



En los bosques templados australes la endozoocoría es determinante para muchas plantas con frutos carnosos, siendo las aves los mayores consumidores de frutos y dispersores de semillas. El zorzal austral Turdus falcklandii, es un ave residente que consume frutos de la pitra Myrceugenia planipes, incrementando la germinación de las semillas y reduciendo el tiempo requerido para la aparición del hipocótilo, lo que se traduce en ventajas competitivas para los nuevos individuos.
In the temperate austral forests endozoochory is crucial for many plants with fleshy fruits, birds being the biggest consumers of fruits and seed dispersers. The Austral thrush Turdus falcklandii is a resident bird that consumes fruits of Myrceugeia planipes, increasing seed germination and reducing the time required for the appearance of the hypocotyl, which translates into competitive advantages for new individuals.
Los vertebrados frugívoros, en especial las aves tienen generalmente un efecto positivo en la capacidad germinativa de las semillas de los frutos consumidos (Traveset, 1998). En particular, se ha señalado a los zorzales (Turdus spp.) como importantes aves frugívoras capaces de aumentar el porcentaje de germinación y disminuir el tiempo de emergencia del hipocótilo (Paulsen y Högstedt, 2002). En los bosques templados australes, uno de los principales dispersores de semillas es el zorzal austral Turdus falcklandii, ave endémica y residente que consume frutos y posteriormente defeca las semillas sin afectar la viabilidad y germinación de estas (Figueroa y Castro, 2002; Mora y Smith-Ramírez, 2016; Orellana et al., 2014). T. falcklandii entre otras características, tiene la capacidad de consumir frutos carnosos con tamaños mayores al tamaño de su pico (> 10mm), convirtiéndola en la principal ave dispersora de semillas (i.e., no las daña) de grandes frutos del bosque templado de Sudamérica (Amico y Aizen, 2005; Gho-Illanes, Smith-Ramírez, Vásquez y Díaz, 2015; Salvande, Figueroa y Armesto, 2011). Esta característica es compartida con el marsupial Dromiciops gliroides (Amico, Rodríguez-Cabal y Aizen, 2009).
El género Myrceugenia (Myrtaceae) es nativo de Sudamérica, distribuyéndose solo en el sureste de Brasil, el archipiélago de Juan Fernández en Chile (2 especies endémicas) y en los bosques templados australes de Chile y Argentina (Landrum, 1981). En estos últimos, la pitra Myrceugenia planipes es un árbol tolerante a la sombra, con requerimientos lumínicos de luz difusa menores en comparación a otras plantas del bosque. Es abundante en sitios húmedos al interior del bosque integrando el dosel intermedio y sotobosque del tipo forestal Siempreverde (Donoso, 2006). M. planipes se caracteriza por presentar una estrategia reproductiva de xenogamia obligada (Riveros, Umaña y Arroyo, 1996). Sus frutos son bayas de color violáceo de tamaño superior a otros frutos nativos (12.5mm y 800mg aproximadamente), presentan 4 a 7 semillas con una testa delgada y una pulpa carnosa comestible, la cual inhibe la germinación de las semillas (Figueroa y Castro, 2002). Este efecto inhibidor de la pulpa es determinante en la dinámica regenerativa de la planta, siendo indispensable para la germinación de las semillas un agente biótico que extraiga la pulpa de los frutos sin dañarlas (Figueroa y Castro, 2002).
En la isla de Chiloé, solo se ha visto un efecto neutro en la germinación de semillas de M. planipes posterior a su paso por el tracto digestivo de las aves frugívoras (T. falcklandii y Xolmis pyrope) (Figueroa y Castro, 2002). El objetivo del estudio es evaluar el efecto de la frugivoría de T. falcklandii en la germinación de semillas de M. planipes en el continente, respecto de las islas, ya que podrían existir diferencias en la germinación producto de la variación geográfica y las adaptaciones locales (Galloway, 1995).
En noviembre de 2008, se recolectaron 14 frutos de M. planipes de un bosque nativo cercano a la ciudad de Osorno, sur de Chile (40°54′33″ S, 72°51′57″ O). En esta época del año, tanto la oferta de recursos alimenticios animales como los frutos de otras especies de plantas es baja (Orellana et al., 2014). Los frutos maduros recolectados de 7 árboles fueron llevados intactos al laboratorio donde se les extrajo manualmente la pulpa. Las semillas encontradas en las deposiciones de T. falcklandii fueron obtenidas de 5 posaderos ubicados al interior del bosque. En total se obtuvieron 110 semillas, 55 extraídas de frutos y 55 de deposiciones. Las semillas fueron distribuidas en grupos de 5 en compartimientos independientes en bandejas de poliestireno, obteniéndose un promedio de semillas germinadas por bandeja. Todas las semillas fueron sembradas en sustrato de tierra obtenida del suelo del bosque y dispuestas en un invernadero, siendo regadas 2 veces por semana. El porcentaje y tiempo de germinación fue estimado como el número promedio de semillas germinadas por bandeja y el tiempo promedio en días transcurridos desde la siembra hasta la aparición del hipocótilo. Las diferencias en el número de semillas germinadas, el tiempo de germinación y la combinación de ambas variables fueron analizadas mediante un Anova de medidas repetidas. Posteriormente, se realizó una prueba de contrastes de Tukey. Los resultados son expresados como el promedio ± 1 error estándar.
