



Se identificaron 12 especies de cangrejos ermitaños del material recolectado en la bahía Chamela, Jalisco. La familia Diogenidae fue la mejor representada con 7 especies. Nueve especies representan registros nuevos para la bahía y 2 pertenecientes al género Paguristes se descubrió que no han sido descritas. Se encontró asociación de los cangrejos ermitaños con conchas de 29 especies de gasterópodos que utilizaban como habitáculo; las conchas más comunes fueron Cantharus sanguinolentus (Duclos, 1833) y Mancinella speciosa (Valenciennes, 1832).
Twelve species of hermit crabs were identified from material recollected in Chamela Bay, Jalisco. The family Diogenidae was the best represented with 7 species. Nine species represent new records for Chamela Bay and 2 species of the genus Paguristes were found to be undescribed. We found an association of hermit crabs with shells used as home in 29 gastropods; the commonest shells used were Cantharus sanguinolentus (Duclos, 1833) and Mancinella speciosa (Valenciennes, 1832).
Entre los principales propósitos de las áreas naturales protegidas se incluyen la conservación y manejo sustentable de las poblaciones de especies susceptibles de explotación comercial; así como la preservación de los ambientes y la conservación de su biodiversidad (Palumbi, 2001). Bajo ese mismo esquema, el conocer la diversidad biológica nos permite establecer estrategias eficientes para la preservación de los recursos. La mayoría de los estudios biológicos se ha enfocado hacia el conocimiento de la biología y dinámica de las especies de interés comercial. Sin embargo, el manejo adecuado de los recursos naturales y la protección de los ecosistemas requiere necesariamente del conocimiento integral de su biodiversidad.
El orden Decapoda es uno de los grupos más diversos de crustáceos, con más de 17 600 especies, y es uno de los mejor conocidos debido a que muchas de sus especies presentan importancia económica (De Grave et al., 2009). Dentro de este orden, los anomuros (infraorden Anomura) presentan el mayor grado de disparidad morfológica, con formas corporales que van desde los cangrejos ermitaños simétricos y asimétricos a las langostillas y cangrejos rey (McLaughlin, 1980; Tsang et al., 2011). Los cangrejos ermitaños, representados por poco más de 1 100 especies a nivel mundial (McLaughlin et al., 2010) son considerados de gran importancia ecológica por su participación en las redes tróficas y en la recirculación de nutrientes del fondo marino, por sus interacciones con otros organismos y por su papel en la generación de sustratos estables mediante el uso de materiales calcáreos como refugio, tales como conchas vacías de moluscos o tubos de poliquetos (Williams y McDermott, 2004; Campos et al., 2011).
La fauna de cangrejos ermitaños en el Pacífico oriental es relativamente bien conocida aunque aún quedan muchas áreas costeras por estudiar. Estudios recientes han permitido aumentar considerablemente el conocimiento de la diversidad y distribución de los cangrejos ermitaños de la superfamilia Paguroidea en la región (McLaughlin, 1981, 1982; Harvey y McLaughlin, 1991; McLaughlin y Haig, 1993; Lemaitre y McLaughlin, 1996; Hendrickx y Esparza-Haro, 1997; Lemaitre, 1999; Hendrickx y Harvey, 1999; Lemaitre y Cruz-Castaño, 2004; Hendrickx et al., 2008; Ayón-Parente, 2009; Ayón-Parente y Hendrickx, 2005, 2006, 2007, 2009, 2010).
De acuerdo con Hendrickx (1993), Hendrickx y Harvey (1999) y Ayón-Parente y Hendrickx (2010), en el Pacífico mexicano se reconocen 90 especies de 4 familias de Paguroidea, mientras que en la costa de Jalisco se han registrado 12 especies de las familias Coenobitidae, Diogenidae y Paguridae (Ríos-Jara et al., 2013). En bahía Chamela sólo se tiene el registro de 4 especies: Aniculus elegans Stimpson, 1859; Calcinus californiensis Bouvier, 1898; C. explorator Boone, 1930 y Trizopagurus magnificus (Bouvier, 1898), todas ellas asociadas a formaciones coralinas (López-Uriarte y Ríos-Jara, 2004; López-Uriarte et al., 2009).
