



Por medio de técnicas paleoecológicas se documentó la historia florística y ambiental del bosque mesófilo de montaña con presencia relictual de Acer saccharum subsp. skutchii del centro-occidente de México. La reconstrucción de la vegetación de los últimos 720 años se efectuó a través del núcleo de sedimento (TLP-N2) de 37cm de profundidad; los indicadores geoquímicos utilizados incluyen susceptibilidad magnética, fluorescencia de rayos X y pérdida por ignición; mientras que el microcarbón fue empleado como indicador de incendios forestales. Los resultados muestran el inicio de la pequeña edad de hielo a partir de 1341-1858 dC, con la presencia de abundantes taxones leñosos y pteridofitas. El periodo más seco se presentó de 1653-1720 dC, etapa que corresponde con el mínimo de Maunder en la actividad solar; este se caracteriza por un aumento de polen de herbáceas como Poaceae, Piper y Arisaema. Las condiciones ambientales actuales se establecieron a partir de 1871 dC, que se asemeja con las condiciones del periodo cálido medieval (1230-1319 dC), mientras que la actividad humana relacionada con la presencia de polen de maíz se infiere desde hace 720 años. Los resultados del registro paleoecológico sugieren que el bosque mesófilo ha sido fluctuante y resiliente durante el último milenio.
Through the use of palaeoecological techniques we inferred the environmental and floristic history of the cloud forest with relict presence of Acer saccharum subsp. skutchii in west-central Mexico. A 37cm depth sediment core (TLP-N2) was used to reconstruct the vegetation of the last 720 years; paleoenvironmental proxy consisted of magnetic susceptibility, XRF and loss of ignition, while microfossil charcoal was used as a proxy reflecting fire occurrence. The beginning of the Little Ice Age is reported around 1341-1858 with the presence of abundant woody taxa and pteridophytes. The driest period was detected from 1653 to 1720, which corresponds with the Maunder minimum in solar activity; this period is characterized by an increase in Poaceae, Piper and Arisaema. The present conditions were established since 1871, resembling the conditions of the Medieval Warm Period (1230-1319), while human activity related to the presence of maize pollen is inferred from 720 years ago. Results from this study suggest that cloud forests have been fluctuating but resilient over the last millennium.
Los patrones de distribución actual de la vegetación son un reflejo de cambios ambientales ocurridos en el pasado; dichos patrones presentan fluctuaciones en la abundancia y composición de taxones reconocibles a escala regional y local por medio de indicadores de cambio ambiental como la variación en la concentración elemental, la susceptibilidad magnética y la pérdida por ignición de sedimentos (Lozano-García y Vázquez-Selem, 2005; Verma, Lozano-Santa Cruz y Girón, 1996). Por otra parte, es posible reconstruir la historia de la vegetación utilizando como indicador el polen fósil de edades geológicas relativamente recientes como sería el caso del Holoceno, periodo durante el cual se establecieron las condiciones climáticas actuales con fluctuaciones en la temperatura y precipitación (Lozano-García y Cevallos-Ferriz, 2011). El paleoclima del Holoceno se caracteriza por un intervalo cálido conocido como el periodo cálido medieval (PCM), 1000-1300 dC (Mann et al., 2009), así como por la anomalía climática conocida como la pequeña edad de hielo (PEH), ocurrida entre 1350-1850 dC, que se define como un intervalo frío muy variable en el cual las temperaturas del hemisferio norte se redujeron menos de 1°C (Grove, 2001). Los principales forzamientos externos asociados a la PEH se caracterizaron por una baja radiación solar (Bond et al., 2001) y una actividad volcánica prolongada (Crowley, 2000; Shindell, Schmidt, Miller y Mann, 2003). En México un considerable número de secuencias sedimentarias de cuencas lacustres aportan evidencias sobre el cambio climático ambiental ocurrido durante la PEH asociada a la disminución de temperatura de 2°C del promedio normal (Lozano-García, Caballero y Ortega, 2007), así como una variabilidad considerable en las condiciones de humedad y sequías recurrentes en el siglo xviii (Cuna et al., 2014; Metcalfe, Jones, Davies, Noren y Mackenzie, 2010; Sosa-Nájera, Lozano-García, Roy y Caballero, 2010). En ecosistemas forestales es posible reconstruir estas oscilaciones ambientales y florísticas a través de núcleos de sedimento obtenidos en depresiones naturales, sobre los cuales se deposita polen a lo largo del tiempo (Calcote, 1995). Sin embargo, evidencias provenientes de bosques de montaña son escasas (Cerano-Paredes et al., 2013; Figueroa-Rangel, Willis y Olvera-Vargas, 2008, 2010, 2011; Ortega-Rosas, Peñalba, López-Sáez y van Devender, 2008).
El bosque mesófilo de montaña (BMM) es considerado un relicto de la flora del Neógeno (Graham, 1999; Rzedowski, 1996) en donde han sobrevivido linajes antiguos de floras divergentes (Luna-Vega y Magallón, 2010). El BMM ocupa menos del 1% del territorio mexicano y alberga la mayor riqueza de especies vasculares en comparación con otros bosques (Challenger, 1998; Rzedowski, 1991). Este tipo de vegetación generalmente se encuentra disperso sobre cañadas aisladas en donde frecuentemente recibe humedad en forma de neblina, factores que probablemente contribuyen para que su composición florística y ambiental sea muy contrastante entre sí. El BMM generalmente se localiza entre los 800-2,500m de altitud en temperaturas de 12 a 23°C (Rzedowski, 1996). En el occidente de México este tipo de vegetación se distribuye en 2 regiones fisiográficas: la sierra Madre del Sur y el Eje Volcánico Trasversal (Conabio, 2010). La historia de esta zona de transición biogeográfica es muy importante con el fin de entender la biodiversidad que le caracteriza; gran parte de ello se debe a la convergencia de las floras de origen Neártico y Neotropical, con la presencia de elementos florísticos de bosque caducifolio de la región de América del Norte y Asia, así como elementos de hoja perenne de América del Sur (Miranda y Sharp, 1950). El BMM presenta oscilaciones en la composición florística muy heterogéneas que varían de una localidad a otra; esta diversidad estructural corresponde a una singular composición de especies arbóreas establecidas durante los periodos glaciares, cuando las condiciones ambientales eran más frías y húmedas que las actuales (Palacios-Chávez y Rzedowski, 1993), convirtiéndose durante el Holoceno en refugio de diversos taxones de árboles templados por el aumento de la temperatura y la prevalencia de las condiciones más áridas (Vargas-Rodríguez, Vázquez-García, Quintero, Muñiz-Castro y Shalisko, 2010; Vázquez-García, Vargas-Rodríguez y Aragón, 2000; Vegas-Vilarrúbia, Nogué y Rull, 2012). Los atributos más importantes de esta comunidad vegetal son su alta biodiversidad y endemismos, además, cumple una función muy importante en la provisión de recursos hídricos, por lo que se convierte en una zona de prioridad crítica para su conservación (Conabio, 2010). El enfoque paleoecológico representa una alternativa muy apropiada que proporciona evidencias de la respuesta del BMM a los cambios ambientales pasados; los registros de estas características se pueden utilizar como análogos del pasado con el fin de inferir la respuesta del BMM a los cambios ambientales futuros (Vegas-Vilarrúbia, Rull, Montoya y Safont, 2011). De acuerdo con los registros paleoecológicos, la respuesta de las comunidades bióticas al cambio climático puede darse de diferentes formas en función de la escala espacio-temporal —migración de especies, cambios en la composición de la comunidad, cambios evolutivos o la extinción— (Vegas-Vilarrúbia et al., 2011). En este sentido se ha demostrado que el BMM en el occidente de México es resiliente al efecto de factores climáticos, sin embargo, es muy dependiente del clima predominante (Figueroa-Rangel et al., 2011). Las tendencias ambientales actuales son desfavorables para el BMM considerando el impacto de diversas actividades antropogénicas como la sobreexplotación de los recursos y el cambio de uso de suelo, que ocasionan la pérdida del hábitat y la fragmentación del bosque (Cayuela, Golicher y Rey-Benayas, 2006).
En este trabajo se presenta el análisis paleoecológico de los últimos 720 años del BMM dominado por Acer saccharum subsp. skutchii localizado en el centro-occidente de México. El objetivo del estudio fue reconstruir la historia de la vegetación de este bosque con el fin de responder a las siguientes preguntas: ¿Cuáles son los principales cambios en la composición taxonómica del BMM durante los últimos 720 años? ¿Cuál es la respuesta de este ante la variabilidad climática? ¿Cuál es el efecto de la dinámica ambiental en el recambio de taxones? y ¿Cuál ha sido la respuesta de la vegetación ante la perturbación antropogénica?
