


The taxonomic identification of species in the genus Philornis Meinert (Diptera: Muscidae) has been based primarily upon analysis of morphological characters. However, relying solely on morphology has led to frequent species misidentification. This has been empirically demonstrated when specimens identified morphologically as Philornis seguyi have been reclassified as Philornis torquans based on DNA sequences of the second ribosomal internal transcribed spacer (ITS2), showing that the morphologic distinction between these 2 species is extremely challenging. Herein, new molecular data on specimens assigned to P. torquans and P. seguyi from the type locality of P. seguyi (Misiones province, Argentina) were analyzed. The ITS2 sequences obtained differed from those previously reported for Philornis, increasing to 3 the number of documented genotypes of flies morphologically indistinguishable from P. torquans. These results strongly suggest that subcutaneous Philornis spp. from southern South America could represent a species complex (P. torquans complex).
La identificación taxonómica de especies del género Philornis Meinert (Diptera: Muscidae) ha estado basada principalmente en el análisis de los caracteres morfológicos. Sin embargo, el uso exclusivo de información morfológica ha llevado con frecuencia a errores en la identificación de las especies. Esto fue demostrado de forma empírica cuando especímenes morfológicamente identificados como P. seguyi fueron reclasificados como P. torquans utilizando secuencias del segundo espaciador interno ribosomal transcrito (ITS2), lo que demuestra que la distinción morfológica entre estas 2 especies es extremadamente complicada. En el presente trabajo se analizaron nuevos datos moleculares de especímenes identificados como P. torquans y P. seguyi provenientes de la región reportada como localidad tipo para P. seguyi (provincia de Misiones, Argentina). Las secuencias de ITS2 obtenidas fueron diferentes a aquellas previamente reportadas para otras especies de Philornis y se incrementa a 3 el número de genotipos documentados de moscas morfológicamente indistinguibles asignadas a P. torquans. Estos resultados indican que los organismos de Philornis spp. subcutáneos del sur de Sudamérica podrían representar un complejo de especies (complejo P. torquans).
The genus Philornis Meinert (Diptera: Muscidae) comprises about 50 known species mainly distributed in the Neotropical region. Adults are free living, whereas larvae are strictly associated with bird nestlings, establishing different types of associations (coprophagous, semihaematophagous or subcutaneous; Dudaniec & Kleindorfer, 2006). The study of ecology and systematics of this group is highly relevant, since Philornis flies are parasites of some endangered species of conservation concern (e.g. Darwin finches parasitized by Philornis downsi Dodge & Aitken; Koop, Huber, Laverty, & Clayton, 2011), have the potential to be favored by global environmental change (Antoniazzi et al., 2010), and also are excellent models for the study of parasite ecology (Manzoli et al., 2013). To date, one semi-haematophagous (P. downsi) and 3 subcutaneous Philornis species (Philornis blanchardi García, Philornis torquans Nielsen and Philornis seguyi García) were reported for Argentina (Couri, Antoniazzi, Beldomenico, & Quiroga, 2009; Silvestri, Antoniazzi, Couri, Monje, & Beldomenico, 2011). The taxonomic identification of Philornis spp. is currently based mainly upon analysis of morphological characters of adult specimens (descriptions of larvae and pupae are available for a reduced number of species). According to the last published taxonomic review for Philornis spp. from Argentina (Couri et al., 2009), P. seguyi can be distinguished from P. torquans by the width of frons (females) and number of setae on the anterodorsal surface of the hind tibia (males). Differentiation between females from these 2 species could be considered clear (no overlap in values of distinctive characters), though this is not the case for males (the number of setae on the anterodorsal surface of the hind tibia is 3–4 in P. seguyi and 4–6 in P. torquans). As a consequence of this limitation, such identification approaches have been inaccurate, leading to frequent species misidentification (Monje et al., 2013).