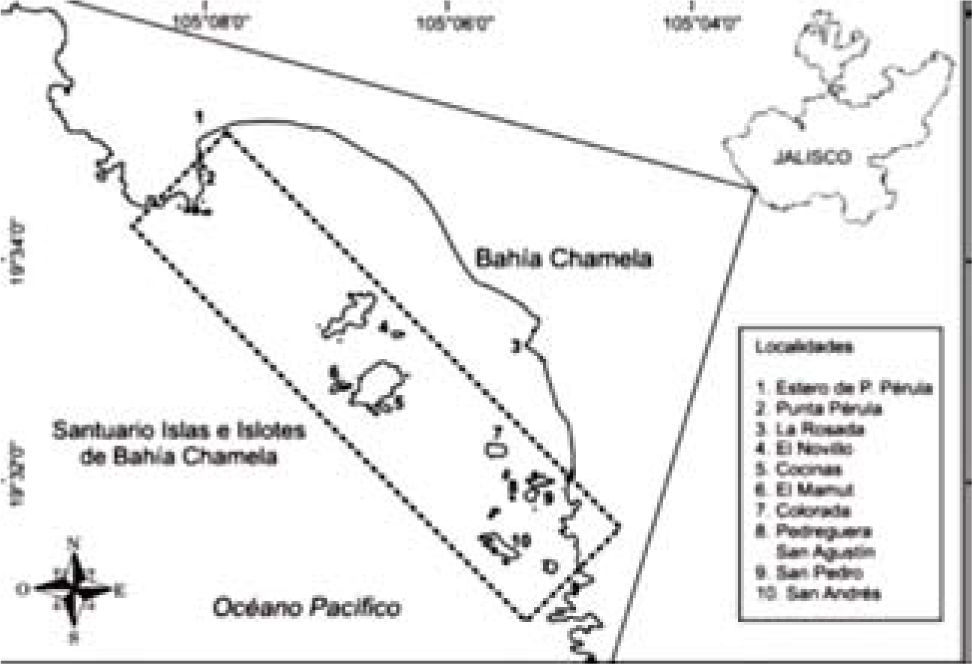
La bahía Chamela se encuentra localizada en la costa central de Jalisco (19°32' N, 105°06' O), en el Pacífico central mexicano. Tiene una extensión aproximada de 7.5km con dirección noroeste-suroeste y presenta una costa de tipo abrasivo con playas largas alternando con puntas rocosas (Cotler et al., 2002) (Fig. 1). Dentro de la bahía se encuentra una línea de islas e islotes en su mayoría conformados por acantilados rocosos, sólo algunas presentan playas arenosas o de canto rodado. El fondo marino de la bahía está constituido, principalmente, de sustratos rocosos, coralinos y arenosos los cuales ofrecen diversas opciones de hábitat y refugio para las especies (López-Uriarte et al., 2009). Dada la gran riqueza de flora y fauna registradas en las islas de la bahía, se emitió un decreto en el Diario Oficial de la Federación (DOF) en 2002 en el que se les considera como área natural protegida con la categoría de Santuario (DOF, 2002). Este ecosistema es aledaño a la Reserva de la Biosfera Chamela-Cuixmala, de la cual se tiene un extenso inventario de la flora y fauna (Conanp, 2008). Sin embargo, el conocimiento sobre la biodiversidad del Santuario Islas de la Bahía Chamela es todavía limitado (Román et al., 1991; León-Regagnon et al., 1997; López-Uriarte y Ríos-Jara, 2004; Nuño-Hermosillo et al., 2006; López-Uriarte et al., 2009; Galván-Villa, 2010; Ríos-Jara et al., 2013). Con el propósito de contribuir al conocimiento de la diversidad de invertebrados de la bahía Chamela, en el presente trabajo se registra la presencia y distribución de los cangrejos ermitaños recolectados en los distintos ambientes, además, el tipo de habitáculo (conchas) ocupado por éstos.