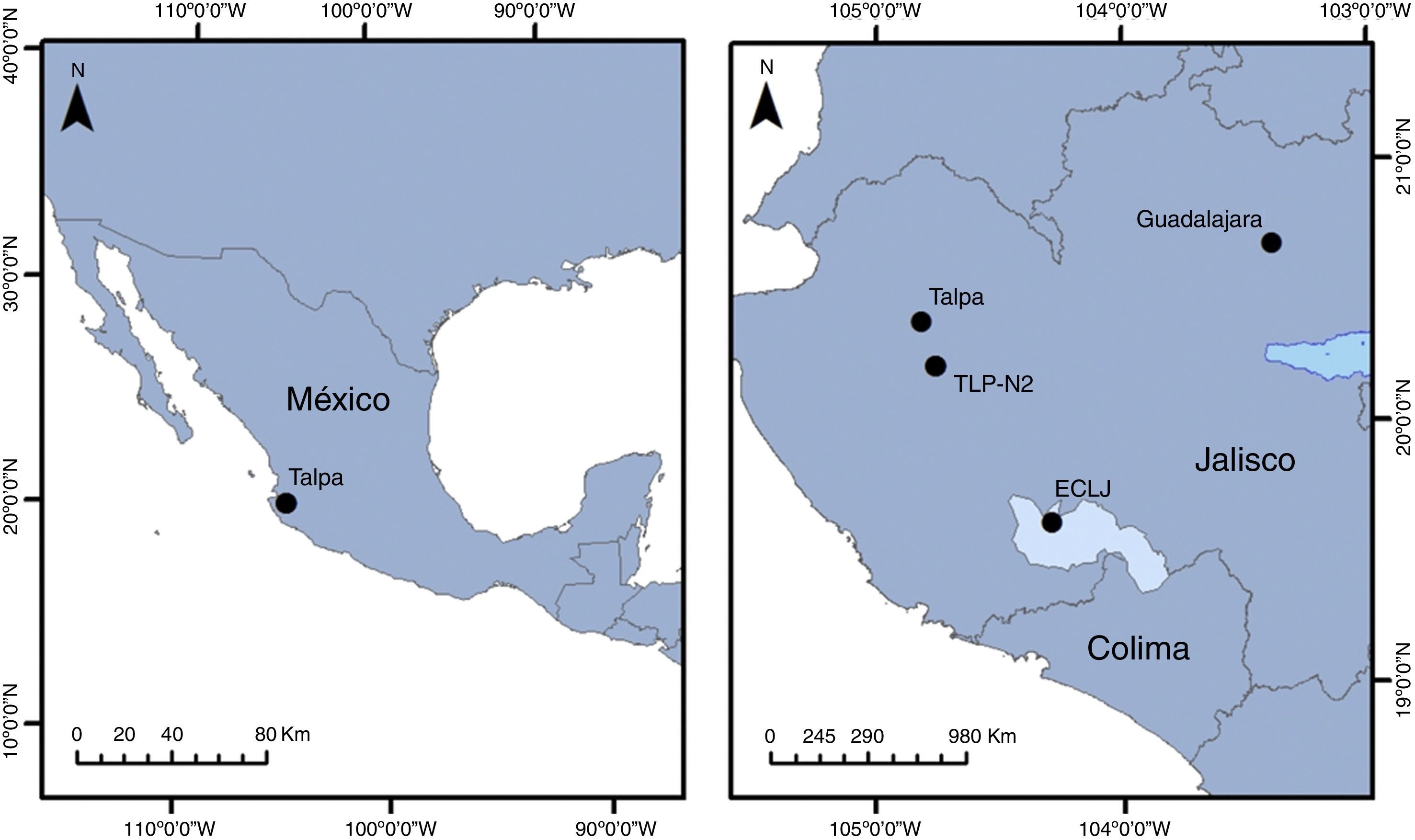
Materiales y métodosEl BMM con presencia de Acer saccharum subsp. skutchii es un pequeño remanente de este tipo de vegetación que se ubica a los 1,800m snm al este-sureste (104°49′ O, 20°23′ N) del municipio de Talpa de Allende, en la región sierra Occidental del estado de Jalisco. Los suelos dominantes son Regosol dístrico, Cambisol dístrico y Leptosol (Vargas-Rodríguez, 2005) y un clima templado subhúmedo con temperatura media anual de 21°C y precipitación pluvial promedio anual de 1,003mm. El área de estudio se encuentra en el sector occidental del Bloque Jalisco, que está formado por 2 zonas litológicas superficiales dominadas por rocas ígneas extrusivas del Oligoceno-Mioceno y andesitas del Terciario inferior (Maillol, Bandy y Ortega-Ramírez, 1997). La geomorfología del sitio es predominantemente montañosa. El estado actual de conservación de este bosque es bueno, de acuerdo con los estudios de Vargas-Rodríguez (2005) con alta densidad de árboles y de mayor tamaño diamétrico que en otras localidades de México.
El componente arbóreo dominante en la actualidad está representado por: Abies jaliscana, Acer saccharum subsp. skutchii, Carpinus tropicalis, Clethra flagrans, Cleyera integrifolia, Clusia salvinii, Conostegia volcanalis, Cornus disciflora, Cyathea costaricensis, Dendropanax arboreus, Ilex brandegeana, Ilex dugesii, Ostrya virginiana, Persea hintonii, Pinus sp., Podocarpus matudae, Quercus salicifolia, Symplocos citrea, Ternstroemia lineata y Zinowewia concinna; además, existe regeneración natural de Acer saccharum subsp. skutchii y Podocarpus matudae. El componente de herbáceas incluye helechos terrestres (e.g. Cyathea costaricensis), epífitos, musgos, líquenes, orquídeas y bromelias.
Las principales actividades antrópicas en el área de estudio son el cambio de uso de suelo en terrenos agrícolas y pastizales para ganado, minería y extracción de madera (Vargas-Rodríguez, Platt, Vázquez-García, Boquín, 2010; Vargas-Rodríguez, Vázquez-García, Quintero, Muñiz-Castro y Shalisko, 2010). El municipio de Talpa es poseedor de una tradición y cultura religiosa que se remonta a la época de la ocupación española con registros históricos de visitas de peregrinos por más de 3 siglos (Fernández-Poncela, 2012). Esta actividad la realizan desde distintos puntos del estado por parajes dentro del bosque (Vargas-Rodríguez, Platt, Vázquez-García, Boquín, 2010; Vargas-Rodríguez, Vázquez-García, Quintero, Muñiz-Castro y Shalisko, 2010). Los grupos prehispánicos que ahí se asentaron fueron de la fase Nahuapa del año 1000 aC, de la fase Aztatlán 600-1000 aC y de la fase Guayacán de 300-600 aC (Mountjoy, 1987) de origen náhuatl y conformaban un pueblo llamado Tlallipan que significa «sobre la tierra» perteneciendo al reino de Xalisco. A la llegada de los españoles en el año de 1540 fueron incorporados a los dominios de la corona española (Martínez-Réding, 1992).
Extracción de sedimentoSe extrajeron 9 núcleos de sedimento de una pequeña depresión en el bosque (forest hollow, sensuCalcote, 1995, 1998) a 1,773m snm en las coordenadas 20°12′56.18″ N, 104°45′25.63″ O (fig. 1); de estos solamente uno presentó las condiciones para la reconstrucción de la historia de la vegetación. Los núcleos de sedimento fueron extraídos con un equipo de perforación Eijelkamp y, posteriormente envuelto en papel film y aluminio, colocados en tubos de PVC de forma horizontal con el fin de mantener condiciones anaeróbicas. Posteriormente los núcleos fueron trasladados al Laboratorio de Paleoecología y Cambio Climático del Departamento de Ecología y Recursos Naturales de la Universidad de Guadalajara para conservarse a 5°C para su posterior análisis.

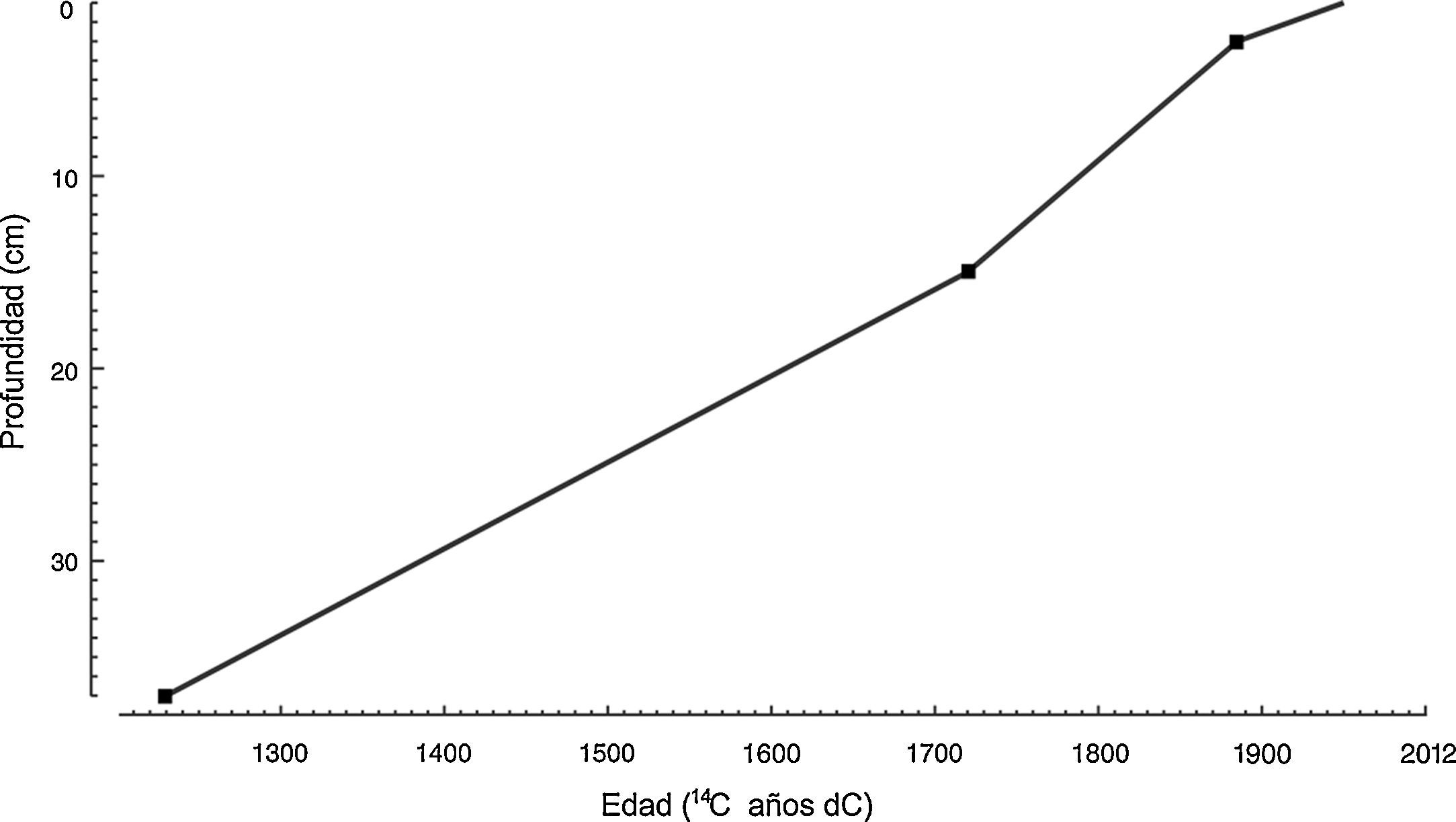
La cronología del núcleo de sedimento se determinó con base en 3 fechados de 14C por medio de la técnica accelerator mass spectrometer en el Laboratorio Beta Analytic, Miami, Florida, EE. UU. Para la calibración en años calendáricos fueron utilizados los programas Incal 04.14 y Calib versión 5.02 de Stuiver y Reimer (Stuiver y Reimer, 1993). Para elaborar la cronología se obtuvo un modelo edad-profundidad por interpolación lineal de acuerdo con el método de Bennett (1994) mediante el paquete Psimpoll versión 4.25 (tabla 1).
