de altura, de las especies Cedrela odorata y Ceiba pentandra. El estrato arbóreo intermedio con una altura de entre 10 y 15 m, fue el de mayor riqueza y codominancia, al presentar 12 especies con VIR similar (tabla 3). Las especies características fueron: B. simaruba, Cecropia obtusifolia, C. odorata, Croton draco, G. ulmifolia, Inga vera, P. copal y Pseudolmedia oxyphyllaria. El estrato arbóreo inferior, de entre 4 y 9 m de altura, estaba representado principalmente por A. barbinervis, Cnidoscolus multilobus, Dendropanax arboreus, Pleurantothodendron lindenii, Sabal mexicana, Tabernaemontana alba y Trichospermum mexicanum; en el mismo estrato se encontraron varias especies de Heliocarpus, Piper y de bejucos y aráceas. En los sitios de mayor altitud (500 m) se desarrollaban encinares tropicales dominados por Quercus qlaucoides y Quercus oleoides (fig. 2).
Selva secundaria intermedia
Se encontró en 3 localidades: fraccionamiento Núñez Soto, Metlaltepec y Tohuaco, todas ubicadas en serranías y cañadas con alto grado de perturbación, principalmente a causa de la agricultura, ganadería y las plantaciones forestales (café y plátano). La altitud fluctuó entre 100 y 460 m.
La riqueza taxonómica total fue de 66 especies 56 géneros y 25 familias, y la riqueza promedio de 39 especies por localidad. La familia Fabaceae fue la más representativa, con 14 especies, junto con Malvaceae y Moraceae, con 6 cada una y Euphorbiaceae con 5.
Los valores de densidad arbórea en las 3 localidades fluctuaron entre 810 y 1,250 individuos/ha, con un promedio de 1,021.7 individuos/ha. El DNP fue de 23.5cm y la altura promedio del dosel de 11.4 m, con especies emergentes de hasta 20 m de alto. El porcentaje de especies pioneras y el DNP fueron similares a los registrados en la SMS en estadios tempranos, pero la densidad fue menor y la altura ligeramente mayor. Las especies con los VIR más altos en el estadio intermedio fueron B. simaruba y G. ulmifolia. Sin embargo, por la dominancia de las especies y fisonomía de la vegetación se identificaron 3 asociaciones principales: selva de B. simaruba-Protium copal-G. ulmifolia, selva de G. ulmifolia-B. simaruba-S. mexicana, y selva de G. ulmifolia (tabla 3).
