












To determine the diversity of species and subspecific taxa of diatoms of the genus Lyrella found in subtidal sediments of Laguna Guerrero Negro (LGN), BCS-BC (Mexico), samples from 4 seasons were analyzed. Twenty-one species and sub-specific taxa of Lyrella are here recorded within a floristic list of 232 identified epipelic diatom taxa. Seven Lyrella taxa are new records for the Mexican coasts and 12 for the northwestern Mexican region. Additionally, we are proposing 6 transfers of taxa into Lyrella that remained hitherto in the genus Navicula. Morphometric and meristic data, and an iconographic reference are provided. Environmental conditions in the lagoon favor the diversity and abundance of Lyrella species and sub-specific taxa. Thus, LGN is proposed as a diversity hotspot for this genus. Lack of floristic studies on benthic diatoms in much of the Mexican coasts precludes insight of their ecology and distribution. However, continuous floristic surveys should eventually allow precise definition of their biogeographic affinities.
Con el objetivo de determinar la diversidad de especies y taxones subespecíficos de Lyrella presentes en sedimentos del submareal en la laguna Guerrero Negro, BCS-BC (México), se analizaron muestras de 4 estaciones. Se registraron 21 especies y taxones subespecíficos de Lyrella como parte de una lista florística de 232 taxones epipélicos. Siete de estos taxones son nuevos registros para las costas mexicanas y 12 para la región noroeste de México. Además, se proponen 6 transferencias de taxones que hasta ahora eran consideradas dentro del género Navicula hacia el género Lyrella. Se proporcionan datos morfométricos y merísticos, así como una referencia iconográfica. Dado que las condiciones ambientales propias de la laguna favorecen la diversidad y abundancia de especies y taxones subespecíficos de Lyrella, laguna Guerrero Negro se propone como un hotspot de diversidad para este género. Aunque la escasez de estudios florísticos sobre diatomeas bentónicas en las costas mexicanas limita la perspectiva ecológica y biogeográfica que ameritan, se confía en que la continuidad de estudios florísticos reditúe bases para definir sus afinidades biogeográficas.
Studies on benthic diatoms (Bacillariophyceae) from the Baja California Peninsula date back 3 decades to the first list of benthic diatoms (Hernández-Becerril & Álvarez-Borrego, 1983) followed by a more complete study on epiphytic forms thriving on eelgrass Zostera marina Linneo (Siqueiros-Beltrones & Ibarra-Obando, 1985; Siqueiros-Beltrones, Ibarra-Obando, & Loya-Salinas, 1985). Further studies of benthic diatoms included other substrata, such as sediments from hypersaline environments (Brown, Margulis, Ibarra, & Siqueiros, 1985; López-Fuerte & Siqueiros-Beltrones, 2006; López-Fuerte, Siqueiros-Beltrones, & Navarro, 2010; Siqueiros-Beltrones, 1988, 1990; Siqueiros-Beltrones & Morzaria-Luna, 1999; Siqueiros-Beltrones & Sánchez-Castrejón, 1999; Siqueiros-Beltrones, Ibarra-Obando, & Poumián-Tapia, 1991), and highly productive ecosystems (Hernández-Almeida & Siqueiros-Beltrones, 2012; Siqueiros-Beltrones, 2006). Although these studies recorded over a 1,000 species for this region, the many distinct ecosystems along the latitudinal gradient and various types of substrata are yielding taxa not seen before, or showing that hitherto rare taxa may occur abundantly. In this study, we focused on the benthic diatoms from fine sediments covering the sub-tidal bottom of Laguna Guerrero Negro (LGN), located between BCS and BC, a coastal lagoon found half way on the western coast of the BC Peninsula, Mexico.
Initial observations of said sediments suggested that the diatom flora in this location would be highly diverse and that hitherto rare or uncommon taxa recorded in previous studies in the region would be better represented, such as Lyrella Karayeva (Bacillariophyceae: Lyrellales). This taxonomic group comprises solely marine species which are commonest in the epipelon of sandy sediments (Round, Crawford, & Mann, 1990). Species formerly classified as Navicula were first allocated in the genus Lyrella by Karayeva (1978) based on a single morphological trait, the conspicuous double lyrae. This taxonomic segregation was later justified in depth by Mann and Stickle (1993), showing that their former status as Navicula was highly artificial, and currently they represent separate orders.
Although only 53 taxa transferred by D.G. Mann were listed by Round et al. (1990), currently 82 Lyrella taxa have been uploaded in the algaebase webpage (Guiry & Guiry, 2015) thus supporting the early expectations by Mann and Stickle (1993) on the diversity of this taxon. Species of the genus Lyrella include naviculoid, solitary individuals mostly appearing in valval view. Valve face are flat or slightly undulate with relatively shallow mantles. Frustules are heavily silicified, broadly linear or lanceolate with bluntly rounded or rostrate poles. The raphe system is central and straight. Striae are coarse, uniseriate, and are interrupted by a lyre-shaped thickened area of silica, often depressed (Round et al., 1990). Notwithstanding, the similarity with certain naviculales such as Fallacia posed the risk of confusion in routine identifications. However, based on the above, the presence of a single plastid in Fallacia and 2–4 in Lyrella, that justifies their placement in different orders is very helpful to discriminate between them observing fresh material. Also, relying on experience with benthic diatom taxocoenoses, the strongly silicified frustule of Lyrella species projects the double lyrae, that are simulated traits (fallacious) in Fallacia species which are mostly small and inconspicuous; with certain exceptions most likely from fresh-water habitats, where Lyrella do not occur.