A critical issue that obscures our knowledge on the systematics of Philornis spp. from southern South America is that holotypes of several species described in Argentina are lost (i.e., P. blanchardi, P. seguyi, Patitucci, Mulieri, Domínguez, & Mariluis, 2015), which complicates a proper taxonomic characterization of Philornis species in South America. A paradigmatic example of this problem is the case of P. seguyi. This species was described by García (1952) based on a single female originally housed at the collection of the “Administración Nacional de Laboratorios e Institutos de Salud Dr. Carlos G. Malbrán (ANLIS-Ent)” (Buenos Aires, Argentina), though is now considered lost (Patitucci et al., 2015). Moreover, the type locality of P. seguyi was only reported by García (1952) as “Misiones, Argentina” (Fig. 1A) without any specific geographic location. More recently, Couri, Rabuffetti, and Reboreda (2005) reported Philornis specimens in Magdalena (Buenos Aires province, Argentina), which females matched the description of the single female of P. seguyi, together with males that were assumed to belong to that species. In this work, the authors redescribed the species including both sexes, and reported it for several areas of Argentina.
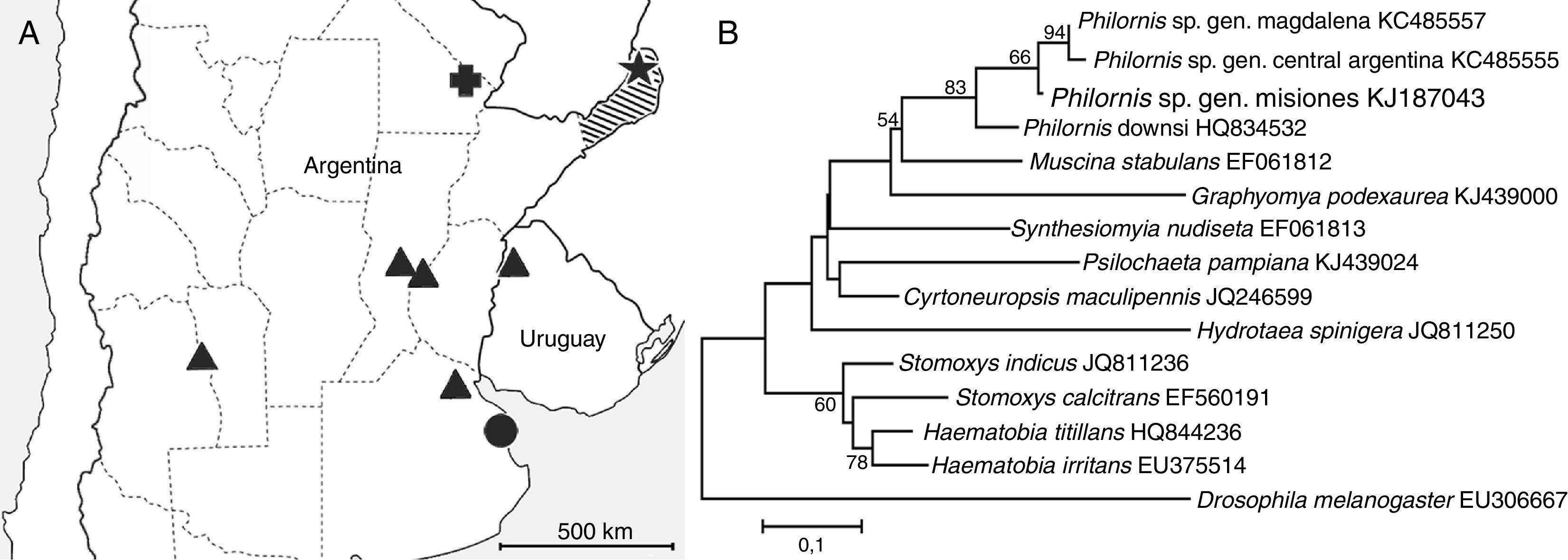
 Map of part of southern South America showing geographical distribution of Philornis spp. used in this study. ‘Philornis sp. genotype Misiones’ (star), ‘Philornis sp. genotype Central Argentina’ (triangle), ‘Philornis sp. genotype Magdalena’ (circle) and Philornis downsi (cross). Striped area: Misiones province. (B) Phylogenetic analysis of ITS2 sequences from adult specimens of Philornis species present in Argentina and closely related fly species. Bootstrap consensus tree was inferred by Maximum-likelihood method. The ITS2 sequence for Drosophila melanogaster (EU306667) was used as an outgroup.")