En cuanto a la distribución vertical de las especies, el estrato superior estaba compuesto por árboles emergentes de B. simaruba, D. arboreus y Enterolobium cyclocarpum, de más de 20 m de altura. El estrato intermedio fue el más abundante con alturas de 8 a 12 m; las especies con mayor cobertura fueron A. barbinervis, G. ulmifolia, Heliocarpus donnell-smithii, Piscidia pisipula, S. mexicana, algunos individuos de Ficus sp. e incluso Q. glaucoides (Tohuaco). El estrato inferior midió entre 4 y 8 m de altura y estaba representado por Cecropia obtusifolia, Croton draco, C. guatemalensis, Protium copal, Tabernaemontana alba y Trophis racemosa (fig. 3).

Las especies estructuralmente importantes dentro de esta comunidad fueron de porte bajo y pocas especies pioneras dominaron el dosel, fisionomía muy parecida a la selva secundaria joven, sin embargo, la densidad de especies fue menor, lo que se considera como indicador de un estado sucesional más avanzado (tabla 3).
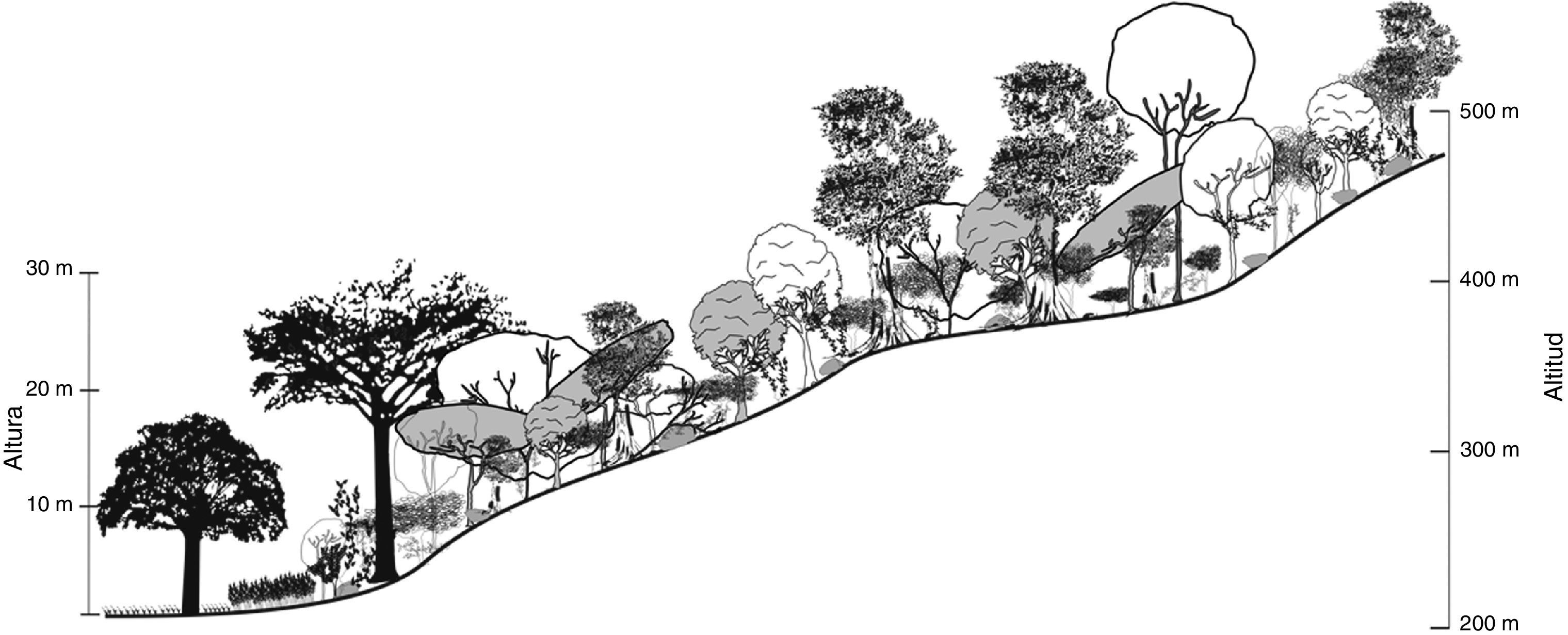
Selva secundaria maduraLas 3 localidades que se definieron en este estadio sucesional (Chalingo, Cojolite y Huemaco), se desarrollaban tanto en sitios con pendientes pronunciadas como en tierras bajas, en un intervalo altitudinal de 90 a 470 m.
En esta etapa la SMS presentó los valores más altos de riqueza taxonómica, albergando 81 especies, 69 géneros y 28 familias; las familias más representativas fueron Fabaceae (19 especies), Malvaceae (8 especies), Moraceae (7 especies), Euphorbiaceae (5 especies), Meliaceae (5 especies), y fue el estadio con más representantes de Sapotaceae (4 especies).
La SMS madura presentó la menor densidad arbórea con 1,008.6 individuos/ha, así como el mayor diámetro de tallos (DNP de 36.5cm); la altura promedio de los individuos fue de 17.6 m (tabla 1). La especie arbórea dominante B. alicastrum, presentó densidad, frecuencia y área basal elevadas, lo que le confirió un VIR muy alto (tabla 4). A nivel de familia, Moraceae y Sapotacea fueron las de mayor relevancia estructural.
El estrato superior o emergente de la SMS madura estaba representado por C. pentandra, E. cyclocarpum, Zuelania guidonia, Ficus spp. y Pouteria sp., árboles cuya altura sobrepasó los 30 m. El estrato intermedio fue dominante y midió entre 15 y 25 m de altura, las especies con mayor cobertura fueron B. alicastrum, B. simaruba, D. arboreus, Licaria capitata, P. piscipula y P. oxyphyllaria. En el estrato arbóreo inferior, de 7 a 12 m de altura, los individuos de A. barbinervis, G. ulmifolia, P. copal y T. alba, principalmente, fueron frecuentes (fig. 4).