Con el fin de determinar la concentración de material ferromagnético asociada a la variabilidad ambiental del sitio de estudio, se realizó un análisis de susceptibilidad magnética, el cual es una medida del grado de magnetización de un material en respuesta del campo magnético y es directamente proporcional a la cantidad y tamaño de minerales magnéticos contenidos en la muestra que se expresa en unidades χ10 S.I. (Verosub y Roberts, 1995). El análisis de susceptibilidad magnética se realizó cada centímetro en todo el núcleo de sedimento, utilizando un dispositivo Bartington MS2 —Bartington Instruments, Witney Oxford, Reino Unido— en el Laboratorio de Paleoecología y Cambio Climático del Centro Universitario de la Costa Sur de la Universidad de Guadalajara. La composición elemental —Sr, U, Pb, As, Zn, Cu, Ni, Fe, Mn, Ti, Ca y K, expresado en concentración (ppm)— se determinó a través de fluorescencia de rayos X a cada centímetro en todo el núcleo de sedimento con un espectrómetro manual Thermo Scientific Niton XL3t en el Laboratorio de Geoquímica Ambiental del Instituto de Geología de la UNAM. Se determinó la pérdida progresiva por ignición con el fin de estimar el contenido de materia orgánica (MO) y carbonatos en los sedimentos, utilizando las relaciones lineales entre los valores de pérdida progresiva por ignición y el contenido de MO y carbonatos expresados en porcentaje (Dean, 1974; Heiri, Lotter y Lemcke, 2001). Este análisis se realizó cada centímetro en todo el núcleo de sedimento, tomando muestras de 0.5cm3 y mediante la pérdida de peso en las muestras al ser sometidos a altas temperaturas secuenciales (550 y 910°C). La interpretación en los cambios de porcentaje de pérdida progresiva por ignición está relacionada con eventos de mayor productividad y estabilización de la vegetación.
Para reconstruir la historia de la vegetación por medio del polen fósil, se realizó la extracción de palinomorfos modificada de Faergri e Iversen (1989). Para tal efecto se tomaron muestras de sedimento de un cm3 y se agregaron 2 pastillas de Lycopodium clavatum (13,875 lote del frasco) como esporas marcadoras para calcular la concentración de granos de polen por cm3. Las muestras se tiñeron con 2 gotas de safranina y posteriormente se montaron sobre una laminilla en gelatina glicerina para su posterior observación al microscopio. El conteo de palinomorfos —granos de polen de gimnospermas y angiospermas y esporas de pteridofitas sensu lato— se realizó con la ayuda de un microscopio Primo Star de Carl Zeiss con una magnificación de 400 aumentos. Con el fin de asegurar confiabilidad estadística (Maher, 1972) fueron contados los granos de polen hasta alcanzar un mínimo de 400 granos de polen y esporas por cada cm3.
La determinación de polen y esporas se llevó a cabo utilizando la colección de referencia palinológica del Laboratorio de Paleoecología y Cambio Climático de la Universidad de Guadalajara, claves palinológicas y bibliografía especializada. El porcentaje de palinomorfos fue calculado con base en la suma polínica de árboles, hierbas y pteridofitas (Bennett y Willis, 2001). Con el polen local se elaboraron los diagramas polínicos para conocer los cambios en la vegetación en el sitio de estudio. Para identificar la presencia de incendios forestales pasados y su relación con los cambios en la composición taxonómica y ambiental del sitio (Whitlock y Larsen, 2001), se realizó el conteo de micropartículas de carbón sobre las mismas muestras palinológicas siguiendo el método de Clark (1982); esta técnica consiste en el conteo del número de partículas de microcarbón proyectadas sobre puntos determinados en la laminilla palinológica. Los valores de concentración de microcarbón se presentan como el número de micropartículas de carbón por cm3.
Para estimar la cantidad de palinomorfos por unidad de volumen (granos de polen cm-3) se realizó el cálculo de la concentración polínica que brinda información paleoecológica de los cambios en la sedimentación del sitio (Faergri e Iversen, 1989). La tasa de acumulación polínica se calculó a partir de la concentración polínica que estima la cantidad de sedimento depositado en el tiempo y se expresa en cm año-1 (Birks y Birks, 1980). La riqueza palinológica, interpretada en términos de diversidad de plantas en el paisaje, se estimó por medio del análisis de rarefacción para comparar la riqueza de taxones entre las muestras (Birks y Line, 1992), es decir, estima la diversidad de tipos de polen en una muestra y la forma en que esta diversidad difiere entre las muestras. Para estimar el recambio de especies a través del tiempo se estimó la tasa de recambio de la vegetación; al promediar las tasas de vegetación de varios sitios se suavizan los cambios locales, lo que permite estimar la presencia de cambios sincrónicos o regionales (Grimm, 1987). El método utilizado mide la disimilitud entre las muestras adyacentes y divide esta medida por la diferencia temporal entre las muestras (Bennett, Boreham, Sharp y Switsur, 1992). Las zonas palinológicas se definieron utilizando el análisis de agrupamiento aglomerativo, jerárquico y estratigráficamente restringido, Constrained Cluster Analysis by Sum of Squares (CONISS) (Grimm, 1987). Este análisis representa los taxones con patrones similares de presencia a través de la secuencia estratigráfica (Birks y Birks, 1980). Con los porcentajes estimados de la suma palinológica se construyeron los diagramas polínicos, así como de concentración de microcarbón fósil. Los análisis mencionados anteriormente se realizaron con el programa Psimpoll versión 4.25 (Bennett, 2005).
Se realizó un análisis de componentes principales (ACP) con el fin de determinar posibles asociaciones o grupos en la composición taxonómica de cada una de las profundidades del sedimento. Para ello, se elaboró una matriz de datos de la composición taxonómica y las edades del sedimento. El ACP es un método de ordenación indirecto que tiene como principio encontrar una serie de combinaciones lineales de las variables originales que expliquen la mayor parte de la variación total con correlación nula (Ter Braak, 1986). Adicionalmente, con el fin de determinar la relación entre los patrones de la composición de taxones con la variación ambiental, se realizó un análisis de correspondencia canónica (ACC). El ACC es un método de ordenación directo que permite revelar las variables ambientales que mejor se correlacionan con los patrones observados en la composición de los taxones (Ter Braak, 1986). Las variables ambientales incluidas en este estudio fueron la susceptibilidad magnética, el porcentaje de carbonatos, el microcarbón fósil, el porcentaje de MO y el registro de elementos. Ambos análisis multivariados fueron llevados a cabo mediante el programa Canoco versión 4.5. (Lepš y Šmilauer, 2003)