(A) Map of part of southern South America showing geographical distribution of Philornis spp. used in this study. ‘Philornis sp. genotype Misiones’ (star), ‘Philornis sp. genotype Central Argentina’ (triangle), ‘Philornis sp. genotype Magdalena’ (circle) and Philornis downsi (cross). Striped area: Misiones province. (B) Phylogenetic analysis of ITS2 sequences from adult specimens of Philornis species present in Argentina and closely related fly species. Bootstrap consensus tree was inferred by Maximum-likelihood method. The ITS2 sequence for Drosophila melanogaster (EU306667) was used as an outgroup.
Recently, Monje et al. (2013) analyzed specimens of Philornis from several localities from Argentina and Uruguay, and used the second internal transcribed spacer region (ITS2) of the ribosomal DNA as a molecular marker to differentiate adult specimens. The samples analyzed in this study were assigned to P. seguyi and P. torquans following Couri et al. (2009) work. ITS2 sequences were identical for all specimens from Uruguay and the central region of Argentina, though specimens from the southernmost locality (Magdalena city, samples provided by M.S. Couri) that were morphologically identified as P. seguyi had a genetic divergence of 5.9% compared to the former ones. Of the localities studied by Monje et al. (2013), Magdalena was one of the most distinctive, both in terms of distance and habitat, from the type locality of P. seguyi (Misiones province, Fig. 1A), suggesting that the specimens from Magdalena could not represent P. seguyi. Because the morphology of this particular group of flies may sometimes lead to species misidentification, here we refer to the Philornis genotypes identified by Monje et al. (2013) as the ‘Philornis sp. genotype Central Argentina’ and the ‘Philornis sp. genotype Magdalena’.
Herein, new material from the Misiones province mentioned by García (1952) as type locality of P. seguyi were analyzed to increase the number of ITS2 sequences analyzed by Monje et al. (2013), and to contribute to the reconstruction of Philornis systematics in southern South America.
Philornis larvae were collected from broods of Cacicus haemorrhous Linnaeus (Passeriformes: Icteridae) in the rainforest habitat surrounding Puerto Iguazú (25°42′S, 54°27′W). Twelve third-instar larvae were collected from 3 broods and incubated at room temperature. Emerging adults were morphologically identified by a Philornis specialist (MAQ) following a thorough analysis of wings (ciliated/unciliated, r-m and m-m veins pigmentation), eye separation, cheek hair coloration, number of setae in fore/hind tibia, among other characters included in the taxonomic review and description of the species involved (Couri, 1999; Couri et al., 2009).
Genomic DNA from individual flies was extracted using the AccuPrep® Genomic DNA Extraction Kit (Bioneer, USA) following the manufacturer's protocol. An ITS2 fragment was amplified using the following primers as previously described (Monje et al., 2013): ITS2-LEcEn-F and ITS2-LEcEn-R. PCR products were separated by electrophoresis in a 1.5% agarose gel stained with GelRed™ (Biotium, USA) and examined by UV transillumination. All PCR products were column purified and sequenced directly in both directions using amplifying primers. Sequences were aligned with those previously reported for Philornis and closely related fly species using the MUSCLE sequence alignment tool. The sequence alignment and the analyses mentioned below were carried out with the program Mega version 5.0 (Tamura et al., 2011). A phylogenetic analysis was performed with the Maximum-likelihood (ML) method. The best fitting substitution models were determined with the Akaike Information Criterion using the ML model test. Support for the topology was tested by bootstrapping over 1,000 replications, excluding gaps and missing data.
Out of the 9 adult specimens from Misiones that emerged from pupae in the laboratory, 6 were morphologically identified as P. seguyi, whereas the remaining 3 were identified as P. torquans. Simultaneous infestation of 2 morphologically distinct species of Philornis was previously reported by Nores (1995), who described co-infestation of Philornis pici and P. seguyi in broods of Anumbius annumbi Orbigny, Lafresnaye (Passeriformes: Furnariidae) and Pseudoseisura lophotes Reichenbach (Passeriformes: Furnariidae). We excluded from the analysis specimens with intermediate features, and only those that showed all the distinctive morphological characters for each species were processed for sequencing. Three specimens identified as P. seguyi and 1 identified as P. torquans were therefore included in the analysis.