Clasificación y ordenación de la vegetación
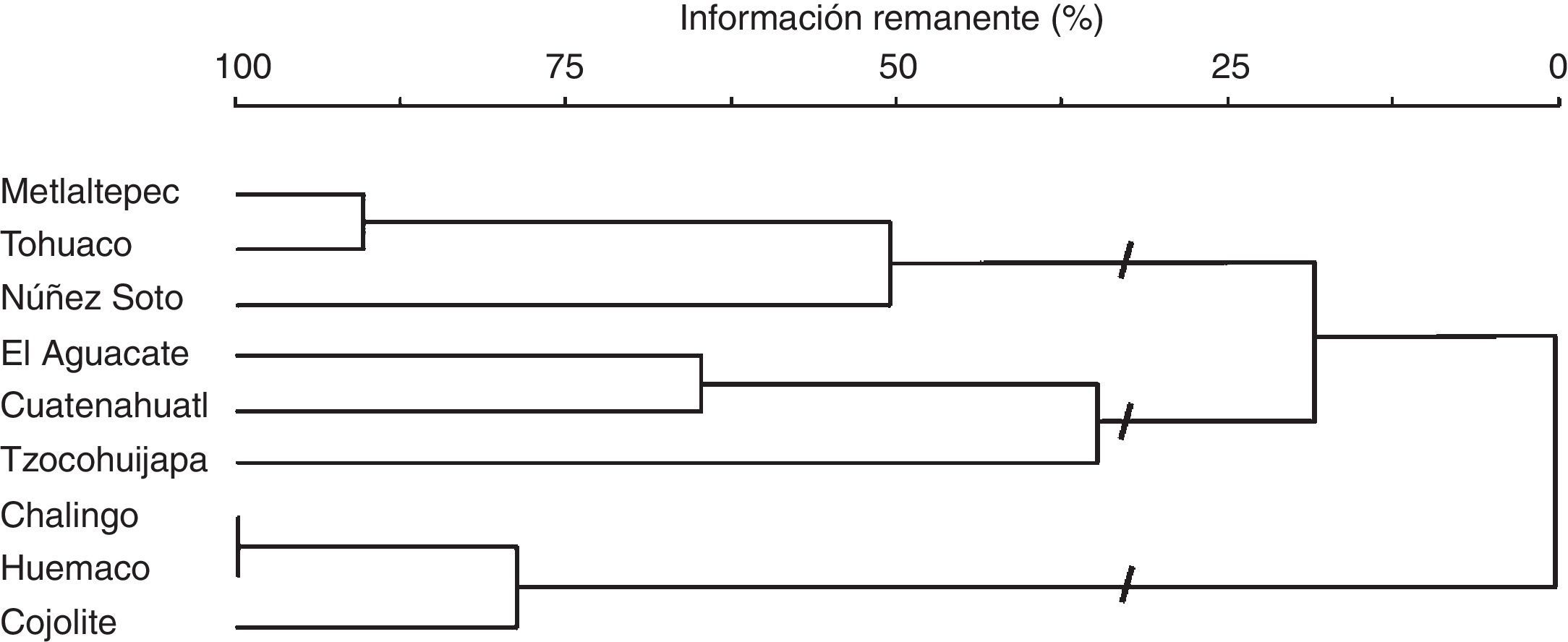
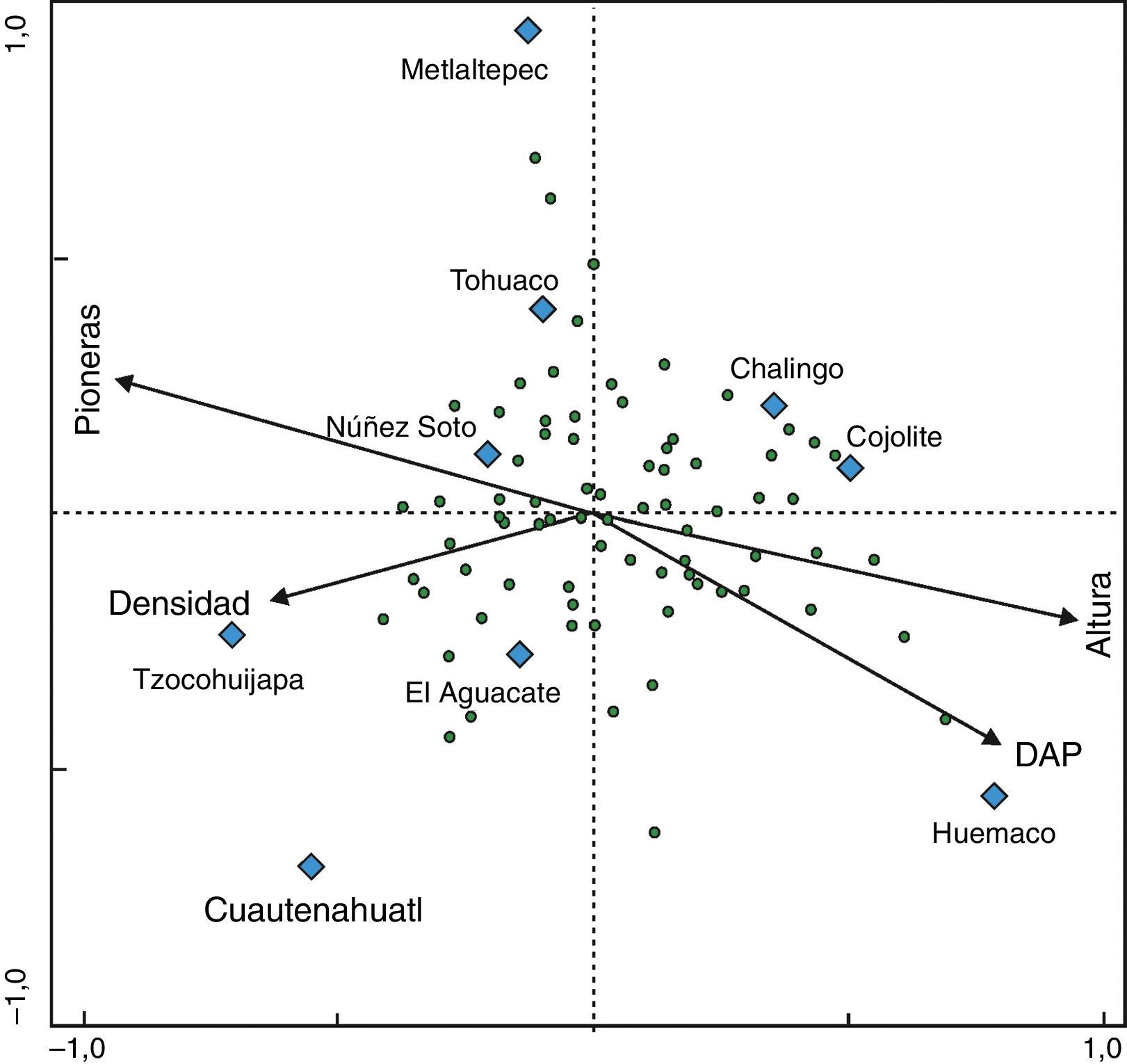
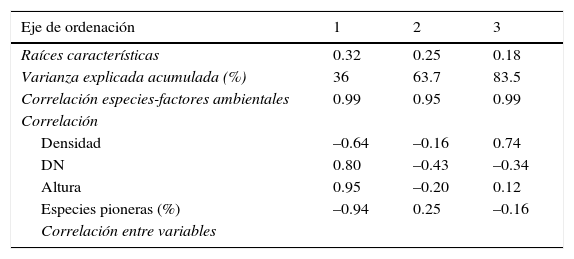
El análisis de agrupamiento, los valores estructurales de las especies y los datos de la literatura revisada, permitieron definir 3 grupos (figs. 5 y 6) que corresponden a las diferentes etapas de la sucesión de la SMS: juvenil, intermedia y madura. En el análisis de correspondencia canónica, los 2 primeros ejes explicaron el 63% de la variación en los datos; las variables con mayor correlación positiva con respecto al primer eje fueron la altura del dosel y el DN, y las de mayor correlación negativa fueron el porcentaje de especies pioneras y la densidad de tallos. Con respecto a los otros ejes de ordenación (tabla 5), solo la variable densidad de tallos presentó correlación positiva alta (0.736) con el eje 3. Los resultados obtenidos sugieren que durante la sucesión, a medida que las comunidades maduran, la densidad de tallos y la abundancia de las especies pioneras disminuye, se acumula mayor cantidad de biomasa (mayor diámetro de los tallos y mayor altura del dosel) y la estructura de la vegetación alcanza mayor complejidad. Las variables estructurales más relevantes para el reconocimiento de las diferentes etapas de la sucesión fueron el DN y la altura del dosel; en el diagrama de ordenación el porcentaje de especies pioneras no permitió diferenciar claramente las etapas juvenil e intermedia de la sucesión (fig. 6, tabla 5 y 6).

Dendrograma del análisis de agrupamiento que muestra las relaciones jerárquicas entre las localidades de muestreo. Cada rama está etiquetada con el nombre de la localidad. La escala horizontal es la función objetiva de Wishart, e indica el porcentaje de información presente a medida que los grupos se fusionan.
.")
Resultados del análisis de correspondencia canónica en donde se indican los valores de las raíces características, la correlación entre las especies y los valores estructurales, y el porcentaje de varianza explicada por los datos de las especies y por las relaciones especies-estructura, para los 3 primeros ejes de la ordenación.
| Eje de ordenación | 1 | 2 | 3 |
|---|---|---|---|
| Raíces características | 0.32 | 0.25 | 0.18 |
| Varianza explicada acumulada (%) | 36 | 63.7 | 83.5 |