Materiales y métodos
Entre julio de 2007 y noviembre de 2011 se realizaron 10 salidas de muestreo en 10 localidades a lo largo de toda la bahía (Fig. 1), abarcando los ambientes supralitoral, intermareal y submareal somero. Los organismos se recolectaron manualmente mediante búsqueda directa en playas y recorridos con buceo autónomo en el submareal somero (≤10m de profundidad). Los sitios de muestreo fueron ubicados con el geoposicionador satelital Garmin Etrex. El material recolectado fue fijado y preservado en etanol al 70%. Todos los ejemplares fueron medidos y sexados, con excepción de los que no pudieron extraerse de las conchas. La talla de los organismos se consideró a partir del largo del escudo (LE), desde la punta del diente rostral hasta la parte media posterior del escudo. Los organismos no medidos ni sexados se indican con NM y NS, respectivamente. El arreglo taxonómico de las familias se realizó de acuerdo con lo propuesto por McLaughlin et al. (2010).
Las conchas de gasterópodos utilizadas por los cangrejos ermitaños fueron separadas e identificadas de acuerdo con Keen (1971) y su identidad taxonómica actualizada con la revisión de Skoglund (2002). El material fue depositado en la colección de referencia del Laboratorio de Ecosistemas Marinos y Acuicultura (LEMA-CR) del Centro Universitario de Ciencias Biológicas y Agropecuarias de la Universidad de Guadalajara (CUCBA-UDG) en Zapopan, Jalisco y en la Colección Regional de Invertebrados (EMU) en el Laboratorio de Invertebrados Bentónicos, ICMYL, UNAM, Mazatlán, México.
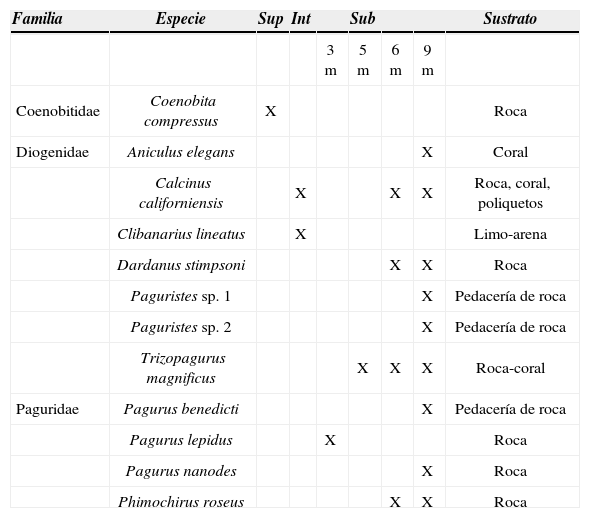
ResultadosSe recolectaron 122 ejemplares de cangrejos ermitaños pertenecientes a las familias Coenobitidae (1 especie), Diogenidae (7 especies) y Paguridae (4 especies). Los organismos fueron recolectados en sustratos duros (roca y coral) y limo-arenoso localizados desde el supralitoral hasta 10 m de profundidad (Cuadro 1). Sólo la especie Clibanarius lineatus fue recolectada en el estero de Punta Pérula en sustrato lodoso. Del total de organismos revisados, 107 ocupaban una concha (Cuadro 2) y el resto no contaba con habitáculo.