Species of Lyrella have been frequently observed in benthos samples from the Baja California Peninsula coasts where they appear conspicuously mainly because of their particularly esthetic forms. However, the variety and abundance of Lyrella species may be considered moderately high in certain environments of the region, such as mangrove systems like Magdalena Bay, BCS where 9 Lyrella taxa were recorded in a single locality (Siqueiros-Beltrones, López-Fuerte, Hernández-Almeida, & Argumedo-Hernández, 2011). Likewise, 10 Lyrella taxa were recorded as epiphytes on macroalgae in the southernmost region of the east coast of the Baja California Peninsula where mangrove forests are common (Siqueiros-Beltrones & Hernández-Almeida, 2006). Furthermore, up to 11 taxa were recorded in a study that comprised 3 mangrove ecosystems of the southern BC Peninsula, and 1 from Sinaloa in mainland Mexico that included tychoplanktonic forms (López-Fuerte et al., 2010). Also, the number of Lyrella species recorded in studies on phytoplankton may be as high as in the sediments. For example, Meave-del Castillo, Zamudio-Resendis, Aké-Castillo, Guerra-Martínez, and Barbosa-Ledezma (2003) compiled 11 Lyrella taxa records in the tychoplankton of the Mexican Pacific, and Moreno-Ruíz, Licea-Durán, and Santoyo (1996) accounted for 13 Lyrella taxa for the Gulf of California. Whilst by comparison Navarro and Hernández-Becerril (1997) compiled 13 taxa records from the highly diverse Caribbean Sea. These accounts set a reference as to the expected species variety in any given area for the genus Lyrella.
Earlier, Magdalena Bay was proposed as a “hot spot” for diatom species diversity, in which the high number of Lyrella (and Mastogloia) taxa marked the difference with other ecosystems in the region (Siqueiros-Beltrones et al., 2011). As stated above our first observations lead us to hypothesize that LGN would harbor a high number of Lyrella species and sub-species taxa, in which case this area would fit the denomination of hotspot used in earlier works.
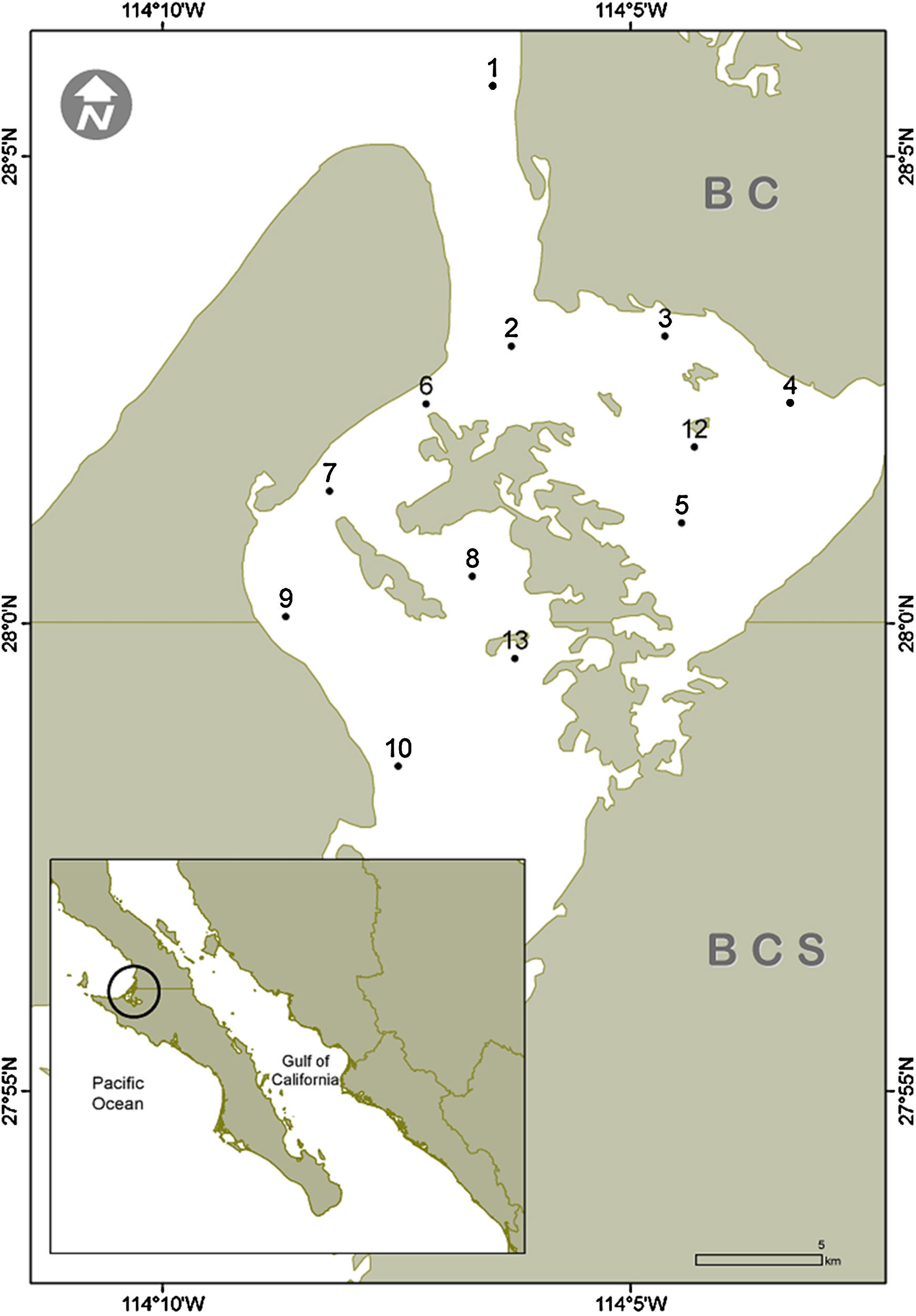
Materials and methodsLaguna Guerrero Negro is located between 27°35′ and 27°52′ N, 113°58′ 114°10′ W in the northern- and southernmost parts of BCS and BC, Mexico, respectively (Fig. 1); it is part of a complex along with Laguna Ojo de Liebre, comprised within the Reserva de la Biosfera del Vizcaíno (Arellano-Martínez, De La Cruz-Agüero, & Cota-Gómez, 1996). The lagoons have separate mouths connecting them with Sebastián Vizcaíno Bay. The climate in the region is arid with a low annual rainfall (mostly in winter) of >100mm (Salinas-Zavala, Llinas, & Rodríguez-Estrella, 1991). LGN is rectangular and extends approximately 2,100ha, with a length of 13km, and a maximum width of 8km, with a narrow channel that opens into Vizcaíno Bay (Contreras, 1985). It has a shallow bottom, mostly between 2 and 12m (Lluch-Cota, Castellanos-Vera, Llinas-Gutiérrez, & Ortega-Rubio, 1993), with a maximum depth of 26m; eel grass is amply distributed from 6m deep up to high tide level (Eberhard, 1966). Lagoon sediments are composed mainly of gray sand mixed with organic alluvial deposits. Normal salinity gradients, and hypersalinity gradients as well, have been reported (Lankford, 1977), although it has also been considered an isohaline lagoon; salinity values range from 35.5 to 37.5 (winter), and 34.7–35.6 (summer) (Phleger & Ewing, 1962).