| Correlación especies-factores ambientales | 0.99 | 0.95 | 0.99 |
| Correlación | |||
| Densidad | –0.64 | –0.16 | 0.74 |
| DN | 0.80 | –0.43 | –0.34 |
| Altura | 0.95 | –0.20 | 0.12 |
| Especies pioneras (%) | –0.94 | 0.25 | –0.16 |
| Correlación entre variables | |||
| Densidad | DN | Altura | |
|---|---|---|---|
| Densidad | 1 | – | – |
| DN | –0.68 | 1 | – |
| Altura | –0.48 | 0.85 | 1 |
| Especies pioneras (%) | 0.42 | –0.83 | –0.99 |
DN: diámetro normal.
Resumen de los valores taxonómicos y estructurales de cada uno de los estados sucesionales.
| Factores | SJ | Sint | SM |
|---|---|---|---|
| Ind/ ha | 1,503.7 | 1,021.7 | 1,008.6 |
| DNP | 23.5 | 23.5 | 36.5 |
| Altura promedio (m) | 10.9 | 11.4 | 17.6 |
| Especies pioneras (%) | 81 | 80.6 | 40.2 |
| Familias | 22 | 25 | 28 |
| Géneros | 53 | 56 | 69 |
| Especies | 61 | 66 | 81 |
DNP: diámetro normal promedio; Sint: selva mediana intermedia; SJ: selva mediana incial o juvenil; SM: selva mediana madura.
El único estudio sobre la composición arbórea de la selva mediana subperennifolia de Hidalgo es el realizado por Puig (1991), en el cual menciona 49 especies de árboles para la región de la Huasteca. En la presente investigación se registraron 92 especies con forma de vida arbórea en un solo municipio de la entidad. Otros trabajos en el mismo tipo de vegetación en distintas regiones de México han registrado valores de riqueza taxonómica un poco más elevados, por ejemplo: 101 especies arbóreas en el centro de Veracruz (Godínez-Ibarra y López-Mata, 2002), 105 especies en Quintana Roo (Sánchez et al., 2007) y 128 especies en el sur de Yucatán (Zamora-Crescencio, García, Flores y Ortiz, 2008).
De las familias, Fabaceae fue la más frecuente y la más representativa por el número de especies en la SMS de Huautla, tal como lo han mencionado diferentes autores para el mismo tipo de vegetación, en otras entidades de México (Godínez-Ibarra y López-Mata, 2002; Gutiérrez-Báez, Ortiz-Díaz, Flores-Guido y Zamora-Crescencio, 2012; Sánchez et al., 2007; Zamora-Crescencio et al., 2012). La mayoría de las especies de Fabaceae registradas en el presente estudio tienen amplia distribución en la SMS de México y por lo tanto, presentan alta tolerancia a diferentes condiciones ambientales, además de que son comunes en vegetación secundaria (Pennington y Sarukhán, 1998; Puig, 1991; Rzedowski, 1978; Zamora-Crescencio et al., 2008).