Distribución vertical de los cangrejos ermitaños. Los géneros y las especies se presentan alfabéticamente. Sup= supralitoral; Int= Intermareal; Sub= Submareal
| Familia | Especie | Sup | Int | Sub | Sustrato | |||
|---|---|---|---|---|---|---|---|---|
| 3m | 5m | 6m | 9m | |||||
| Coenobitidae | Coenobita compressus | X | Roca | |||||
| Diogenidae | Aniculus elegans | X | Coral | |||||
| Calcinus californiensis | X | X | X | Roca, coral, poliquetos | ||||
| Clibanarius lineatus | X | Limo-arena | ||||||
| Dardanus stimpsoni | X | X | Roca | |||||
| Paguristes sp. 1 | X | Pedacería de roca | ||||||
| Paguristes sp. 2 | X | Pedacería de roca | ||||||
| Trizopagurus magnificus | X | X | X | Roca-coral | ||||
| Paguridae | Pagurus benedicti | X | Pedacería de roca | |||||
| Pagurus lepidus | X | Roca | ||||||
| Pagurus nanodes | X | Roca | ||||||
| Phimochirus roseus | X | X | Roca |
Conchas utilizadas por las especies de cangrejos ermitaños recolectados en bahía Chamela, Jalisco, México
| Especies | Coenobita compressus | Aniculus elegans | Calcinus californiensis | Clibanarius lineatus | Dardanus stimpsoni | Paguristes sp. 1 | Paguristes sp. 2 | Trizopagurus magnificus | Pagurus benedicti | Pagurus lepidus | Pagurus nanodes | Phimochirus roseus | TOTAL |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nerita scabricosta | 7 | 1 | 8 | ||||||||||
| Cantharus sanguinolentus | 7 | 3 | 10 | ||||||||||
| Leucozonia cerata | 1 | 1 | 1 | 2 | 3 | 8 | |||||||
| Cerithium maculosum | 1 | 1 | 4 | 1 | 7 | ||||||||
| Mancinella speciosa | 2 | 6 | 4 | 1 | 7 | ||||||||
| Mancinella triangularis | 1 | 2 | 3 | ||||||||||
| Nassarius corpulentus | 1 | 1 | |||||||||||
| Cantharus elegans | 1 | 1 | 1 | 1 | 4 | ||||||||
| Collumbela fuscata | 2 | 2 | |||||||||||
| Nodilittorina aspera | 1 | 1 | |||||||||||
| Opeatostoma pseudodon | 1 | 4 | 5 | ||||||||||
| Rhinoclavis gemmata | 5 | 1 | 6 | ||||||||||
| Conus purpurascens | 1 | 1 | |||||||||||
| Tegula sp. | 4 | 1 | 5 | ||||||||||
| Cerithidea californica | 3 | 3 | |||||||||||
| Cerithidea cf. pulchra | 7 | 7 | |||||||||||
| Stramonita biserialis | 4 | 4 | 8 | ||||||||||
| Vasum caestus | 1 | 1 | |||||||||||
| Neorapana muricata | 5 | 5 | |||||||||||
| Cypreacassis coarctata | 5 | 1 | |||||||||||
| Cymatium lignarium | 1 | 1 | |||||||||||
| Bursa sp. | 1 | 1 | |||||||||||
| Homalocantha cf. oxyacantha | 1 | 1 | |||||||||||
| Favartia lappa | 2 | 2 | |||||||||||
| Anachis scalarina | 1 | 1 | |||||||||||
| Pilsbryspira garciacubasi | 1 | 1 | |||||||||||
| Muricopsis zeteki | 2 | 2 | |||||||||||
| Bursa corrugata | 2 | 2 | |||||||||||
| Conus brunneus | 1 | 1 |
Familia Coenobitidae Dana, 1851
Coenobita compressus H. Milne Edwards, 1837 Material examinado: isla San Andrés (19°31'21” N, 105°06'0), 03 Octubre 2008, intermareal rocoso, 3 machos (LE 1.7-2.6mm), 3 hembras (LE 1.9-3.7mm), LEMA-CE-15. Isla Cocinas (19°32'49” N, 105°06'35” O), 17 Julio 2007, intermareal rocoso, 3 machos (LE 3.6-4.1) y 7 hembras (LE 3.4-6.0), LEMA-CR-16. Isla San Pedro (19°32'10” N, 105°06'15” O), 24 Abril 2009, intermareal rocoso, 6 hembras (LE 3.75-5.28), LEMA-CR-17; 29 Noviembre 2011, manual, intermareal rocoso, 1 macho (LE 4.7mm), 1 hembra (LE 2.9mm), LEMA-CR-18.