Sediment samples were collected in November (2013), January, March, June, and July (2014) in 12 sites within LGN at depths between 3 and 15m (Fig. 1). Approximately 150g of sediments from the upper layer were sampled by scuba diving using a 250mL plastic jar.
At the laboratory, a subsample was placed in a beaker dissolved with 50mL of purified water. This was submerged in water contained in the ultrasound bucket and treated with ultrasound for 1min while agitating gently. Afterwards, sediments were removed by decanting while the remaining fraction was placed in a 100-mL test tube. After 2h, the overlaying water was decanted in-order-to separate a concentrate of diatom frustules in the bottom of the tube. A subsample from this concentrate was examined under the microscope for observing live diatoms. The rest of the sample was treated with alcohol and nitric acid at a ratio of 1:3:5, adjusting to the amount of organic matter in the sample (Siqueiros-Beltrones, 2002). After oxidation, the samples were rinsed repeatedly with purified water until reaching a pH ≥6. From each subsample 2 double permanent slides were mounted using the synthetic resin Pleurax (IR=1.7).
The diatom slides were examined under an optical microscope with phase contrast and planapochromatic optics. Species identification was done based on regional references: and basic literature: Hernández-Almeida and Siqueiros-Beltrones (2008, 2012), López-Fuerte et al. (2010), Moreno-Ruíz et al. (1996), Siqueiros-Beltrones (2002, 2006), Siqueiros-Beltrones and Hernández-Almeida (2006), Siqueiros-Beltrones, Argumedo-Hernández, Murillo-Jiménez, and Marmolejo-Rodríguez (2014), Hustedt (1959, 1966), Peragallo and Peragallo (1908), Round et al. (1990), Schmidt et al. (1959), Stidolph, Sterrenburg, Smith, and Kraberg (2012), and Witkowski, Lange-Bertalot, and Metzeltin (2000). All taxa were updated using AlgaeBase (Guiry & Guiry, 2015). An iconographic reference was built using micrographs taken with a CMOS Konus digital ocular lens at 1,000×. Measurements of specimens were done directly with a calibrated micrometric ocular lens, and images were manipulated so esthetically fit the plates according to size (interval when possible).
ResultsOut of a floristic list of 232 identified diatom taxa from subtidal sediments of LGN for the sampling period, 21 species and sub-species taxa of Lyrella were recorded; most were observed frequently although 6 are hitherto represented only by a single specimen. The Lyrella taxa represent less than 10% of the floristic list, however, its diversity is second only to Amphora (25), and clearly surpasses previous records, thus supporting our proposal of considering LGN as a hot-spot for Lyrella species, at least until further surveys in Mexican coasts are carried out.