Es interesante destacar que el valor del índice de diversidad de Shannon estimado para las especies arbóreas en el municipio de Huautla, Hidalgo (H’ = 4.02) fue mayor al obtenido en otras localidades con SMS de México, como Papantla, Veracruz (H’ = 2.12; Basáñez, Alanís y Badillo, 2008), Vega de Alatorre, Veracruz (H’ = 3.30; Godínez-Ibarra y López-Mata, 2002), Calakmul, Campeche (H’ = 3.29; Zamora-Crescencio et al., 2012), Hampolol, Campeche (H’ = 2.28; Gutiérrez-Báez, Zamora-Crescencio y Puc-Garrido, 2013) y en el ejido Andrés, Quintana Roo (H’ = 3.27; Carreón-Santos y Valdez-Hernández, 2014). Las diferencias en los valores de diversidad obtenidos pueden estar influidas por las diferentes técnicas de muestreo utilizadas y no necesariamente por las condiciones ambientales. Las técnicas de muestreo que consideran la distancia entre árboles en lugar del área (parcelas de muestreo), son poco sensibles a la agregación de los individuos (Bryant et al., 2005; Forbes y Craig, 2013), por lo que el valor de equidad (distribución de abundancias de las especies) generalmente es más elevado y por ende, el valor del índice de diversidad también.
El análisis de correspondencia canónica mostró que las variables estructurales más relacionadas con la composición y fisionomía de la vegetación fueron la altura y el diámetro de los árboles, así como las especies pioneras con mayor VIR. La altura y el diámetro promedio de los individuos incrementan en valor conforme transcurre el proceso de sucesión secundaria en las selvas tropicales, por lo que pueden ser considerados como indicadores de la etapa de madurez de la vegetación (Martínez-Ramos y García-Orth, 2007). El VIR de las especies pioneras está correlacionado negativamente con la altura y el diámetro de los individuos; es decir, a medida que aumenta la altura y el diámetro de los árboles (conforme avanzan los estados de la sucesión) la representatividad estructural de las especies pioneras es cada vez menor (Lohbeck et al., 2013).
En el presente estudio, el porcentaje de especies pioneras, la densidad y la altura promedio de los árboles fue semejante entre los estadios de la sucesión definidos como juveniles (iniciales) e intermedios, lo que podría ser el resultado de una amplia variedad de factores ambientales (Leigh et al., 2004; Sánchez et al., 2007). Sin embargo, el uso del promedio de altura y diámetro de los árboles y el porcentaje de especies pioneras como únicos criterios para definir el estadio de regeneración de la vegetación no es adecuado, dado que pueden existir individuos de especies presentes antes del disturbio, las cuales pueden aumentar significativamente el promedio en ambas características. Es común por ejemplo, que los árboles de porte grande no sean talados, ya sea porque proporcionan sombra, o bien porque es difícil cortarlos (Guevara, Laborde y Sánchez-Ríos, 2005); o bien que algunas especies consideradas como pioneras sean elementos importantes en las comunidades maduras (Sánchez et al., 2007; van Breugel, Bongers y Martínez-Ramos, 2007; Zamora-Crescencio et al., 2008) o viceversa, que especies típicas de las últimas etapas de la sucesión estén presentes en las primeras fases del proceso, de acuerdo con la teoría de la composición florística inicial (sensuEgler, 1954; van Breugel et al., 2007).
La densidad de individuos por sí misma, no representa tampoco un dato concluyente; la dinámica de claros en etapas avanzadas de la sucesión debido a la senescencia de las plantas permite el establecimiento de plantas heliófitas de rápido crecimiento, lo que aumenta la densidad de los tallos de la comunidad madura (Yamamoto, 2000). Debido a lo mencionado en los párrafos previos, es necesario considerar varias características en conjunto para identificar con certeza los diferentes estadios de regeneración de la vegetación, por la naturaleza multifactorial y grado de complejidad del proceso (Guevara et al., 2005; Leigh et al., 2004; Sánchez et al., 2007).
En general, en el presente estudio los estadios sucesionales indicaron un aumento gradual de la altura y del área basal de las especies arbóreas, así como la disminución en la representatividad de las especies pioneras, conforme la selva alcanzó la madurez. Aun cuando la relación entre la densidad de tallos y la etapa de la sucesión fue baja, en el análisis de correspondencia canónica el orden de las localidades por etapas sucesionales fue inverso a la densidad de individuos, por lo que es posible que esta última variable pueda ser un buen indicador de la etapa de madurez, como previamente lo habían sugerido Guariguata y Ostertag (2002). La densidad de tallos disminuye en etapas avanzadas de la sucesión, debido a que las condiciones de umbría ocasionadas por la elevada cobertura del dosel en estadios tardíos de la sucesión (comunidades maduras), impiden el desarrollo de las especies heliófilas (Turner, 2004).
De acuerdo con los resultados obtenidos en el presente estudio y de otras investigaciones (Guariguata y Ostertag, 2001; Lohbeck et al., 2013), es factible clasificar los diferentes estados de la sucesión de la SMS por medio del VIR de las especies, más que por el tiempo de regeneración, ya que la estructura de la vegetación, en conjunto con la composición de especies, están relacionados o son dependientes de diferentes factores estocásticos que se presentan después de un disturbio: como el banco de semillas, la vegetación remanente, la dispersión de semillas, la biología de las especies y sus interacciones, así como con factores abióticos, principalmente luz y propiedades edáficas (Holl, Stout, Reid y Zahawi, 2013).