Distribución: desde bahía Magdalena, costa oeste de Baja California Sur y el golfo de California, México hasta Paita, Perú; incluye las islas Revillagigedo, Cocos y Galápagos (Ball y Haig, 1974; Villalobos-Hiriart et al., 1989; Hendrickx y Harvey, 1999).
Comentarios: esta especie es considerada como una de las más conspicuas del ambiente intermareal rocoso. De acuerdo con Abrams (1978) y Rendón-Salinas (2000), C. compressus utiliza más frecuentemente las conchas de los gasterópodos Nerita sp. y Mancinella sp. En el presente estudio se observó un uso más frecuentemente de conchas de Nerita scabricosta Lamarck, 1822, seguida por Cantharus sanguinolentus (Duclos, 1833) (21%) y Leucozonia cerata (Wood, 1828) (29%). Con menor frecuencia se registraron las conchas de Cerithium maculosum Kiener, 1841, Mancinella speciosa (Valenciennes, 1832), Mancinella triangularis (Blainville, 1832), Nassarius corpulentus (C. B. Adams, 1852), Cantharus elegans (Griffith y Pidgeon, 1834, Ex Gray MS), Columbella fuscata Sowerby, 1832 y Nodilittorina aspera (Philippi, 1846) (Cuadro 2).
Familia Diogenidae Ortmann, 1892
Aniculus elegans Stimpson, 1859
Material examinado: El Novillo (19°33'15” N, 105°07'25” O), 28 Noviembre 2011, buceo, 9m, 1 hembra (LE 4.4mm), LEMA-CR-19.
Distribución: desde bahía Magdalena, costa oeste de Baja California Sur y el golfo de California, México hasta Ecuador (Holthuis, 1979; Snyder-Conn, 1980; Hendrickx y Harvey, 1999).
Comentarios: el único espécimen recolectado fue encontrado en una concha de Conus brunneus Wood, 1828 sobre coral duro (Pocillopora sp.), del cual se alimenta a la vez que contribuye a la producción de sedimentos calcáreos (Glynn, 1997).
Calcinas californiensis (Bouvier, 1898)
Material examinado: isla Mamut (19°32'45” N, 105°06'50” O), 27 Noviembre 2008, buceo, 10m, 1 hembra (LE 4.1mm), 1 ovígera, (LE 3.4mm), LEMA-CR-20. Isla Cocinas (19°32'49” N, 105°06'35” O), 13 Noviembre 2007, buceo, 9m, 4 machos (LE 3.03–4.14mm), LEMA-CR-21; 14 Noviembre 2007, buceo, 2 machos (LE 3.7–4.4mm); 19 Junio 2008, buceo, 8m, 3 machos (LE 3.82–5mm), LEMA-CR-22; 28 Noviembre 2011, buceo, 5m, 4 Juv., LEMA-CR-23. El Novillo (19°33'15” N, 105°07'25” O), 28 Noviembre 2011, buceo, 3m, 8 machos (LE 2.1–4.9mm), LEMA-CR-24. Isla Colorada (19°32'47” N, 105°07'29” O), 29 Noviembre 2011, buceo, 10m, 1 macho (LE 3.32mm), LEMA-CR-25. Pedreguera San Agustín (19°32'0.41” N, 105°5'26.13” O), 29 Noviembre 2011, buceo, 9m, 1 hembra (LE 2.5mm), 1 macho (LE 1.9mm), LEMA-CR-26. Isla San Pedro (19°32'10” N, 105°06'15” O), 01 Octubre 2008, buceo, 8m, 1 macho (LE 3.9mm), 1 hembra (LE 3.6mm); 29 Noviembre 2011, intermareal, 1 macho (LE 4.7mm), 1 hembra (LE 2.8mm), LEMA-CR-27.