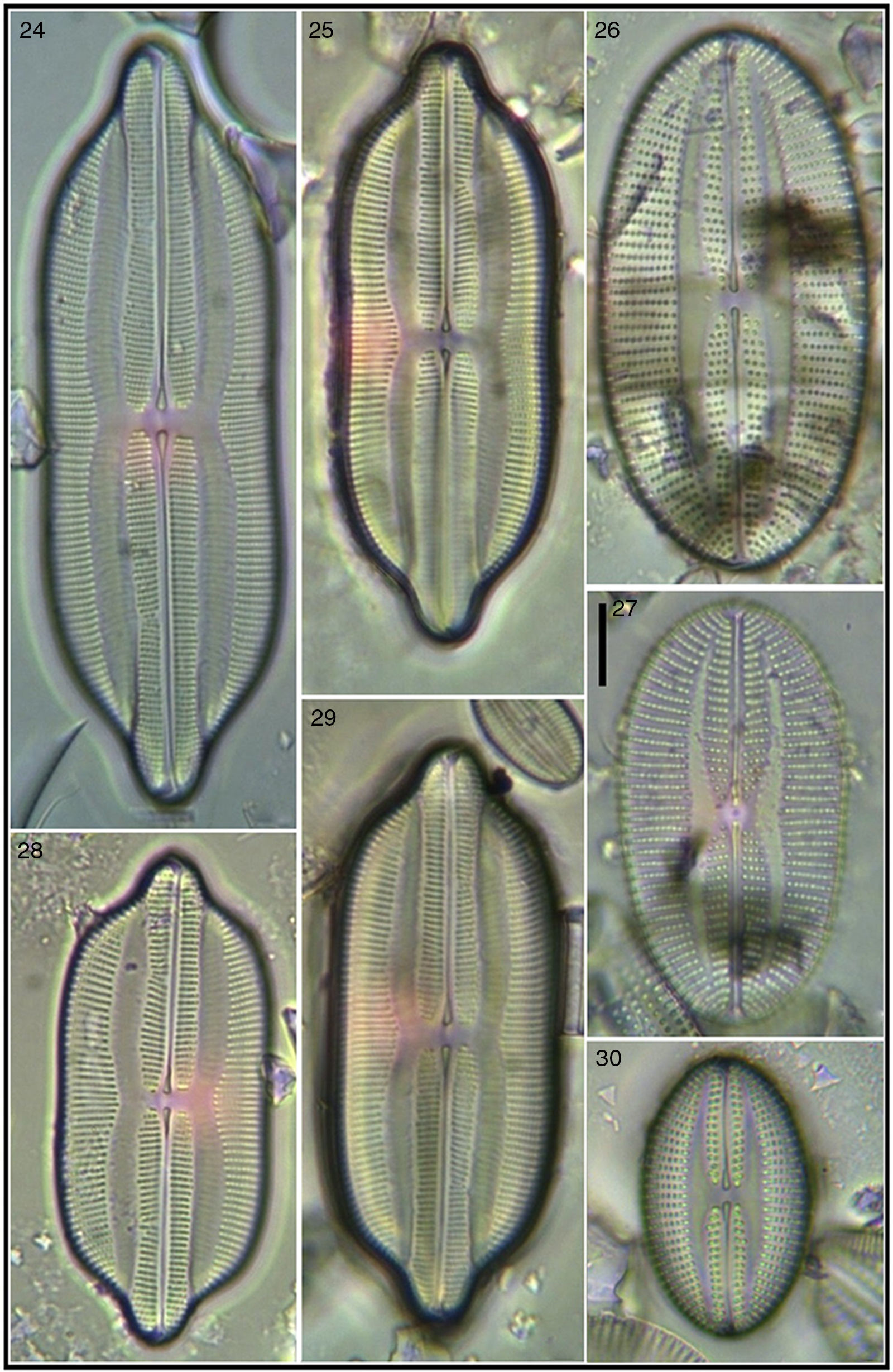
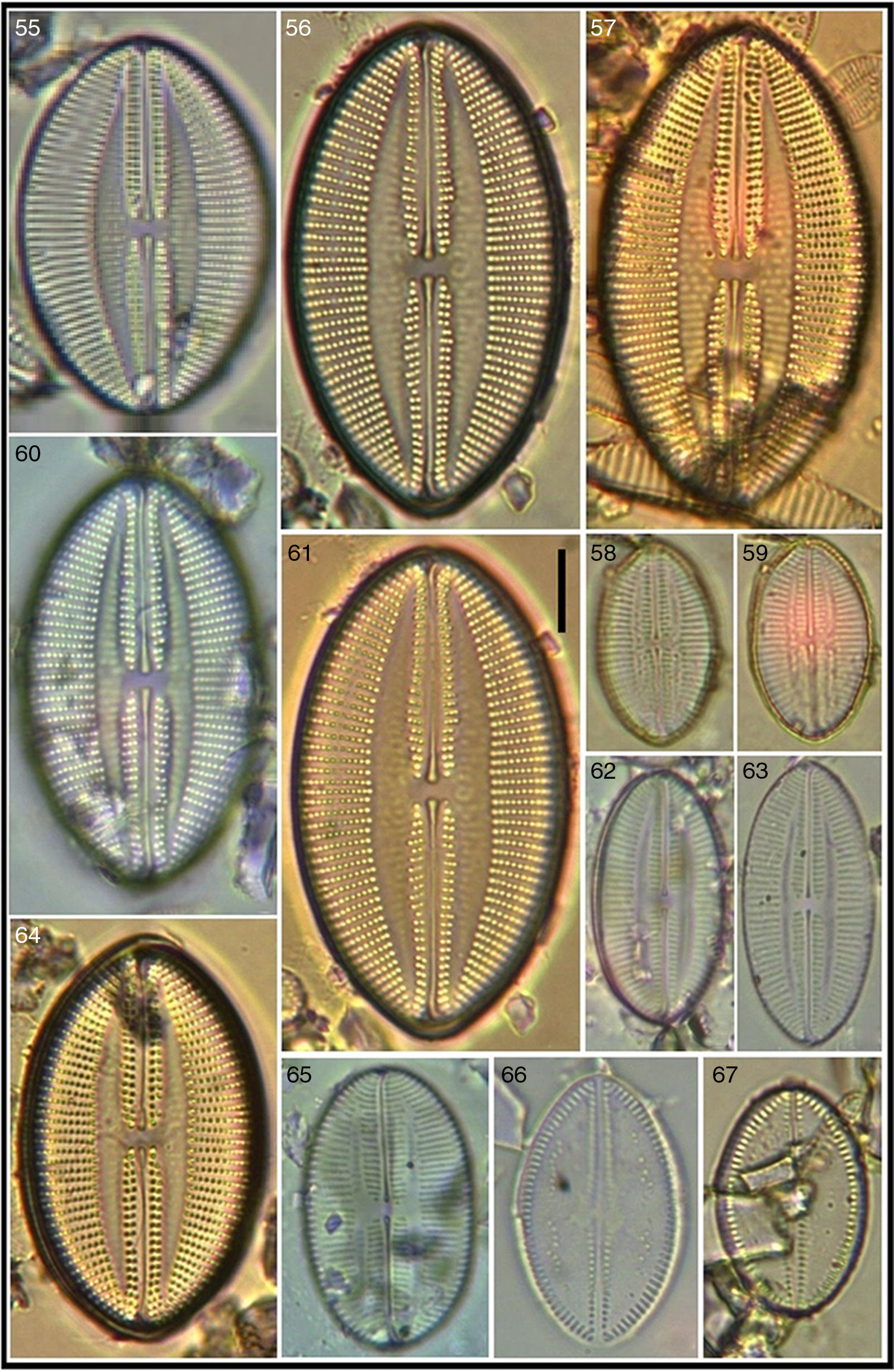
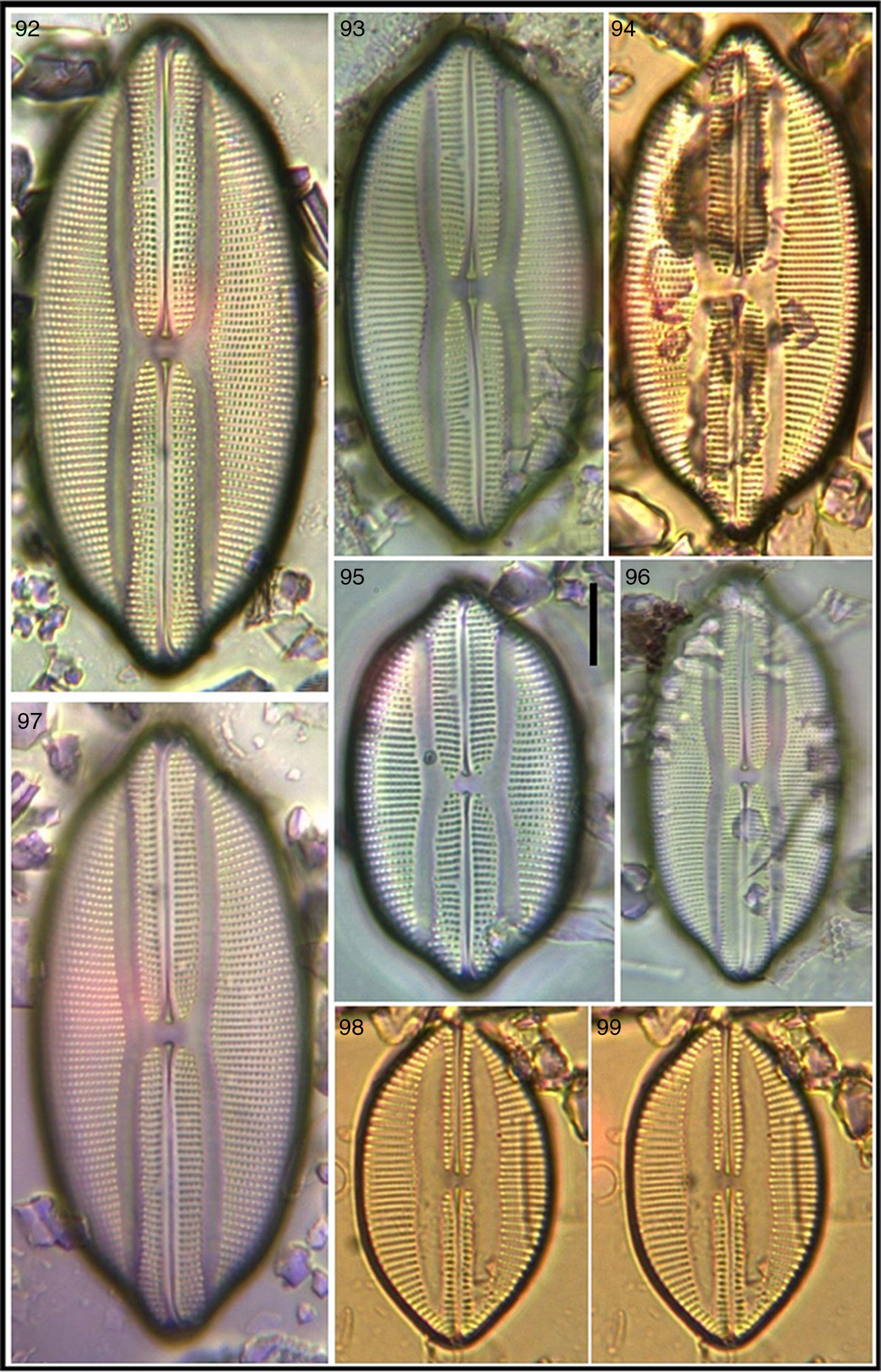
Seven of the Lyrella taxa recorded are new for the Mexican coasts, 12 for the NW Mexican region, and 1 for the western coast of the Baja Peninsula. Six taxa had not been transferred from their original genus Navicula and are here proposed as new combinations and transferred to the genus Lyrella; morphometric and meristic data are included for the identified Lyrella. An iconographic reference (Figs. 2–105) is also provided where it is shown that much fine sediment precluded better images of some specimens which were however included to show variation within the taxon.













In what follows, descriptions of the Lyrella species and sub-species taxa identified in subtidal sediments of Laguna Guerrero Negro are provided. Twelve are new records for the region (*), 7 for the Mexican coasts (●), and 2 for the W coast of the Baja California Peninsula (**). Six new nomenclatural combinations (comb. nov.) are proposed. Morphometric and meristic data are provided for all taxa.
Lyrella abrupta (Gregory) D.G. Mann
Dimensions: length 21–35μm, width 12–19μm, 14 striae in 10μm (n=4).