Algunas localidades se definieron dentro del mismo estadio sucesional porque contenían algunos elementos florísticos representativos en común, pero la composición y valores estructurales de las especies no fueron idénticas, debido a que la trayectoria de la sucesión difiere en cada sitio y etapa: es decir, en estadios tempranos está definida principalmente por las características ambientales presentes en el momento de la colonización, mientras que en los más avanzados adquieren relevancia la capacidad competitiva y la tolerancia de las especies a las condiciones ambientales (Holl et al., 2013; Letcher et al., 2012).
El análisis de agrupamiento permitió definir a Chalingo, Cojolite y Huemaco como localidades con SMS en etapa de madurez, pero con distintas trayectorias de regeneración, por la dominancia en el dosel de Brosimum-Bursera, Brosimum-Ficus y Brosimum-Guazuma. Las 3 localidades se caracterizan por la presencia de especies de la familia Sapotaceae y por la dominancia en el dosel de B. alicastrum, como se ha registrado para las selvas secundarias maduras o primarias en la región (Puig, 1991; Puig y Lacaze, 2004; Rzedowski, 1978) y en otras entidades del país con el mismo tipo de vegetación (selva mediana subperennifolia sensuMiranda y Hernández, 1963), como San Luis Potosí (Rzedowski, 1961) y Veracruz (Godínez-Ibarra y López-Mata, 2002; Palacios-Wassenaar, Castillo-Campos, Vázquez-Torres y del Amo-Rodríguez, 2014). Se trata de una especie con alta tolerancia ecológica y dominancia en etapas finales de la sucesión en la SMS, conformando un dosel continuo y denso, con estratos inferiores pobres en especies, especialmente el herbáceo (Macario et al., 1995; Puig, 1991). Sánchez et al. (2007) y Zamora-Crescencio et al. (2008) mencionan que las especies pioneras heliófilas de larga vida (que se caracterizan por su capacidad de establecerse bajo el dosel arbóreo, pero requieren necesariamente de la formación de claros para crecer), son comunes y abundantes en todas las fases de regeneración y algunas de ellas pueden llegar a ser dominantes en etapas avanzadas. Acorde con lo anterior, el porcentaje de especies pioneras fue similar en las etapas de la sucesión juvenil e intermedia de la SMS de Huatla; y aunque la mayoría de las especies registradas en la fase madura pueden considerarse como árboles no pioneros o de vegetación primaria (Pennington y Sarukhán, 1998), también fue común encontrar algunas especies heliófilas o pioneras de larga vida. Es probable también que el alto porcentaje de especies pioneras en todas las localidades analizadas, esté relacionada con las alteraciones de origen natural o por actividades humanas, que por su constancia generan condiciones adecuadas para la incorporación de especies de rápido crecimiento sin importar la etapa de la sucesión (Rzedowski, 1978; Zamora-Crescencio et al., 2008).
La presencia de corredores de vegetación en distintas etapas de regeneración, como ocurre en la SMS del municipio de Huautla, puede facilitar el restablecimiento de poblaciones de especies características de vegetación primaria. La vegetación secundaria contigua a un remanente de selva primaria, puede proveer recursos extras para algunas especies, facilitando el movimiento y amortiguando cambios en el microclima del fragmento adyacente (Chazdon et al., 2009; Forbes y Craig, 2013; Turner y Richard, 1996; Vilchez, Chazdon y Milla, 2008). Turner y Richard (1996) indican que las selvas secundarias parecen ser ricas en especies en áreas pequeñas, sin embargo, en áreas grandes suelen ser florísticamente monótonas y contienen menos especies que las selvas primarias, lo que concuerda con los valores de riqueza más altos, obtenidos en el presente estudio para la SMS en etapa madura.
Los autores agradecen el apoyo parcial obtenido a través del proyecto FOMIX-Hidalgo titulado «Diversidad biológica del estado de Hidalgo» 191908 (tercera etapa). El primer autor agradece ampliamente al Consejo Nacional de Ciencia y Tecnología, por la beca número 368818 otorgada para la realización de sus estudios de posgrado. Se agradece también el apoyo parcial obtenido por parte de la Universidad Autónoma del Estado de Hidalgo, Presupuesto anual de Investigación, a través del proyecto «Nivel de conservación de especies consideradas en riesgo de reducción poblacional en México» (clave ICBI-PAI-29).