Distribución: de bahía Magdalena, costa oeste de Baja California Sur y golfo de California, México hasta El Salvador (Poupin y Bouchard, 2006).
Comentarios: esta especie es muy parecida a C. mclaughlinaePoupin y Bouchard, 2006 y puede diferenciarse de ésta básicamente por su color en fresco. De acuerdo con Poupin y Bouchard (2006), la presencia de C. californiensis en bahía Magdalena debe ser tomada con reservas, ya que su identificación estuvo basada en material preservado y se requiere de material fresco de la localidad para confirmar su intervalo de distribución conocida.
En Los Troncones, Guerrero, C. californiensis ocupó al menos 18 especies de conchas de gasterópodos (Arce y Alcaraz, 2011), mientras que el presente estudio se encontró ocupando 8 especies, más abundantemente conchas de M. speciosa (29%) y Rhinoclavis gemmata (Hinds, 1844) (24%), y con menor frecuencia Tegula sp, M. triangularis, C. maculosum, L. cerata, Opeatostoma pseudodon (Burrow, 1815) y N. scabricosta.
Clibanarius lineatus (H. Milne-Edwards, 1848)
Material examinado: estero de Punta Pérula (19°35'06.23” N, 105°08'06.57” O), 28 Noviembre 2011, manual, 0-0.2m, 4 machos (LE 2.3–3.8mm), 1 hembra (LE 2.5mm), 1 ovígera (LE 3.5mm), LEMA-CR-28; 29 Noviembre 2011, manual, 0–0.5m, 3 machos (LE 3.4–4.0mm), 1 hembra (LE 3.6mm) LEMA-CR-29.
Distribución: desde bahía Magdalena, costa oeste de Baja California Sur y golfo de California, México hasta Capón, Perú (Hendrickx et al., 2008).
Comentarios: de acuerdo con Rahayu y Forest (1992), Clibanarius panamensis Stimpson, 1859 es un sinónimo junior de C. lineatus. Sin embargo, de acuerdo con Hendrickx et al. (2008), las descripciones de color para esta especie varían a lo largo de su distribución en el Pacífico oriental y se hace necesario realizar más estudios para determinar si las distintas poblaciones denominadas como C. lineatus corresponden en realidad a una o más especies.
Los organismos recolectados ocupaban conchas de Cerithidea cf. pulchra (C. B. Adams, 1852) y C. californica Haldeman, 1840 (Cuadro 2). En la bahía de Altata y el estero de Urías en Sinaloa, C. lineatus ocupa principalmente las conchas de Turritela gonostoma Valenciennes, 1832 (Ayón-Parente, 2009). Borjesson y Szelistowaki (1989) señalaron que en el golfo de Agua Dulce, Costa Rica la especie ocupa principalmente conchas de Thais kioskiformes (Duclos, 1832), Littoraria varia (Sowerby, 1832) y L. fasciata (Gray, 1839).
Dardanus stimpsoniAyón-Parente y Hendrickx, 2009
Material examinado: isla Colorada (19°32'47” N, 105°07'29” O), 19 Junio 2008, buceo, 9m, 1 ovígera (LE 9.5mm), LEMA-CR-30; 29 Junio 2009, buceo, 12m, 1 ovígera (LE 6.4mm), LEMA-CR-31; 29 Noviembre 2011, buceo, 10–12m, 2 macho (LE 5.4–9.5mm), 1 hembra (LE 9.2mm), 1 ovígera (LE 6.6mm), 1 Juv. NM, LEMA-CR-32. Isla Mamut (19°32'45” N, 105°06'50” O), 26 Junio 09, buceo, 10m, 1 ovígera (LE 7.9mm), LEMA-CR-33. El Novillo (19°33'15” N, 105°07'25” O), 28 Noviembre 2011, buceo, 3m, 2NS, NM, 1 ovígera (LE 8.0mm), 1 macho (LE 13.0mm), LEMA-CR-34. Rosada, 17 Julio 2007, 1 ovígera (LE 12.4mm), LEMA-CR-35. Pedreguera San Agustín (19°32'0.41” N, 105°5'26.13” O), 29 Noviembre 2011, buceo, 9m, 2 machos (LE 7.1–9.1mm), LEMA-CR-36.