Reference: López-Fuerte et al. (2010), Witkowski et al. (2000).
Basionym: Navicula lyra var. abrupta Gregory.
Synonym (homotypic): Navicula lyra var. abrupta Gregory.
Previous records: Navachiste, Sinaloa (López-Fuerte et al., 2010); La Ventana Bay, BCS (Siqueiros-Beltrones et al., 2014).
Lyrella approximatoides (Hustedt) D.G. Mann in Round, Crawford et Mann.
Dimensions: length 29–60μm, width 19–25, 10 striae in 10μm (n=12).
Reference: Foged (1984) as Navicula approximatoides; Round et al. (1990).
Basionym: Navicula approximatoides Hustedt.
Synonym: none.
Previous records: Magdalena Bay, BCS (López-Fuerte et al., 2010); La Paz Bay (Siqueiros-Beltrones & Argumedo-Hernández, 2012).
Lyrella atlantica (A. Schmidt) D.G. Mann ●*
Dimensions: length 34–76, width 18–27, 8–10 striae in 10μm.
Reference: Hendey (1964), Witkowski et al. (2000).
Basionym: Navicula lyra var. atlantica A. Schmidt.
Synonym (heterotypic): Navicula lyra var. atlantica A. Schmidt.
Previous records: none.
Lyrella clavata var. caribaea (Cleve) Siqueiros-Beltrones comb. nov.
Dimensions: length 34–88μm, width 20–30, 11–13 striae in 10μm (n=15).
References: Schmidt et al. (1959) as Navicula cariboea Cleve; Peragallo and Peragallo (1908) as Navicula clavata var. cariboea Cleve; López-Fuerte et al. (2010) as Navicula caribaea Cleve.
Basionym: Navicula caribaea Cleve in Schmidt et al. (1959).
Synonym: Lyrella clavata var. indica (Greville) J. L. Moreno-Ruíz.
Previous records: La Bocana, BCS (Siqueiros-Beltrones & Argumedo-Hernández, 2015).
Lyrella clavata var. elongata (H. Peragallo) Díaz-Ramos *●
Dimensions: length 47–84μm, width 26–33μm, 13 striae in 10μm (n=7).
References: Peragallo and Peragallo (1908) as Navicula clavata var. elongata; Díaz-Ramos (2000).
Basionym: Navicula clavata var. elongata H. Peragallo.
Synonym: Navicula clavata f. elongata H. Peragallo in Hustedt (1966).
Previous records: none.
Lyrella clavata var. indica (Greville) J. L. Moreno-Ruíz
Dimensions: length 60μm, width 32μm, 10 striae in 10μm (n=1).
Reference: Hustedt (1966), Moreno-Ruíz et al. (1996) (image of L. caribaea).
Basionym: Navicula indica Greville.
Synonym: Navicula clavata f. indica (Greville) Cleve.
Previous records: Gulf of California (Moreno-Ruíz et al., 1996).
Lyrella excavata (Hustedt) D.G. Mann *
Dimensions: length 32–44μm, width 18–22μm, 12–13 striae in 10μm (n=5).
Reference: López-Fuerte et al. (2010), Round et al. (1990), Schmidt et al. (1959) as Navicula spectabilis f. emarginata; Moreno-Ruíz et al. (1996) as Navicula spectabilis var. excavata (Grevillei) Cleve.
Basionym: Navicula excavata Greville.
Synonym (homotypic): Navicula excavata Greville.
Previous records: Magdalena Bay, BCS (López-Fuerte et al., 2010); Gulf of California (Moreno-Ruíz et al., 1996).
Lyrella exsul (A. Schmidt) D.G. Mann
Slender and with lyra sterna narrower than the type form in Hustedt's description.
Dimensions: length 30–75μm, width 18–24μm, 11–12 striae in 10μm (n=11).
Reference: Hustedt (1966), Peragallo and Peragallo (1908) as Navicula clavata var. exsul (A. S.) Cleve; Round et al. (1990).
Basionym: Navicula exsul A. Schmidt.
Synonyms (homotypic): Navicula exsul A. Schmidt, Navicula clavata var. exsul (A. Schmidt) Cleve.
Previous records: La Paz Bay (Siqueiros-Beltrones & Sánchez-Castrejón, 1999); Magdalena Bay, BCS (López-Fuerte et al., 2010).
Lyrella fogedii Witkowski, Lange-Bertalot et Metzeltin*
Dimensions: length 29μm, width 18μm, 11 striae in 10μm (n=1).
Reference: Witkowski et al. (2000).
Basionym: Lyrella fogediiWitkowski, Lange-Bertalot, & Metzeltin, 2000.
Synonym: Navicula aestimata Hustedt sensu Foged (1975).
Previous records: La Paz Bay (Siqueiros-Beltrones & Hernández-Almeida, 2006).
Lyrella fundata (Hustedt) Siqueiros-Beltrones comb. nov. *●
Dimensions: length 44–55μm, width 23–28μm, 9–10 striae in 10μm (n=3).
Reference: Hustedt (1966), where it is placed as Navicula fundata, in the vicinity of Navicula (Lyrella) abrupta and Navicula (Lyrella) abruptoides.
Basionym: Navicula fundata Hustedt.
Synonym: Navicula fundata Hustedt.
Previous records: none.
Lyrella granulata (Grunow) Nevrova, Witkowski, Kulikovskiy et Lange-Bertalot *●
Dimensions: length 28–67μm, width 18–32, 9–12 striae in 10μm (n=13).
References: Hustedt (1966), Nevrova, Witkowski, Kulikovskiy, Lange-Bertalot, and Kociolek (2013).
Basionym: Navicula hennedyi var. granulata Grunow.
Synonym (homotypic): Navicula hennedyi var. granulata Grunow.