ResultadosEl análisis de 14C realizado sobre la base del núcleo TLP-N2 de 37cm de profundidad registró una fecha de 720 años antes del presente (1230 dC). El modelo de edad/profundidad (fig. 2) se obtuvo mediante interpolación lineal a partir de las dataciones obtenidas de las profundidades 3, 15 y 37cm (tabla 1), lo que permitió estimar la tasa de sedimentación promedio en 19 años/cm.

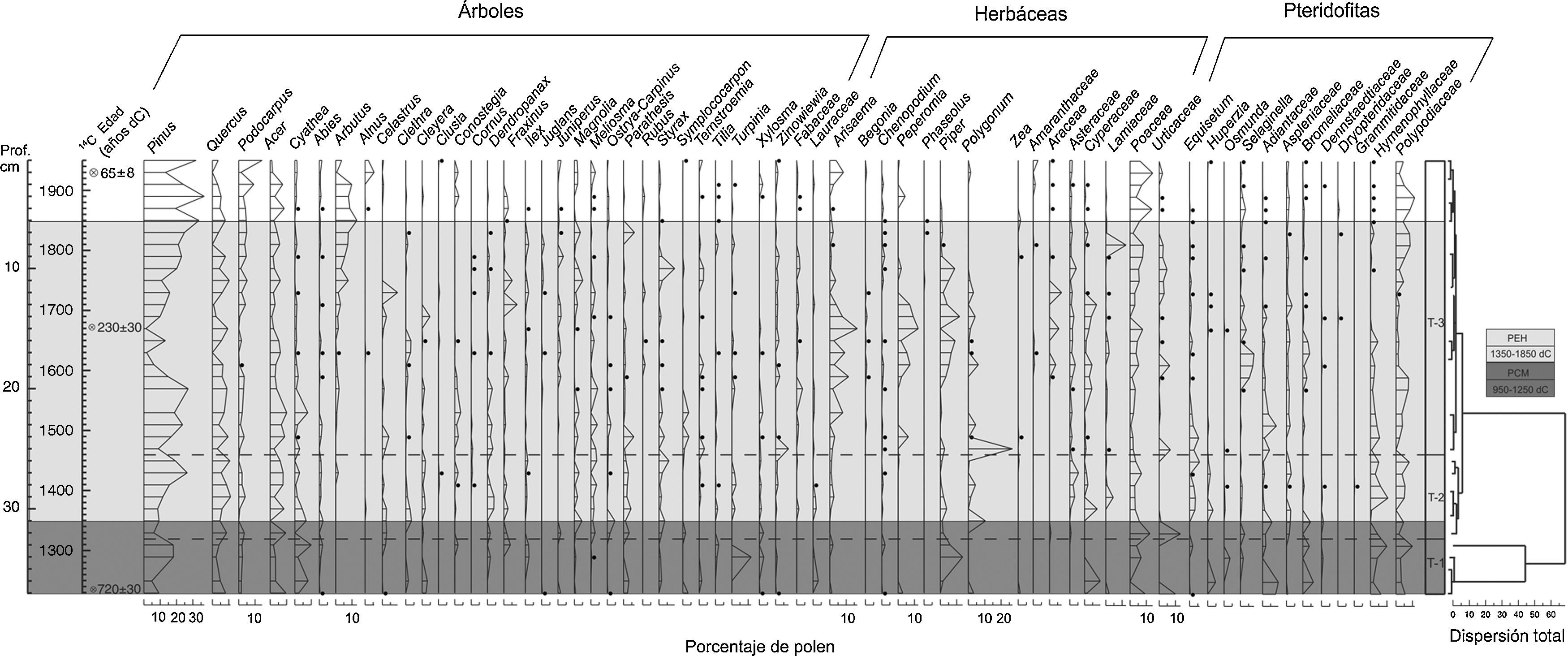
El análisis palinológico de las muestras extraídas del núcleo TLP-N2 de 37cm de profundidad permitió la identificación de 82 taxones, y el análisis de agrupamiento CONISS (Grimm, 1987) separó la secuencia de acumulación de sedimentos en 3 zonas palinológicas (fig. 3) que se describen a continuación:

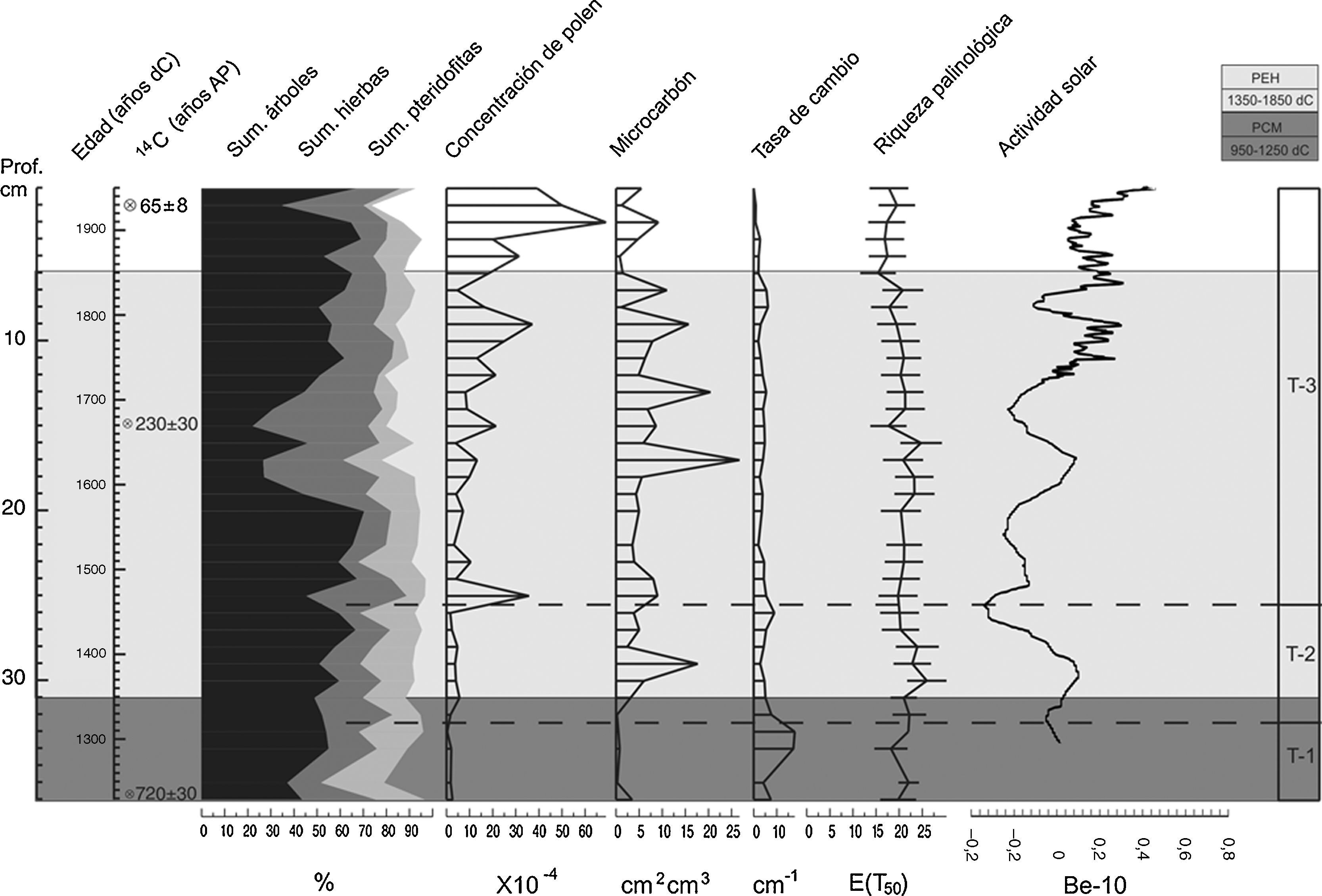
Los taxones arbóreos con mayor porcentaje fueron Pinus, Quercus, Acer, Cyathea y Turpinia (10-20%) (fig. 3). Otros taxones menos abundantes, pero que sobresalen por su importancia ecológica fueron Podocarpus, Abies, Clethra, Cleyera, Parathesis, Styrax, Symplococarpon, Ternstroemia, Xylosma y la familia Lauraceae, que se mantienen presentes en este periodo con porcentajes bajos (∼5%) (fig. 3). El estrato herbáceo, en relación con el anterior, presentó los porcentajes más bajos en esta zona. Las herbáceas más abundantes fueron Piper, Poaceae, Cyperaceae y Urticaceae (10-15%); en el grupo de las pteridofitas se registró a Adiantaceae, Hymenophyllaceae y Polypodiaceae como los taxones más abundantes con el 10%, en tanto que Equisetum, Huperzia, Osmunda, Selaginella, Bromeliaceae, Dennstaedtiaceae y Ophioglossaceae presentaron los porcentajes más bajos. La proporción de quebradas, tapadas y constreñidas fue muy elevada en todas las muestras, alcanzando valores hasta del 10%. Además, se registra la presencia de Zea desde 1230 dC (fig. 3). La concentración de polen en esta zona es considerablemente baja (∼5%), la cual muestra un decremento en 1297 dC. Por su parte, la riqueza palinológica tiene valores bajos (15-20%) en la mayor parte de esta zona y presenta un decremento en 1297 dC, mientras que la tasa de cambio registra el valor más elevado de toda la secuencia entre 1297-1319 dC. La riqueza palinológica muestra un incremento en 1364 dC y, finalmente, la concentración de microcarbón es considerablemente baja en esta zona con valores menores a 5% (fig. 4).
. http://www.ncdc.noaa.gov/paleo/pubs/crowley.html. La división de las zonas se basó en el análisis CONISS. PCM: periodo cálido medieval; PEH: pequeña edad de hielo.")
Síntesis de datos palinológicos. Se presenta la suma de todos los taxones: árboles-arbustos en color negro, herbáceas en color gris y pteridofitas en color gris claro, la concentración total de polen, la concentración de partículas de microcarbón, la tasa de cambio y la riqueza palinológica del núcleo TLP-N2. Adicionalmente se ejemplifica la variación solar con los datos de Be; disponible en: Crowley (2000). http://www.ncdc.noaa.gov/paleo/pubs/crowley.html. La división de las zonas se basó en el análisis CONISS. PCM: periodo cálido medieval; PEH: pequeña edad de hielo.
En esta zona el BMM presentó un ligero aumento en la tasa de cambio en 1453 dC que coincide con la disminución del componente arbóreo y el aumento de taxones herbáceos y pteridofitas (fig. 4). La riqueza palinológica registra un pico en 1386 dC (fig. 4). En el componente arbóreo, los taxones dominantes estuvieron constituidos básicamente por Pinus, Quercus, Acer y Cyathea (10-35%). Otros taxones frecuentes, pero en menor proporción fueron: Podocarpus, Abies, Celastrus, Clethra, Cleyera, Dendropanax, Fraxinus, Magnolia, Meliosma, Parathesis y Styrax. El componente herbáceo dominante está representado por los taxones de Piper, Polygonum, Asteraceae, Cyperaceae, Poaceae y Urticaceae (10-15%). El polen de Lamiaceae, Euphorbiaceae y Liliaceae presentan menor porcentaje en esta zona (5%). En 1475 dC se registró por segunda vez la presencia de polen de Zea. La pteridoflora dominante se representa por taxones de Equisetum, Polypodiaceae, Hymenophyllaceae, Aspleniaceae, Adiantaceae y Bromeliaceae (fig. 3). La concentración de partículas de microcarbón registra un pico de concentración en 1408 dC (fig. 4).
Zona T-3: (25.5-0cm; 1497-2012)En esta zona la concentración de polen fue más alta en comparación con las 2 zonas anteriores. El primer pico de riqueza palinológica se produjo en 1497, el segundo en 1802, el tercero y más elevado de todos, se presentó en 1885 dC (fig. 4). En el estrato arbóreo se observó un registro irregular de Pinus, manteniendo altos porcentajes en esta zona con valores de 20-35%. Asimismo, Quercus, Podocarpus, Acer y Arbutus se establecen como codominantes, alcanzando valores mayores al 10% (fig. 3). Taxones como Cyathea, Celastrus, Clethra, Cleyera, Conostegia, Fraxinus, Magnolia, Meliosma, Parathesis y Styrax se mantienen presentes con porcentajes≥10%. Además, se evidencia el aumento de Alnus a partir de 1885 dC con valores del 5% hasta el presente. Otros elementos arbóreos poco representados en esta zona son: Clusia, Cornus, Dendropanax, Ilex, Juglans, Juniperus, Ostrya-Carpinus, Symplococarpon, Ternstroemia, Tilia, Turpinia, Xylosma y Zinowiewia (fig. 3). El estrato herbáceo se incrementó considerablemente con la presencia de Arisaema, Peperomia, Piper, Polygonum, Cyperaceae, Lamiaceae y Poaceae. Se registra nuevamente polen de Zea en 1802 y 1844 dC. Sin embargo, se encuentran en menor proporción los taxones herbáceos de Begonia, Chenopodium, Plantago, Reseda, Amaranthaceae, Araceae, Asteraceae, Euphorbiaceae, Liliaceae, Melastomataceae, Solanaceae y Urticaceae. Además, se presentó una disminución importante de la pteridoflora con valores menores al 5%, únicamente Polypodiaceae permaneció con porcentajes mayores al 10% (fig. 3). En relación con la suma de árboles, herbáceas y pteridofitas (fig. 4), se encontró una dominancia de los taxones de árboles y arbustos desde 1520 hasta 1609 dC; posteriormente se registró una disminución considerable entre 1653-1720 dC, recuperándose a partir de 1802 dC. Al disminuir los taxones arbóreos y pteridofitas, las herbáceas aumentan sus porcentajes. La concentración de partículas de microcarbón en esta zona fue más elevada que en las 2 anteriores, presentándose un incremento en 1675, 1748, 1802, 1830 y 1885 dC (fig. 4).