Distribución: golfo de California a Guerrero, México (Ayón-Parente, 2009; Ayón-Parente y Hendrickx, 2009).
Comentarios: de las especies de Dardanus en el Pacífico mexicano, ésta es la más común y con mayor amplitud batimétrica, desde los 2 a 144m (Ayón-Parente, 2009; Ayón-Parente y Hendrickx, 2010). Las conchas ocupadas por estos organismos fueron Neorapana muricata (Broderip, 1832) (42%), Stramonita biserialis (de Blainville, 1832) (34%), Cypreacassis coarctata (Sowerby, 1825), L. cerata y Vasum caestus (Broderip, 1833), cada una con una contribución del 8%. Cabe resaltar, que en todos los casos las conchas fueron, además, usadas como sustrato para la fijación de la anémona Calliactis sp. De acuerdo con Ross (1971)Dardanus mantiene una asociación con la anémona Calliactis sp., la cual coloca sobre su concha y obtiene el beneficio de camuflaje, protección contra depredadores y cubierta cuando utiliza conchas débiles.
Paguristes sp. 1
Material examinado: isla Cocinas (19°32'49” N, 105°06'35” O), 03 Octubre 2008, buceo, 10m, 1 ovígera (LE 5.12mm), EMU-9695.
Comentarios: se descubrió que la hembra recolectada en el presente estudio parece corresponder a una especie no descrita, muy similar a Paguristes digueti Bouvier, 1892 y P. sanguinimanusGlassell, 1938 (Ayón-Parente, en proceso). Fue recolectada en una concha de Conus brunneus.
Paguristes sp. 2
Material examinado: isla Colorada (19°32'47” N, 105°07'29” O), 29 Noviembre 2011, buceo, 12m, 1 hembra (LE 4.3mm), 1 ovígera (LE 3.8mm), LEMA-CR-37. Pedreguera San Agustín (19°32'0.41” N, 105°5'26.13” O), 29 Noviembre 2011, buceo, 9m, 2 machos (LE 2.2–4.2mm), 2 hembra (LE 2.4–3.5mm), 1 ovígera (LE 3.2–mm), 3 SS, SM LEMA-CR-38; 1 hembra (LE 5.9mm), 1 macho (LE 5.3mm), EMU-9697.
Comentarios: los ejemplares también corresponden a una especie nueva para la ciencia. Por la morfología de las escamas oculares, el diente rostral y los quelípedos, esta especie puede ser incluida dentro del complejo de especies de Paguristes anahuacusGlassell, 1938. Presenta mayor similitud morfológica con P. anahuacus, pero pueden diferenciarse fácilmente por el patrón de coloración. En Paguristes sp. 2 los pedúnculos oculares son crema y azul china sobre la mitad distal vs. crema con una banda café subdistal; los apéndices ambulatorios y quelípedos son de color naranja, mientras que en P. anahuacus son de un color crema uniforme. Las conchas más utilizadas por esta especie fueron O. pseudodon y C. maculosum (33% cada una), también se recolectaron en conchas de L. cerata, Cymatium lignarium (Broderip, 1833) y Bursa sp.
Trizopagurus magnificus (Bouvier, 1898)
Material examinado: isla Cocinas (19°32'49” N, 105°06'35” O), 13 Noviembre 2007, buceo, 9m, 1 macho (LE 7.25mm), LEMA-CR-39; 03 Octubre 2008, buceo, 9m, 3 machos (LE 5.8–8.8mm), LEMA-CR-40; 28 Noviembre 2011, 5m, snorkel, 1 macho (LE 1.6mm), LEMA-CR-41. El Novillo (19°33'15” N, 105°07'25” O), 28 Noviembre 2011, buceo, 5m, 1 hembra (LE 3.7mm), LEMA-CR-42.