Previous records: none.
Lyrella hennedyi var. crassa (Peragallo) Siqueiros-Beltrones comb. nov. *●
Striae bordering the raphe somewhat elevated.
Dimensions: length 72μm, width 34μm, 12 striae in 10μm (n=1).
Reference: Peragallo and Peragallo (1908).
Basionym: Navicula hennedyi var. crassa Peragallo.
Synonym: Navicula hennedyi var. crassa Peragallo in Peragallo and Peragallo (1908).
Previous records: none.
Lyrella hennedyi var. furcata (Peragallo) Siqueiros-Beltrones comb. nov. *●
Dimensions: length 30–48μm, width 18–26μm, 11–12 striae in 10μm (n=3).
Reference: Desikachary (1989), Lam. 374, fig. 10; Hustedt (1966).
Basionym: Navicula hennedyi var. furcata Peragallo.
Synonym: Navicula hennedyi var. furcata Peragallo in Peragallo and Peragallo (1908).
Previous records: none.
Lyrella impercepta (Hustedt) J. L. Moreno-Ruíz
Dimensions: length 20–33μm, width 15–20μm, 9–10 striae in 10μm (n=8).
Reference: Hustedt (1966) as Navicula impercepta; Moreno-Ruíz et al. (1996).
Basionym: Navicula impercepta Hustedt.
Synonym (homotypic): Navicula impercepta Hustedt.
Previous records: Gulf of California (Moreno-Ruíz et al., 1996); Magdalena Bay, BCS (López-Fuerte et al., 2010; Martínez-López, Siqueiros-Beltrones, & Silverberg, 2004).
Lyrella implana (Hustedt) J. L. Moreno-Ruíz**
Dimensions: length 40μm, width 25μm, 14 striae in 10μm (n=1).
Reference: Hustedt (1966) as Navicula implana; Moreno-Ruíz et al. (1996).
Basionym: Navicula implana Hustedt.
Synonym (homotypic): Navicula implana Hustedt.
Previous records: Gulf of California (Moreno-Ruíz et al., 1996).
Lyrella irrorata (Grevillei) D.G. Mann
Dimensions: length 55–110μm, width 27–34μm, 6 striae in 10μm (n=6).
Reference: Hustedt (1966), Round et al. (1990).
Basionym: Navicula irrorata Greville.
Synonym (homotypic): Navicula irrorata Greville.
Previous records: La Paz Bay (Siqueiros-Beltrones & Sánchez-Castrejón, 1999); Navachiste, Sinaloa (López-Fuerte et al., 2010).
Lyrella lyra (Ehrenberg) Karayeva
Dimensions: length 42–80μm, width 24–32μm, 11–12 striae in 10μm (n>20).
Reference: Round et al. (1990), Moreno-Ruíz et al. (1996), Witkowski et al. (2000).
Basionym: Navicula lyra Ehrenberg.
Synonyms: (homotypic): Navicula lyra Ehrenberg, Schizonema lyra (Ehrenberg) Kuntze, Clevia lyra (Ehrenberg) Mereschkowsky.
Previous records: Gulf of California (Moreno-Ruíz et al., 1996); Mexican Caribbean (López-Fuerte, Siqueiros-Beltrones, & Hernández-Almeida, 2013).
Lyrella lyra var. constricta (Peragallo) Siqueiros-Beltrones comb. nov. *●
Figures 13, 16, 24, 25, 28, 29
Dimensions: length 45–88μm, width 21–27μm, 11 striae in 10μm.
Reference: Hustedt (1966).
Basionym: Navicula lyra var. constricta Peragallo.
Synonym: Navicula lyra var. constricta Heiden.
Previous records: none.
Lyrella lyra var. subtypica (A. Schmidt) Siqueiros-Beltrones comb. nov. *
Earlier referred to as such by Moreno-Ruíz et al. (1996), Sánchez-Rueda (2002), and Martínez-López et al. (2004) as L. lyra f. subtypica, albeit a formal taxonomic transference was not proposed.
Dimensions: length 70–103μm, width 30–37μm, 12 striae in 10μm (n=9).
Reference: Hustedt (1966), Peragallo and Peragallo (1908).
Basionym: Navicula lyra var. subtypica A. Schmidt. http://www.algaebase.org/search/species/detail/?species_id=q7e8fb0c05459c57c
Synonym: Navicula lyra var. subtypica A. Schmidt in Hustedt (1966).
Previous records: Gulf of California (Moreno-Ruíz et al., 1996); Gulf of México (Sánchez-Rueda, 2002); off Magdalena Bay (Martínez-López et al., 2004).
Lyrella spectabilis (Gregory) D.G. Mann *
Dimensions: length 62μm, width 40μm, 10 striae in 10μm (n=1).
Reference: Hustedt (1966), Stidolph et al. (2012).
Basionym: Navicula spectabilis Gregory. http://www.algaebase.org/search/species/detail/?species_id=q7e8fb0c05459c57c
Synonym (homotypic): Navicula spectabilis Gregory.
Previous records: Campeche Bay (Stidolph et al., 2012).
Lyrella spectabilis (Gregory) D.G. Mann var. 1*
We refer this as a possible variety due to our specimen is clearly smaller and with a less conspicuous appearance than the former which is more robust.
Dimensions: length 33μm, width 20μm, 12–13 striae in 10μm (n=1).
Reference: Desikachary (1989), Lam. 648, fig. 4.
Basionym: Navicula spectabilis Gregory. http://www.algaebase.org/search/species/detail/?species_id=q7e8fb0c05459c57c
Synonym (homotypic): Navicula spectabilis Gregory.
Previous records: none.