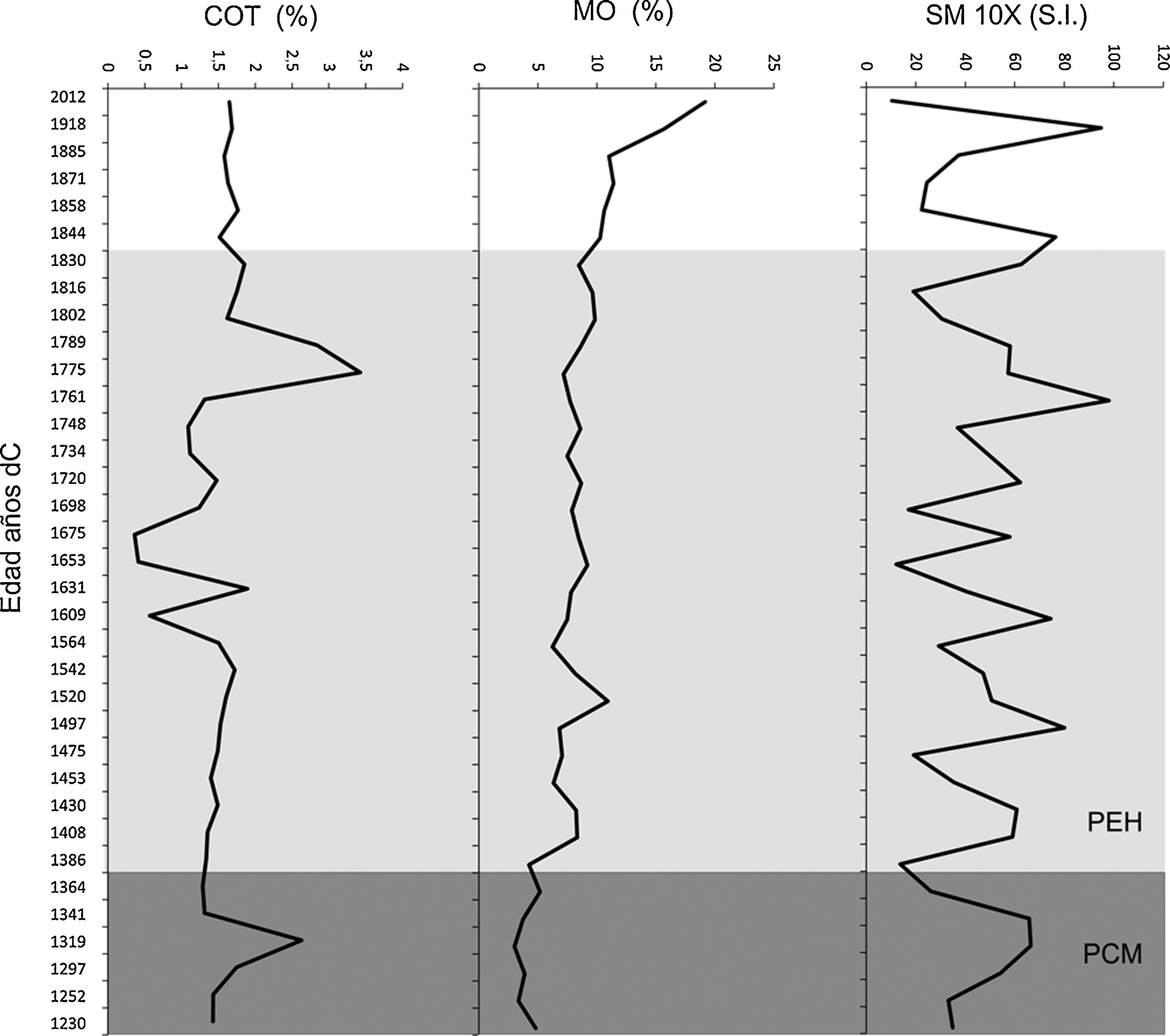
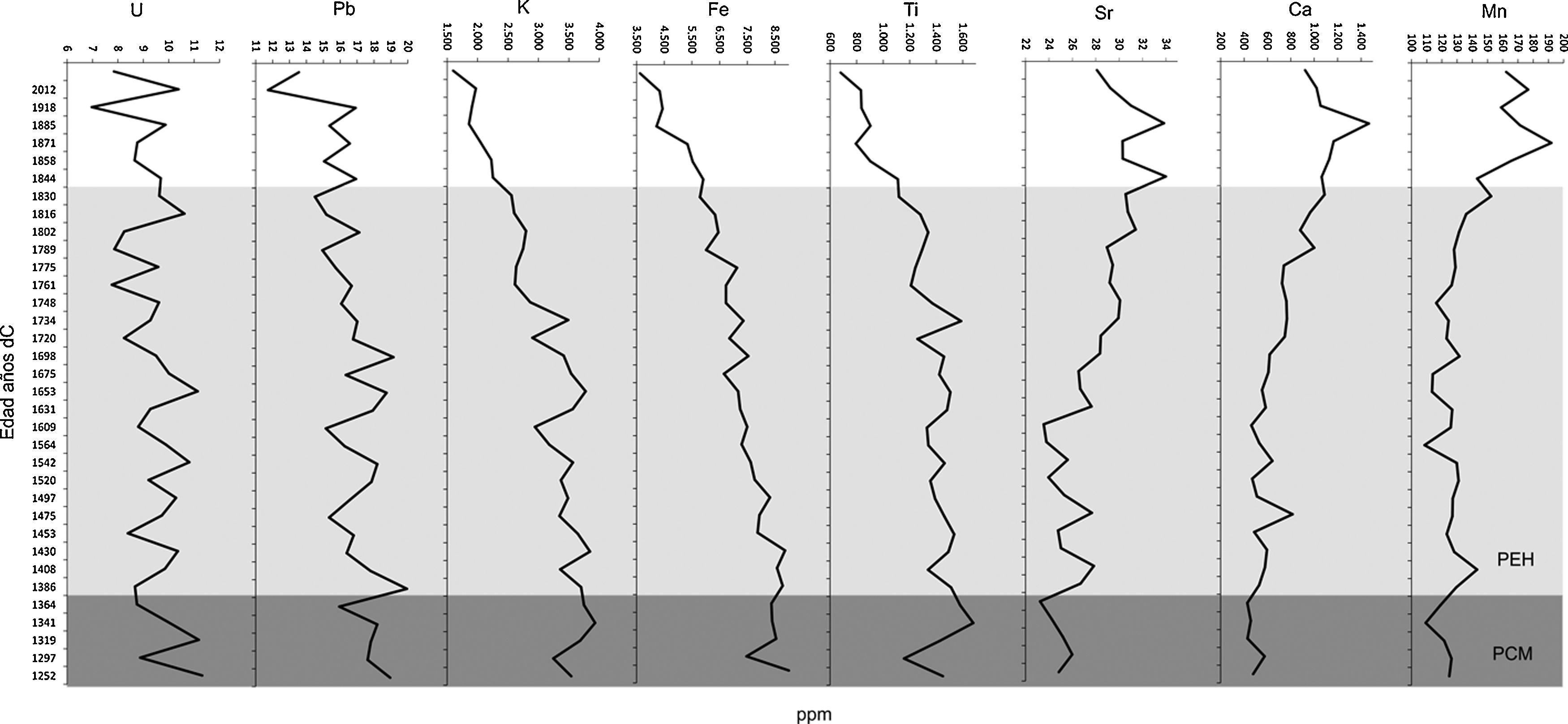
Análisis de la variación ambientalLos valores de la susceptibilidad magnética fueron muy variables a lo largo de la secuencia; los valores más bajos se registraron en 1386 dC en la zona T-2 y en 1653 dC en la zona T-3, mientras que los mayores se presentaron en 1761 y 1918 dC en la zona T-3 (fig. 5). De la misma manera, el porcentaje de la MO fue muy bajo en la base de la secuencia (5%) con un incremento gradual en 1520 (∼12%) con una disminución drástica en 1564 dC (5%). A partir de este mismo año, el valor de la MO se incrementa hasta la alcanzar los valores cercanos al 19% en el presente (fig. 5). Los valores más bajos de MO estuvieron entre 3-7% registrados entre 1609 y 1675 dC (fig. 5). El contenido de carbonatos mostró una tendencia heterogénea presentando 3 picos de mayor intensidad en 1319 (∼25%), 1631 (∼20%) y en 1775 dC (∼30%). Elementos como el Mn y el Ca presentaron también valores bajos en la base de la secuencia, incrementándose gradualmente hasta alcanzar valores de ∼18ppm en Mn en 1858 dC y el Ca alcanzó el mayor valor en 1871 dC. Por el contrario, elementos como K, Fe y Zn presentaron valores mayores en la base de la secuencia (35, 90 y 50ppm, respectivamente) observándose un decremento en la parte superior del registro estratigráfico. Otros elementos como Sr, U y Pb también presentaron valores irregulares a lo largo de la secuencia, mientras que Ti registró valores bajos (12-14ppm) en la base de la secuencia y gradualmente aumentó su concentración en la zona 3 (fig. 6).
, materia orgánica (MO) y carbonatos (COT). La división de las zonas se basó en el análisis CONISS. PCM: periodo cálido medieval; PEH: la pequeña edad de hielo.")
; uranio (U); plomo (Pb); potasio (K); hierro (Fe); titanio (Ti); manganeso (Mn) y calcio (Ca). La división de las zonas se basó en el análisis CONISS. PCM: periodo cálido medieval; PEH: pequeña edad de hielo.")