| Familia/Especie | SJ | Sint | SM | Est | Nombre común |
|---|---|---|---|---|---|
| Anacardiaceae | |||||
| Spondias mombium L. | 1 | 0 | 1 | HL | Jobo |
| Annonaceae | |||||
| Rollinia membranacea Triana y Planch | 0 | 0 | 1 | ST | Anona |
| Apocynaceae | |||||
| Rauvolfia tetraphylla L. | 1 | 0 | 1 | ST | Ajillo, apasmo |
| Stemmadenia donnell-smithii (Rose) Woodson | 1 | 1 | 0 | ST | Cojón de toro, huevo de burro |
| Tabernaemontana alba Mill. | 1 | 1 | 1 | HL | Cojón de gato, toctoxite |
| Araliaceae | |||||
| Dendropanax arboreus (L.) Planch. Et Decne | 1 | 1 | 1 | ST | Tamalcahuite, mano de león, olivo |
| Arecaceae | |||||
| Sabal mexicana Mart. | 1 | 1 | 1 | HL | Palma real, ápatz, apachijtli |
| Bignoniaceae | |||||
| Parmentiera aculeata (Kunth) Seem | 1 | 1 | 1 | HL | Chote |
| Tabebuia rosea (Bertol.) DC. | 1 | 1 | 1 | HL | Palo de rosa |
| Boraginaceae | |||||
| Cordia alliodora (Ruiz y Pav.) Oken | 0 | 1 | 0 | HL | Bojón, suchicuahua, chachalaco |
| Burseraceae | |||||
| Bursera simaruba (L.) Sarg. | 1 | 1 | 1 | Hl | Chaca, palo mulato |
| Protium copal (Schltdl. y Cham.) Engl. | 1 | 1 | 1 | ST | Copal |
| Calophyllaceae | |||||
| Calophyllum brasiliense Cambess. | 0 | 0 | 1 | E | Zapotillo |
| Clethraceae | |||||
| Clethra mexicana DC. | 1 | 0 | 1 | HL | Aguacatillo |
| Ebenaceae | |||||
| Diospyros digyna Jacq. | 0 | 0 | 1 | ST | Zapote negro, muneque |
| Euphorbiaceae | |||||
| Adelia barbinervis Schltdl. y Cham. | 1 | 1 | 1 | HL | Espino blanco, itztacuitzli |
| Cnidoscolus multilobus (Pax) I.M. Johndt. | 1 | 1 | 1 | H | Mala mujer, ortiga |
| Croton draco Schltdl. | 1 | 1 | 1 | H | Sangregrado, escuáhuitl |
| Croton reflexifolius Kunth | 1 | 0 | 1 | H | Olivo |
| Croton guatemalensis Losty | 1 | 0 | 1 | H | Huilocuahuitl |
| Fabaceae | |||||
| Acacia cornigera (L.) Willd | 1 | 1 | 1 | H | Cuerno de toro, cornezuelo |
| Acacia coulteri A.Gray | 1 | 1 | 1 | H | - |
| Bauhinia divaricata L. | 1 | 0 | 1 | HL | Papalocuáhuitl, pata de cabra |
| Caesalpinia mexicana A. Gray | 1 | 0 | 1 | ST | Brasil |
| Cojoba arborea (L.) Britton y Rose | 1 | 1 | 0 | ST | Frijolillo, ecuahuitl |
| Diphysa sennoides Benth. | 1 | 1 | 1 | - | Quebracha, tenquiques |
| Enterolobium cyclocarpum (Jacq.) Griseb | 1 | 0 | 1 | HL, | Orejón, guanacaste |
| Erythrina americana Mill. | 1 | 1 | 1 | ST | Pemuche, colorín |
| Erythrina herbacea L. | 1 | 1 | 1 | ST | - |
| Gliricidia sepium (Jacq.) Steud. | 0 | 1 | 1 | HL | Cocuite, cacahuananche |
| Inga jinicuil G. Don | 0 | 1 | 1 | HL | Chalahuite, jinicuil |
| Inga vera Willd. | 0 | 1 | 1 | HL | Chalahuite |
| Leucaena pulverulenta (Schltdl.) Benth. | 1 | 1 | 0 | HL | Huashe |
| Lysiloma acapulcensis (Kunth) Benth. | 0 | 1 | 1 | HL | Tepehuaje |
| Mucuna pruriens (L.) DC. | 1 | 0 | 1 | HL | Pica-pica |
| Piscidia piscipula (L.) Sarg. | 0 | 0 | 1 | HL | Chijol |
| Pithecellobium dulce (Roxb.) Benth. | 0 | 0 | 1 | HL | Humo |
| Pithecellobium lanceolatum (Humb. y Bonpl. ex Willd.) Benth. | 0 | 0 | 1 | HL | Huamúchil |
| Pterocarpus sp. L. | 0 | 0 | 1 | ST | - |
| Senna sp. Mill. | 1 | 0 | 1 | HL | - |
| Senna spectabilis (DC.) H.S. Irwin y Barneby | 0 | 0 | 1 | HL | Candelilla o candelillo |
| Fagaceae | |||||
| Quercus glaucoides M. Martens y Galeotti | 1 | 1 | 1 | E | Encino |
| Quercus oleoides Schltdl. y Cham. | 1 | 1 | 0 | E | Encino, tesmol |
| Lauraceae | |||||
| Licaria capitata (Schltdl. y Cham.) Kosterm. | 1 | 1 | 1 | ST | Misanteco, cuishcuahuitl |
| Persea americana Mill. | 1 | 1 | 1 | ST | Aguacate |