DiscussionIn none of the few surveyed areas of the Mexican NW region, nor in other coasts of Mexico for that matter, have so many Lyrella species been recorded hitherto. Several of these species occur frequently in the Gulf of California, either as tychoplankton (Moreno-Ruíz et al., 1996), or have been collected directly from the benthos (Hernández-Almeida & Siqueiros-Beltrones, 2012; López-Fuerte & Siqueiros-Beltrones, 2006; López-Fuerte et al., 2010; Siqueiros-Beltrones, 2006; Siqueiros-Beltrones & Morzaria-Luna, 1999; Siqueiros-Beltrones & Sánchez-Castrejón, 1999; Siqueiros-Beltrones et al., 2011). On the other hand, taxa such as Lyrella approximata (Greville) D.G. Mann and Lyrella circumsecta (Grunow ex A. Schmidt) D.G. Mann (Siqueiros-Beltrones & Hernández-Almeida, 2006), or Lyrella abruptoides (Hustedt) D.G. Mann (Siqueiros-Beltrones, 2002) were not found in our samples, although all have been recorded previously further south just within the Gulf of California, with adjoining mangrove forests. Laguna Guerrero Negro is located northward beyond the geographic distribution of mangroves along the western coast of the Baja California Peninsula (González-Zamorano, Nava-Sánchez, León-de la Luz, & Díaz-Castro, 2011).
The variety of Lyrella species and sub-specific taxa, and their frequencies in the samples indicate that environmental conditions in LGN promote the high diversity of the genus. Which, along with the 12 new Lyrella records for the Mexican NW region back up the proposal that this area be considered a hot-spot for Lyrella. The fact that 6 taxa were represented by a single specimen shows the need for further observations, including other seasons. Current analysis of the whole taxocoenosis as well as continuing sampling during other months (only 4 were surveyed) may yield other Lyrella species, or further information on the temporal distribution of those recorded, providing further evidences for comprising this and the rest of the taxa under the hot-spot concept.
It has to be implied that the geographic distributions of the Lyrella taxa recorded here are also extended. In this subtropical zone, the possible influence of tropical waters in the area has been underlined by Hernández-Rivas, Jiménez-Rosenberg, Funes-Rodríguez, and Saldierna-Martínez (2000), albeit the California Current and regional upwelling events stand out in the region. The low temperature water they provide correlate with certain characteristic biota recorded by Eberhard (1966), which is found also in San Quintín bay and Punta Banda estuary further north in Baja California. This may be defining a transitional zone as proposed earlier by Dawson (1960) and observed later in the structure of phytoplankton assemblages (Gárate-Lizárraga & Siqueiros-Beltrones, 1998) for the Magdalena Bay lagoon complex. All this may also be used to explain the presence of Lyrella species from tropical, subtropical and temperate regions. Furthermore, López-Fuerte, Siqueiros-Beltrones, and Yabur (2015) observed a particular benthic diatom flora in Isla Guadalupe, located further north off the west coast of Baja California for which a biogeographical affinity could not be established, with many tropical species (Mastogloia spp.) including recent records from the Caribbean Sea. Likewise, the transitional nature of the oceanographic region would be expected to influence the temporal occurrence of certain Lyrella species. However, particular environmental characteristics will determine whether these remain as permanent residents of LGN or not.
From another angle, the transfer of the six Navicula taxa to Lyrella in this paper is an example of the general inertia caused by the establishment of new genera. At first, taxa are rapidly included but process slows down gradually until new surveys record the remaining taxa, as is presented here as a side effect of a floristic effort. As it turns out, identification of diatom species and construction of floristic lists is a still much needed activity that is improving on the basis of comprehensive taxonomic works that incorporate the latest advances in diatom systematics. In this case our contribution is possible because of the combined work of Guiry and Guiry (2015), Hustedt (1959), Karayeva (1978), Mann in Round et al. (1990).
In spite of the above, identification of diatom species and sub-species taxa poses much difficulty and always will. Although Lyrella species are conspicuous some difficulty exists especially for the untrained eye when certain Fallacia taxa appear in the samples, and certain experience with benthic diatom taxocoenoses is required to readily discriminate between these genera relying on the taxonomic traits exposed earlier. Moreover, trying to discern between certain Lyrella species or sub-species taxa may imply an important degree of uncertainty. An example is the raphe valve of Cocconeis californica var. kerguelensis Heiden, initially mistaken for Lyrella and then Craspedopleura (Figs. 66 and 67). This happens particularly in the traditional practice of identifying diatoms from samples that yield over 150 species and resorting to several taxonomic references. An example is the nine Lyrella from Magdalena Bay (Siqueiros-Beltrones et al., 2011) which were reduced from 15 that were recorded earlier (López-Fuerte & Siqueiros-Beltrones, 2006). Also, because in this study we focused solely on diatoms of the Lyrella group it was expected that a more efficient discrimination of the taxa be done, thus resulting in the higher number of species and sub-specific taxa here recorded. In this case, our floristic effort yielded 12 new records for the region although only samples for 4 months were surveyed. It is thus evident that the lack of floristic studies on benthic diatoms in much of the Mexican coasts precludes insight of their ecology and distribution. However, continuous floristic surveys should allow precise definition of biogeographic affinities of benthic diatom flora in the coasts of Mexico.
This study was financed by project SIP-20150537, Instituto Politécnico Nacional. We appreciate advice given by Sylvia Sala, Nelson Navarro, Eberto Novelo and Oscar Romero on formal taxonomy of diatoms. DASB is COFAA and EDI fellow of the IPN. FOLF currently holds a postdoctoral research grant from Conacyt. The reviews by 2 anonymous referees helped to improve this manuscript.
Peer Review under the responsibility of Universidad Nacional Autónoma de México.