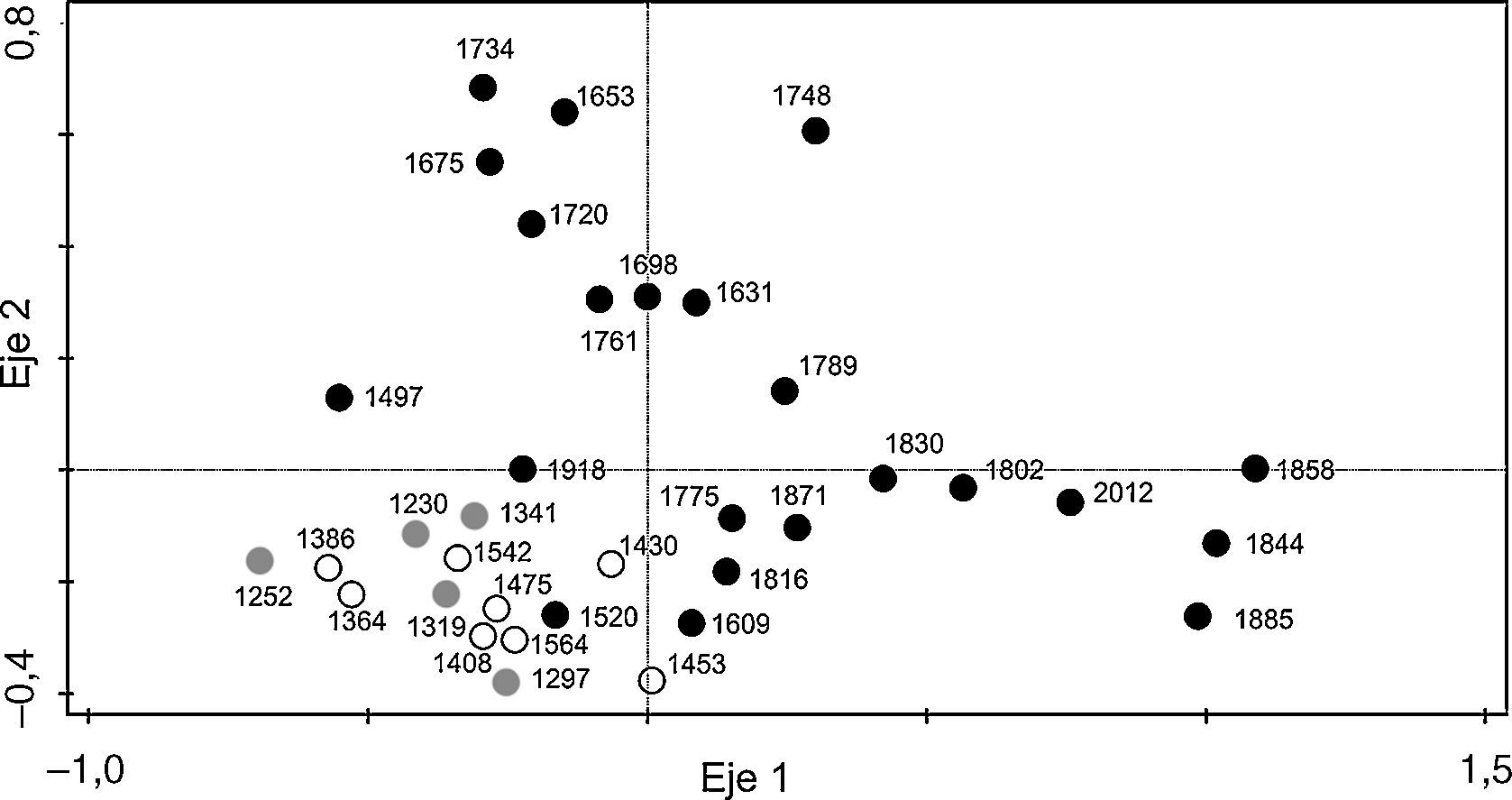
El ACP reveló una importante separación de las edades más recientes con las más antiguas en el eje 1 (eigenvalue=0.2036; un 20.36% del total de la variación), agrupándose juntas las edades más antiguas que corresponden con la zona T-1 (1230-1341 dC) y las de la zona T-2 (1341-1475 dC). Las edades recientes, que corresponden con la zona T-3 (1497-2012 dC), se mostraron más dispersas a lo largo del eje 2 (eigenvalue=0.0877) (fig. 7).
; los círculos blancos las profundidades intermedias (T-2) y los círculos negros representan las profundidades más recientes (T-3).")
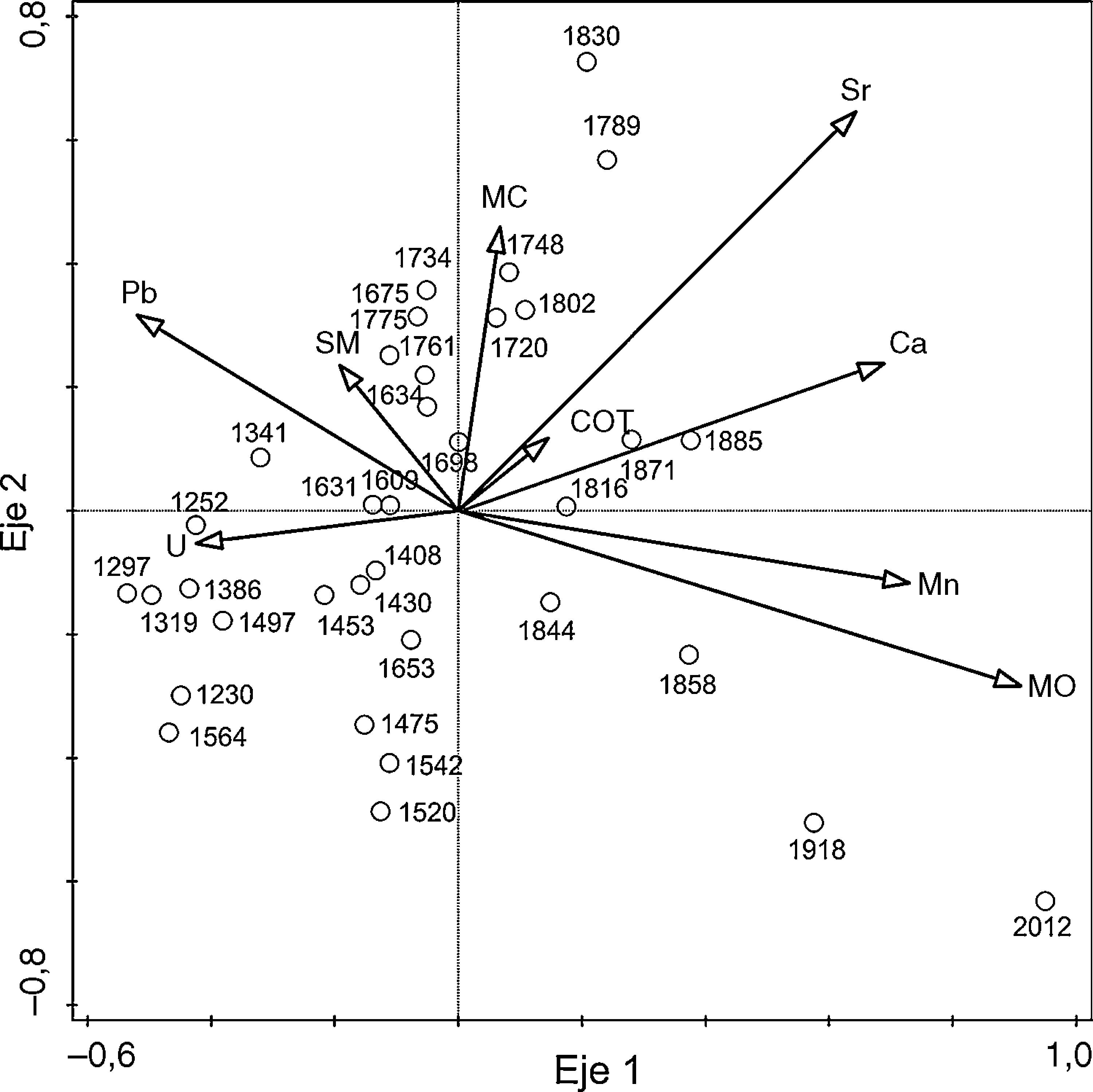
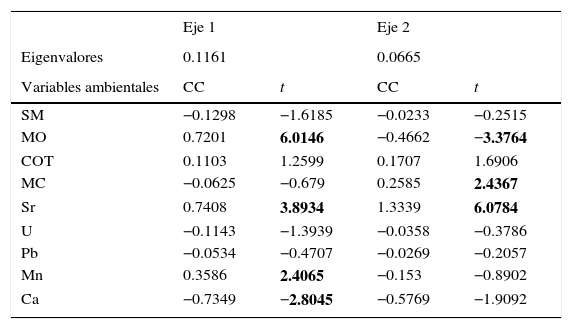
El ACC explicó el 33.1% del total de la varianza. En el eje 1 las variables más importantes fueron la MO, el Sr, el Mn y el Ca (tabla 2), cuyos vectores se asocian con las edades recientes de 1720 dC al presente (fig. 8). Mientras que en el eje 2 la MO, el microcarbón y el Sr resultaron las variables más importantes (tabla 2; fig. 8) asociadas a las edades más antiguas (1698-1230 dC).
Análisis de correspondencia canónica para las profundidades de núcleo de sedimento TLP-N2 dentro del BMM. Coeficientes de correlación (CC) de variables ambientales y valores de t (t); los números en negritas representan el valor crítico de 2.1 (df≥α=0.05).
| Eje 1 | Eje 2 | |||
|---|---|---|---|---|
| Eigenvalores | 0.1161 | 0.0665 | ||
| Variables ambientales | CC | t | CC | t |
| SM | −0.1298 | −1.6185 | −0.0233 | −0.2515 |
| MO | 0.7201 | 6.0146 | −0.4662 | −3.3764 |
| COT | 0.1103 | 1.2599 | 0.1707 | 1.6906 |
| MC | −0.0625 | −0.679 | 0.2585 | 2.4367 |
| Sr | 0.7408 | 3.8934 | 1.3339 | 6.0784 |
| U | −0.1143 | −1.3939 | −0.0358 | −0.3786 |
| Pb | −0.0534 | −0.4707 | −0.0269 | −0.2057 |
| Mn | 0.3586 | 2.4065 | −0.153 | −0.8902 |
| Ca | −0.7349 | −2.8045 | −0.5769 | −1.9092 |
; materia orgánica (MO); susceptibilidad magnética (SM); carbonatos (COT); calcio (Ca); estroncio (Sr); manganeso (Mn); plomo (Pb) y uranio (U).")
Análisis de correspondencia canónica de las edades del núcleo de sedimento TLP-N2. Las flechas indican las variables ambientales representadas por micropartículas de carbón (MC); materia orgánica (MO); susceptibilidad magnética (SM); carbonatos (COT); calcio (Ca); estroncio (Sr); manganeso (Mn); plomo (Pb) y uranio (U).
Durante el análisis palinológico del núcleo de sedimento TLP-N2 se identificaron 82 taxones, incluyendo los poco frecuentes y los raros por su importancia ecológica en el funcionamiento del BMM (Rzedowski, 2006). Pinus, Acer, Podocarpus y Quercus dominaron el estrato arbóreo, mientras que Clusia, Magnolia, Ostrya y Meliosma fueron poco abundantes en toda la secuencia. Pinus se presentó como un elemento dominante a lo largo de la secuencia con algunas fluctuaciones marcadas que coinciden con algunos cambios en la composición taxonómica y ambiental.