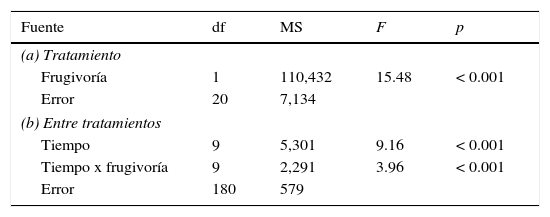
Hubo un efecto significativo de la frugivoría, el tiempo, y la interacción entre ambas variables sobre la germinación de las semillas de M. planipes (tabla 1). Las semillas extraídas de frutos y deposiciones comenzaron a germinar 18 días después de la siembra. La frugivoría de T. falcklandii incrementó la germinación de las semillas de M. planipes entre los días 18 y 20, posteriormente, la germinación de las semillas extraídas de frutos y deposiciones fue equivalente. Al final del experimento (día 27) no se observaron diferencias significativas en los porcentajes de germinación de las semillas extraídas de frutos y deposiciones, siendo estas entre un 42.9% y 48.2%, respectivamente (fig. 1).
Resumen de Anova de medidas repetidas para los efectos de la frugivoría de Turdus falcklandii en la germinación de semillas y el tiempo de germinación de Myrceugeia planipes.
| Fuente | df | MS | F | p |
|---|---|---|---|---|
| (a) Tratamiento | ||||
| Frugivoría | 1 | 110,432 | 15.48 | < 0.001 |
| Error | 20 | 7,134 | ||
| (b) Entre tratamientos | ||||
| Tiempo | 9 | 5,301 | 9.16 | < 0.001 |
| Tiempo x frugivoría | 9 | 2,291 | 3.96 | < 0.001 |
| Error | 180 | 579 | ||
df: grados de libertad; F: estadístico asociado al Anova; MS: cuadrado medio; p: probabilidad asociada.

El hecho de que T. falcklandii pueda consumir grandes frutos como los de M. planipes, sumado al efecto positivo en la germinación que tiene el paso de las semillas por el tracto digestivo y que sea un residente de estos bosques (i.e., no migra) permitiéndole consumir sincrónicamente los frutos de M. planipes, justo en el pico de su maduración, convierten a esta ave en el principal dispersor de sus semillas (Orellana et al., 2014; Robertson, Trass, Ladley y Kelly, 2006). El efecto positivo en la germinación producto de la escarificación mecánica y química producida en el aparato digestivo del ave es indispensable para frutos cuya pulpa tienen compuestos químicos (e.g., azúcares y glicoalcaloides) que inhiben la germinación (Samuels y Levey, 2005; Traveset, Riera y Mas, 2001). Estas características sugieren que M. planipes depende en gran parte de sus dispersores de semillas (Figueroa y Castro, 2002).
En el bosque templado austral de la isla de Chiloé, Figueroa y Castro (2002) encontraron que la ingesta de frutos de M. planipes por parte de las aves frugívoras no afecta la germinación de semillas, aunque sí germinan antes que las semillas no ingeridas. Nuestros resultados concuerdan con estos autores, aunque destacamos que en el continente, T. falcklandii acelera significativamente la germinación y emergencia del hipocótilo. No obstante, las diferencias en la germinación de semillas respecto de la isla de Chiloé no son tan marcadas (cerca del 10%) como para inferir grandes variaciones geográficas que las distingan. De hecho, las características morfológicas y fisiológicas en la germinación de M. planipes en toda su área distribución son similares (Donoso, 2006).
Concluimos que T. falcklandii es un dispersor legítimo de semillas de M. planipes, lo que se traduce en ventajas competitivas para los nuevos individuos al evitar los inhibidores químicos del fruto y poder escapar de patógenos y depredadores de semillas (Horvitz y Schemske, 1994; Vandelook, Van de Moer y Van Assche, 2008). Se sugiere que T. falcklandii cumpliría un rol clave en el éxito reproductivo de M. planipes del continente y en la sucesión secundaria del bosque templado austral de Chile.
El presente trabajo contó con el apoyo del Laboratorio de Ecología, Laboratorio de Vida Silvestre y la Dirección de Investigación y Posgrado de la Universidad de Los Lagos, Chile.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.