| Persea shiedeana Nees | 1 | 1 | 1 | ST | Ahuacachile, pahua, aguacatillo |
| Malvaceae | |||||
| Carpodiptera ameliae Lundell | 1 | 0 | 1 | E | Petcum, hoja anchom, peltón |
| Ceiba pentandra (L.) Gaertn. | 1 | 1 | 1 | H,E | Ceiba |
| Guazuma ulmifolia Lam. | 1 | 1 | 1 | HL | Guácima |
| Hampea nutricia Fryxell | 1 | 1 | 1 | ST | Jonote blanco, majagua |
| Heliocarpus appendiculatus Turcz. | 1 | 1 | 1 | H | Jonote rojo |
| Heliocarpus donnell-smithii Rose | 1 | 1 | 1 | H | Jonote baboso |
| Muntigia calabra L. | 1 | 1 | 0 | H | Capulín, capulincillo |
| Pleuranthodendron lindenii (Turcz.) Sleumer | 1 | 1 | 1 | H | Polocastle, palo de maíz |
| Trichospermum mexicanum (DC.) Baill. | 1 | 0 | 1 | H | Jonote capulín, cuapetate |
| Melastomataceae | |||||
| Conostegia xalapensis (Bonpl.) D. Don ex DC. | 1 | 1 | 1 | H | Capulincillo, teshuate |
| Meliaceae | |||||
| Cedrela odorata L. | 1 | 1 | 0 | HL | Cedro |
| Melia azedarach L. | 1 | 1 | 1 | HL | Pioche, piocha, paraíso |
| Swietenia macrophylla King | 1 | 0 | 1 | HL, E | Caoba |
| Trichilia americana (Sessé y Moc.) T.D.Penn. | 0 | 0 | 1 | HL | - |
| Trichilia havanensis Jacq. | 1 | 1 | 1 | HL | Xopilcuáhuitl, palo de cuchara |
| Moraceae | |||||
| Brosimum alicastrum Sw | 1 | 1 | 1 | S,E | Ojite, ramón |
| Ficus crocata (Miq.) Miq. | 1 | 1 | 1 | S,E | Jalamate negro, amate prieto |
| Ficus maxima Mill. | 1 | 0 | 1 | S,E | Jalamate grande |
| Ficus sp. | 1 | 0 | 0 | S | Amate |
| Ficus tecolutensis (Liebm.) Miq. | 1 | 1 | 1 | S,E | Jalamate chico, mata palo |
| Maclura tinctoria (L.) D. Don ex Steud | 0 | 1 | 1 | HL | Mora |
| Pseudolmedia oxyphyllaria Donn.Sm. | 1 | 0 | 1 | ST | Huelor, ramón colorado, ramón de mico |
| Trophis racemosa (L.) Urb. | 0 | 1 | 1 | S | Zacate ramón, asincuahuitl |
| Myrtaceae | |||||
| Eugenia oerstediana O. Berg | 1 | 0 | 1 | H | Pistlillo |
| Psidium guajava L. | 1 | 0 | 1 | ST | Guayaba |
| Picramniaceae | |||||
| Alvaradoa amorphoides Liemb. | 0 | 0 | 1 | HL | Camarón, pie de gallo |
| Picramnia antidesma Sw. | 1 | 0 | 0 | - | Huihuitecómit, chilillo |
| Piperaceae | |||||
| Piper arboreum Aubl. | 1 | 1 | 0 | H | - |
| Piper sp. L. | 1 | 1 | 1 | H | - |
| Platanaceae | |||||
| Platanus mexicana Moric. | 1 | 0 | 1 | E | Álamo, haya |
| Rubiaceae | |||||
| Psychotria sp. L. | 1 | 0 | 1 | ST | Cafecillo |
| Randia laetevirens Standl. | 1 | 0 | 0 | ST | Uña de gato, cruzeta |
| Rutaceae | |||||
| Murraya paniculata (L.) Jack | 1 | 0 | 0 | - | Limonaria |
| Zanthoxylum acuminatum (Sw.) Sw. | 1 | 0 | 0 | E | - |
| Salicaceae | |||||
| Salix humboldtiana Willd. | 1 | 0 | 1 | HL | Sauce |
| Zuelania guidonia (Sw.) Britt y Millsp. | 0 | 0 | 1 | ST,E | Volador, petlácotl |
| Sapindaceae | |||||
| Cupania dentata Moc. y Sessé ex DC. | 1 | 1 | 1 | HL | Cosalcahuite |
| Sapotaceae | |||||
| Chrysophyllum mexicanum Brandegee | 1 | 0 | 0 | ST | Thíjul, caimito, capulín |
| Manilkara sapota (L.) Van Royen | 1 | 0 | 1 | S | Chicozapote |
| Pouteria campechiana (Kunth) Seem | 0 | 0 | 1 | S | Chocohuite |
| Pouteria sapota (Jacq.) H.R.Moore y Strearn | 0 | 0 | 1 | S, E | Mamey, zapote mamey |
| Sideroxylon palmeri (Rose) T.D. Penn. | 1 | 0 | 1 | ST | Capulín |
| Solanaceae | |||||
| Cestrum dumetorum Schltdl. | 1 | 0 | 0 | H | Arcajuda |
| Ulmaceae | |||||
| Aphananthe monoica (Hemsl.) J.F. Leroy | 1 | 0 | 1 | S, E | Ajuate, mora, cuachichile |
| Trema micrantha (L.) Blume | 0 | 1 | 1 | HL | Pua, puam, puhuam |
| Urticaceae | |||||
| Cecropia obtusifolia Bertol. | 1 | 1 | 1 | H | Guarumo, picone |
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.