Los resultados encontrados sugieren que la composición florística del BMM con relicto de Acer ha fluctuado a través del último milenio. Las 3 zonas delimitadas en el núcleo de sedimento se caracterizan por una alternancia de taxones dominantes; estas fluctuaciones florísticas responden a la variación temporal del clima, así como al régimen de perturbación observado. De 1230 a 1341 dC, a pesar de las bajas concentraciones de polen relacionadas al tipo de sedimento (arenas) y su escasa preservación, los taxones dominantes como Pinus, Quercus, Podocarpus, Acer y Cyathea reflejan la existencia de este tipo de vegetación. La abundancia de taxones arbóreos frecuentes en el BMM como Cleyera, Clethra, Dendropanax, Parathesis, Turpinia y Xylosma (Gual-Díaz y Rendón-Correa, 2014) sugieren que este tipo de vegetación está presente desde 1230 dC en el área de estudio. Los resultados encontrados sugieren, además, la presencia de altos porcentajes en Hymenophyllaceae y Polypodiaceae en 1230 dC, seguidas por Adiantaceae y Bromeliaceae en 1252 dC. Abundancias similares de estos taxones son registradas en los mismos periodos para la sierra de Manantlán (Figueroa-Rangel et al., 2010), así como para la sierra Madre Occidental en el estado de Sonora (Ortega-Rosas et al., 2008). Las pteridofitas también estuvieron bien representadas en el BMM del área de estudio, y en la actualidad se estima que en México la diversidad florística de este ecosistema es de más de 1,000 especies (Martínez-Salas y Ramos, 2014), de las cuales 295 son consideradas exclusivas del BMM (Gual-Díaz y Rendón-Correa, 2014; Rzedowski, 1996). Este patrón de abundancia en la pteridoflora se mantiene hasta 1653 dC. Sin embargo, a partir de 1761 dC decrece al mismo tiempo que las herbáceas, como Poaceae, aumentan como resultado de la transición hacia un tipo de vegetación característico de claros grandes que pudieron ser inducidos por fluctuaciones del clima o causados por agentes de perturbación naturales o antropogénicos (Almeida, Hooghiemstra, Cleef y van Geel, 2005; Berrio, Arbelaez, Duivenvoorden, Cleef y Hooghiemstra, 2003). Un resultado interesante de este estudio indica que el registro de polen de maíz a partir de 1230 dC sugiere la posibilidad de ocupación humana en esta región. Por otro lado, los bajos valores de microcarbón —los más bajos de toda la secuencia analizada— muestran que es muy posible que en el área de estudio no ocurrieron incendios severos de 1230 a 1341 dC. Otros indicadores ambientales como la susceptibilidad magnética con valores altos sugieren que en este periodo ocurrieron elevadas tasas de erosión en el área de estudio. Trabajos previos llevados a cabo en el lago de La Luna, en el centro de México, así como en la laguna de Juanacatlán, esta última cercana al sitio de estudio, indican para este periodo condiciones de mayor humedad, que marcan el final de la anomalía del PCM (Metcalfe et al., 2010; Cuna et al., 2014).
De 1341 a 1497 dC la abundancia de taxones característicos del BMM refleja su florecimiento en este periodo. Condiciones similares fueron registradas durante este periodo en el bosque mesófilo de la sierra de Manantlán (Figueroa-Rangel et al., 2010, 2011). Por su parte, Acer se encontró presente en toda la secuencia del núcleo, aunque con algunas fluctuaciones, manteniendo su registro hasta el presente. En la actualidad este género muestra una distribución disyunta en poblaciones muy separadas, principalmente en cañadas protegidas y aisladas de bosque mesófilo en México y Guatemala (Jardel-Peláez, Cuevas, Santiago, Muñoz y Aragón, 1996; Vargas-Rodríguez, 2011; Vázquez-García et al., 2000). Estudios florísticos del sitio en la actualidad muestran que Acer saccharum subsp. skutchii se encuentra asociada a géneros de árboles templados como Podocarpus, el cual ocurre como un taxón característico de América del Sur, donde alcanza su mayor diversidad y se conoce su distribución más septentrional en los BMM del noreste de México (Contreras-Medina, Luna-Vega y Alcántara-Ayala, 2006). La presencia de taxones herbáceos como Arisaema, Peperomia, Piper, Polygonum, Selaginella, Zea, Cyperaceae y Amaranthaceae sugieren la posibilidad que la apertura de claros para la agricultura fue muy frecuente en el área de estudio. Estos cambios ambientales se registran también en los indicadores geoquímicos con valores altos de susceptibilidad magnética, la cual está asociada con mayor aporte de materiales terrígenos originados por la erosión del suelo, al igual que las altas concentraciones de Fe. Por su parte, las partículas de microcarbón registraron por primera vez un aumento en 1408 dC, lo que sugiere la frecuencia de incendios severos en la región del BMM, originando fluctuaciones en la vegetación como la disminución del polen de Pinus, Quercus, Acer y Cyathea.
A partir de 1497 dC se registra un cambio en la composición de la vegetación, con una dominancia de más del 30% en la suma polínica de Pinus y a partir de 1761 dC se encontró a Arbutus codominando con más del 10%, mientras que Quercus y Acer también se mantienen como abundantes. Este grupo de elementos holárticos se caracteriza por su habilidad para establecerse en etapas tempranas de la sucesión y, generalmente, no se establecen bajo sombra densa (Jardel-Peláez, Ezcurra, Cuevas-Guzmán, Santiago-Pérez y Cruz-Cerda, 2004), lo que sugiere un cambio en la apertura del dosel. En este sentido, la composición florística puede variar de acuerdo con las características estructurales y de distribución en los elementos del bosque mesófilo, llegando a contener una gran diversidad de especies, pero con pocos individuos, tal es el caso de Fraxinus, Magnolia, Meliosma, Parathesis, Styrax y Xylosma que presentaron bajos porcentajes en el espectro polínico, sin embargo, tienen buena distribución en las sierras meridionales, como las del occidente de México (Alcántara, Luna y Velázquez, 2002). En el componente herbáceo, se encontró una dominancia de Poaceae, un taxón que se relaciona con aperturas de claros y perturbación antropogénica (Bush, 2002; Lozano-García et al., 2010). Además, la abundancia de Piper, que se ha documentado como característico de ambientes secos y de alteraciones (Goman y Byrne, 1998), refleja que durante este periodo el BMM enfrentaba condiciones de mayor sequía, originando cambios en los taxones dominantes. Por otro lado, la presencia de las pteridofitas se ve drásticamente disminuida con excepción de Polypodiaceae que se mantiene presente con porcentajes menores al 10%; en particular este grupo es resistente a la desecación, por lo que se puede encontrar en zonas con poca humedad (Hietz y Briones, 1998), lo cual puede explicar su abundancia junto con Poaceae. Esto implica que el cambio en las comunidades de vegetación herbáceas se haya visto favorecido por los frecuentes incendios reflejados en las altas concentraciones de microcarbón a partir de 1675 dC al presente. Una característica importante de este periodo es el decremento en las concentraciones de Ti, Fe y K, con los subsecuentes aumentos de Ca y Mn, lo que podría estar reflejando condiciones de menor humedad (Sosa-Nájera et al., 2010).
Respuesta del ecosistema de montaña ante la variabilidad climáticaLa variabilidad climática del último milenio se atribuye a las anomalías del PCM y la PEH, esta última ha sido identificada en varios registros paleoclimáticos como un periodo de enfriamiento con alta variabilidad temporal (Lozano-García et al., 2007). La comparación del registro palinológico y ambiental del BMM se asocia con los cambios en la actividad solar del isótopo cosmogénico 10Be durante los últimos 700 años, lo cual indica que los cambios ambientales están relacionados con este forzamiento global, sin embargo, no está muy claro cómo actúa este mecanismo, por lo que se requiere más investigación en este campo.
Registros históricos en México revelan eventos de sequías entre el siglo xiv y el xix (Contreras-Servín, 2005; Cuna et al., 2014; Florescano, Sancho y Pérez, 1980; Jones y Mann, 2004). Reconstrucciones previas de diferentes indicadores fósiles han detectado que el cambio climático ha ocurrido a gran escala con cambios de temperatura y precipitación durante el último milenio (Cerano-Paredes et al., 2013; Lozano-García et al., 2007; Metcalfe, 1995; O’Hara y Metcalfe, 1997; Therrell, Stahle y Soto, 2004). Estos cambios fueron detectados en los registros de isótopos de oxígeno de un núcleo de sedimento tomado en Aguada X¿caamal en Yucatán (Hodell et al., 2005), en los datos de polen y diatomeas de lago Verde del centro-oeste de México (Lozano-García et al., 2007), en el registro de Ti en el lago Santa María del Oro en Nayarit (Sosa-Nájera et al., 2010), en los registros de anillos de crecimiento de árboles en la Reserva de la Biosfera Sierra de Manantlán (Cerano et al., 2013), en El Parque Nacional Pico de Tancítaro en Michoacán (Cerano-Paredes et al., 2014), además, en registros de Ti en los sedimentos laminados de la laguna de Juanacatlán (Metcalfe et al., 2010) y en los datos de palinomorfos, diatomeas y restos de cladóceros en la laguna de La Luna del Volcán Nevado de Toluca (Cuna et al., 2014).
Otras evidencias que aportan información al respecto son la disminución en las concentraciones de Ti, que se relacionan con reducción en la humedad. Los incendios que están estrechamente vinculados con la variabilidad climática y las actividades antropogénicas (Gavin et al., 2007), se registraron durante 1675-1885 dC, que coinciden durante la PEH, lo que significa que los paleofuegos pueden responder a las transiciones humanas y a la respuesta de la vegetación ante el clima. El régimen de incendios durante la PEH registrados en el BMM se caracteriza por intervalos más largos e intensos a los 1675, 1748, 1802 y 1830 dC coincidiendo durante los periodos más secos registrados en la secuencia; esto sugiere que el impacto del fuego ha sido un elemento esencial en la dinámica y recambio de taxones reflejado en la conformación del paisaje actual. Estos resultados muestran que durante la PEH la vegetación del BMM respondió a los cambios en la variabilidad climática derivada de estos forzamientos.