The ITS2 sequences from the above 4 specimens were identical regardless of their morphological identification, whereas their divergence from other known Philornis’ ITS2 sequences, ‘Philornis sp. genotype Central Argentina’ (KC485555), ‘Philornis sp. genotype Magdalena’ (KC485557) and P. downsi (HQ834532) was of 11.7%, 9.2% and 27.9%, respectively. In the light of these results, hereafter all specimens from Misiones are referred to as ‘Philornis sp. genotype Misiones’.
The phylogenetic tree inferred from maximum likelihood analysis of the ITS2 sequences for Philornis spp. and closely related fly species showed the ‘Philornis sp. genotype Misiones’ nested in a clade together with ‘Philornis sp. genotype Central Argentina’ and ‘Philornis sp. genotype Magdalena’, all of which are considerably similar morphologically. The morphologically distinct P. downsi was, on the other hand, placed in a separate clade (Fig. 1B). The newly generated sequences were deposited on GenBank (GenBank accession numbers KJ187043–KJ187046). The sequence alignment and the ML tree can be retrieved from the Treebase website (www.treebase.org; ID number 19285).
Monje et al. (2013) identified a single genotype (here named ‘Philornis sp. genotype Central Argentina’) in samples from 5 out of 6 localities studied (Fig. 1A), whereas a second lineage named ‘Philornis sp. genotype Magdalena’ was found in specimens from the southernmost locality (Magdalena city, Fig. 1A). On the basis of morphological characteristics, specimens from Magdalena were identified as P. seguyi (Couri et al., 2005; Monje et al., 2013), though it is unlikely that they actually belong to this species given that Misiones is spatially and ecologically distant from Magdalena. Herein the ITS2 sequences of Philornis specimens identified as P. torquans and P. seguyi from Misiones province (P. seguyi type locality) presented differences from sequences of Argentine subcutaneous Philornis previously reported (Monje et al., 2013), increasing to 3 the number of documented genotypes of these morphologically undistinguishable flies.
The genetic differences observed between the ITS2 sequences of these 3 genotypes of Philornis are higher than the interspecific variations observed between many fly species (Song, Wang, & Liang, 2008; Thanwisai, Kuvangkadilok, & Baimai, 2006). It is unknown whether ‘Philornis sp. genotype Misiones’ is indeed P. seguyi. Unfortunately, since the holotype of P. seguyi is lost, further morphological examination for confirmation of this species’ validity is infeasible. Our results highlight the need to revise and reconstruct the systematics of the genus Philornis complementing both molecular and morphological techniques.
Cryptic Philornis species are being ignored while scientific literature continues to report wrongly identified species. In this regard, the simultaneous infestation by 2 species of Philornis reported by Nores (1995) might also be due to misidentification. Taking into consideration that currently accepted taxonomic reviews (Couri, 1999; Couri et al., 2009) could not identify the species for each of the 3 genotypes described so far, ‘Philornis sp. genotype Central Argentina’, ‘Philornis sp. genotype Magdalena’ and ‘Philornis sp. genotype Misiones’, and that an analysis of their ITS2 sequences showed a close genetic relationship, we believe that these species could actually represent a complex of species under the taxon P. torquans.
We propose that, until more extensive studies of Philornis species are carried out including molecular data coupled with detailed morphological analyses and cross-mating experiments, the subcutaneous Philornis resembling P. seguyi or P. torquans in southern South America should be treated as the ‘Philornis torquans complex’, as previously proposed for other taxa (e.g. Bertrand et al., 2014; Nava et al., 2014).
We would like to thank Dr. David Anderson for proofreading this work. All procedures conducted in this study comply with the current National and Provincial laws, and were approved by the Bioethics Committee of Universidad Nacional del Litoral. This work was funded by the Argentine Council for Research and Technology (Grant No. PIP 1122010010026).
Peer Review under the responsibility of Universidad Nacional Autónoma de México.