Dinámica ambiental y recambio de taxonesEl ACP agrupó las profundidades más antiguas incluyendo algunas de las más recientes; estas últimas se caracterizan por mayor abundancia de taxones leñosos y arbustivos. Este patrón es similar al observado en el ACC donde los sedimentos más recientes muestran alta relación con la MO, Sr, Mg y Ca. Esta variación ambiental sugiere que el BMM del sitio de estudio responde como un ecosistema sensible ante las fluctuaciones naturales del clima, con expansiones en épocas con aumento de humedad y contracciones en periodos prolongados de sequías (Figueroa-Rangel et al., 2010), por lo que se han adaptado a perturbaciones naturales a pequeña escala a través del tiempo (Cole, Bhagwat y Willis, 2014).
Otro evento de menor humedad ocurrió en 1430 y 1497 dC cuando se observa un incremento en el registro de elementos traza como Fe, K y Pb. Las altas concentraciones de algunos de estos elementos están relacionadas con factores climáticos como las sequías (Tipping, Smith, Lawlor, Hughes y Stevens, 2003). Durante estos periodos se redujeron la mayoría de los taxones leñosos como Pinus, Quercus, Podocarpus, Acer, Abies, Magnolia, Styrax y Ternstroemia, al igual que las pteridofitas como Adiantaceae, Aspleniaceae, Bromeliaceae e Hymenophyllaceae; en contraste ocurre un aumento en las herbáceas, por ejemplo, Polygonum, Asteraceae y Poaceae. En la mayoría de las comunidades el cambio en la composición de especies tiende a ser impulsada por los cambios en el clima, que también afectan la capacidad de recuperación del bosque (Brown, Morgan, Parody y Haskell, 2001; Figueroa-Rangel et al., 2011). En este registro se observa que los cambios ambientales modifican temporalmente los taxones dominantes, pero mantienen una composición de especies relativamente estable a través del tiempo. Estas condiciones de sequía se asemejan con los detectados en el lago de Santa María del Oro en 1413 y en 1437 dC y en la laguna de Juanacatlán a partir de registros de Ti (Metcalfe et al., 2010; Sosa-Nájera et al., 2010); igualmente se han detectado periodos de sequías en registros dendrocronológicos en Nuevo León a partir de 1412 a 1420 dC y de 1439 a 1459 dC (Villanueva-Díaz et al., 2007), en Chihuahua-Sonora entre 1487-1496 dC, además, en Durango, Sonora y Sinaloa en 1451-1455 y de 1462-1465 dC (Villanueva-Díaz, Fulé, Cerano-Paredes, Estrada-Ávalos y Sánchez-Cohen, 2009), también al noreste de México en 1450 dC (Cerano-Paredes, Villanueva-Díaz, Valdez-Cepeda, Méndez-González y Constante-García, 2011). En registros de códices Aztecas se mencionan los años 1452-1454 dC como una sequía prolongada que históricamente ha sido identificado como un periodo muy crítico, ocasionando carencia de granos básicos y epidemias en la población (Therrell et al., 2004).
Una tendencia hacia condiciones más secas se desarrolló en 1564, 1609, 1698, 1748, 1816 y 1858 dC, con el valor más bajo de Ti de toda la secuencia sedimentaria; además, coincide con los valores más altos de Ca. Este patrón se hace evidente en los eventos de sequías severas y prolongadas registradas en el estudio dendroclimático de la sierra de Manantlán, una región relativamente cercana al BMM con relicto de Acer (Cerano-Paredes et al., 2013), indicando periodos de sequía con una presencia cíclica cada 55 años que coincide con los ciclos solares (Crowley, 2000). Documentos históricos señalan que en la época colonial las sequías coincidían con heladas y granizadas que provocaban desastres agrícolas y hambre, en particular durante las crisis de 1695, 1785 y en 1808-1809 dC (Florescano et al., 1980). Para el lago de Santa María del Oro en Nayarit se detectó una intensa sequía registrada en los valores más bajos de Ti de 1655-1670 dC durante el mínimo de Maunder (Sosa-Nájera et al., 2010); esta sequía documentada en Nayarit coincide con un cambio en el registro palinológico del BMM entre 1653-1675 dC, observando un cambio importante en la cobertura del dosel, con la disminución de todos los taxones leñosos, lo que sugiere un cambio en la cobertura vegetal. Las herbáceas respondieron a esta apertura del dosel, lo cual se deduce del aumento en los registros de los porcentajes de polen de Arisaema, Peperomia, Piper, Polygonum, Araceae, Asteraceae, Cyperaceae, Lamiaceae, Poaceae y Urticaceae. A partir de entonces la composición del BMM cambió en algunos de sus taxones leñosos dominantes, por ejemplo, Arbutus se convirtió en un taxón codominante junto con Pinus, lo cual nos indica que el recambio de taxones en el BMM con relicto de Acer ha sido muy dinámico.
La relación temporal en los cambios ambientales detectados en el BMM con relicto de Acer entre los registros de indicadores de alta resolución y los datos históricos muestran que los eventos climáticos extremos sucedidos en otras regiones del país también se presentaron en esta comunidad vegetal, con algunas fluctuaciones entre eventos de sequías y recambio de taxones para la región occidental de México. A partir de 1609 hasta 1775 dC se observa una clara tendencia en el recambio de taxones arbóreos característicos del BMM que abarca el periodo denominado PEH.
Respuesta de la vegetación a alteraciones antropogénicasLos pobladores de Talpa se han dedicado por varios siglos a la agricultura, prueba de ello es el polen de Zea, que es evidente desde 1230 dC; posteriormente a lo largo de la secuencia aparece en 1475, 1520, 1816 y 1858 dC. Esto sugiere que las actividades agrícolas en el área de estudio probablemente se han realizado desde el establecimiento de los primeros habitantes indígenas de la región y que se ha mantenido desde entonces. La evidencia paleoecológica revela que la disminución en los valores de MO registrados a los 1564 dC pudiera estar asociado con la llegada de los pobladores españoles en la zona de Talpa y se vea reflejado con la disminución en la productividad del bosque derivado de actividades agrícolas. Es posible que la disminución de taxones como Pinus, Quercus y Podocarpus entre 1653-1720 dC se deba a la oscilación climática conocida como PEH, lo que generó algunos cambios en el conjunto del BMM. Simultáneamente con la disminución de Pinus y Quercus se observa una correspondencia de aumento de polen de herbáceas de Poaceae, posiblemente causados por los incendios severos y periodos de extrema sequía. A partir de 1858 dC se observa también un aumento considerable de Pinus, lo cual podría también estar influenciado por la acción antrópica. Actualmente en el sur de México se atribuye la expansión de este taxón a la influencia de la actividad humana (Ramı¿rez-Marcial, González-Espinosa y Williams-Linera, 2001); conforme la actividad de deforestación aumenta con fines agrícolas Pinus y Arbutus se incrementan, lo cual ocurre de manera similar en este bosque.
En los últimos 65 años se registra una marcada disminución en todos los taxones del polen de Pinus, Quercus, Podocarpus, Acer, Arbutus y Fraxinus. En las últimas décadas, la evidencia palinológica sugiere que las actividades productivas han tenido un impacto mucho mayor en el BMM con relicto de Acer. Las posibilidades de conservación de este tipo de bosque se vuelven una necesidad apremiante dado que la mayoría de los BMM en México se han convertido en terrenos agrícolas, para ganadería, para el cultivo de café o de explotación forestal (Challenger, 1998; Rzedowski, 2006), por lo que todas estas actividades han contribuido a la fragmentación del BMM (Cayuela et al., 2006; Williams, Manson y Isunza, 2002); sin embargo, el establecimiento de reservas como estrategia de conservación puede ser viable para mantener la biodiversidad del BMM en su conjunto (Figueroa-Rangel et al., 2010; González-Espinosa et al., 2012; Vargas-Rodríguez, Platt, Vázquez-García y Boquín, 2010). Por ello se considera una buena opción para la preservación de la biodiversidad del BMM con relicto de Acer.
El registro paleoecológico para el BMM con relicto de Acer ha estado sujeto a cambios ambientales los últimos 720 años, lo que se evidencia por los registros de indicadores ambientales, así como por la dinámica en el recambio de taxones. Las comunidades vegetales se manifiestan en periodos alternados, como respuesta a condiciones húmedas y por la transición hacia condiciones más secas. Estas respuestas del BMM a los cambios ambientales están en función de las características locales y regionales —condiciones fisiográficas—, que se presentan como reorganizaciones temporales y mantienen la presencia de poblaciones relictas como Acer, Podocarpus y Cyathea en pequeños microrrefugios.
Los cambios ambientales también influyen en la frecuencia de incendios, los cuales son más frecuentes durante el periodo de la PEH; adicionalmente las actividades humanas tienen repercusiones en el recambio de taxones y en la composición de la vegetación. Por lo tanto, la heterogeneidad temporal en la abundancia de los distintos taxones del BMM pudo haber desempeñado un papel importante en la capacidad de recuperación después de una perturbación. El BMM actual —los últimos 230 años— parece estar dominado por taxones de Pinus, Quercus, Podocarpus y Arbutus, además de herbáceas de Poaceae, relacionados con actividades antropogénicas. Los resultados presentados documentan las evidencias de la respuesta del bosque mesófilo ante la variabilidad climática, lo cual nos permite obtener una perspectiva temporal del cambio ambiental, con la posibilidad de contribuir con el entendimiento del impacto de la biodiversidad en los bosques mesófilos del occidente de México. Adicionalmente, sugerimos el desarrollo de estudios paleoecológicos a escalas temporales mayores que los aquí presentados, con la finalidad de combinarlos con estudios filogeográficos que pudieran dar una mejor resolución a la dinámica de la vegetación para comprender las respuestas de estos ecosistemas ante los efectos del cambio climático aunado a las perturbaciones antropogénicas.
Agradecemos el apoyo del M. en C. Arturo Solís-Magallanes y a los estudiantes de IRNA y Agronomía del CUCSUR-Universidad de Guadalajara, durante los trabajos de perforación. Asimismo, un especial agradecimiento a la Dra. Susana Sosa-Nájera y M. en C. Esperanza Torres-Rodríguez por su asesoría en la extracción e identificación de Palinomorfos. Este trabajo fue realizado por medio del apoyo financiero de los proyectos: Conacyt CB-2008-106435 y Coecytjal 5-2010-875, y gracias a la beca otorgada por el Conacyt a la primera autora para la realización de sus estudios de posgrado.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.