Distribución: desde bahía Magdalena, costa oeste de Baja California Sur y bahía Santa María, Sinaloa hasta el norte de Perú, incluyendo las islas Malpelo y Galápagos (Forest, 1995; Hendrickx y Harvey, 1999; Hendrickx et al. 2008).
Comentarios: esta especie es relativamente común en las localidades donde hay presencia del coral duro (Pocillophora) del cual se ha documentado que se alimenta (Guzmán, 1988). Sin embargo, en las localidades donde no existen formaciones coralinas se desconoce cuál es la dieta específica de este ermitaño. Es una especie común en la bahía Chamela. Se le encontró ocupando conchas de S. biserialis y Homalocantha cf. oxyacantha (Broderip, 1833) y C. elegans, sobre formaciones coralinas.
Familia Paguridae Latreille, 1803
Pagaras benedicti (Bouvier, 1898)
Material examinado: pedreguera San Agustín (19°32'0.41” N, 105°5'26.13” O), 29 Noviembre 2011, buceo, 9m, 1 macho (LE 1.8mm), 1 hembra (LE 1.8mm). LEMA-CR-43.
Distribución: desde bahía Magdalena, costa oeste de Baja California Sur y golfo de California, México hasta Ecuador (McLaughlin y Haig, 1993).
Comentarios: Pagurus benedicti se diferencia fácilmente del resto de las especies del grupo Provenzanoi con distribución en el Pacífico mexicano, en esta especie las escamas oculares terminan en un solo diente, mientras que en las demás especies las escamas oculares están armadas con 2–6 dientes. Los 2 organismos recolectados ocupaban conchas de Favartia lappa (Broderip, 1833).
Pagurus lepidus (Bouvier, 1898)
Material examinado: Punta Pérula (19°34'53.71” N, 105°8'4.41” O), 24 Junio 09, buceo, 3m, 1 macho (LE 2.1mm), 1 ovígera (LE 1.9mm), LEMA-CR-44. Distribución: costa oeste de Baja California Sur y golfo de California, México a Perú (Haig y McLaughlin, 1991; Vargas y Cortés, 2006).
Comentarios: Pagurus lepidus es la especie del grupo Provenzanoi más ampliamente distribuida en el Pacífico este tropical. De acuerdo con Haig y McLaughlin (1991), ésta es una especie de coloración variable a lo largo de su distribución geográfica. Los organismos recolectados en bahía Chamela corresponden a la población sureña, caracterizada por llevar flagelos antenales usualmente con 3 artículos de color verde oliva interrumpidos por un artículo blanco, presentan también escudo moteado de color verde oliva y blanco, pedúnculos oculares moteados de color verde oliva y blanco con córnea roja, los pedúnculos antenulares presentan manchas blancas dorsalmente y bandas verdes proximalmente sobre el último segmento, con una banda verde oscura distalmente sobre el penúltimo segmento, quelípedos con dáctilo blanco y el dedo fijo, palma, carpo y mero moteados de verde oliva y blanco. Fue recolectado en conchas de C. elegans y R. gemmata.
Pagurus nanodes Haig y Harvey, 1991
Material examinado: pedreguera San Agustín (19°32'0.41” N, 105°5'26.13” O), 29 Noviembre 2011, buceo, 9m, 1 macho (LE 1.4mm), EMU-9698; mismos datos, 1 ovígera (LE 1.6mm), LEMA-CR-45.
Distribución: Costa Rica a Ecuador (Haig y Harvey, 1991; Hendrickx y Harvey, 1999). En México solamente se había recolectado en Mazatlán (Ayón-Parente y Hendrickx, 2005).
Comentarios: este es el segundo registro de la especie para México (3.6 grados al sur de Mazatlán) e incluye una extensión en su distribución batimétrica de 3.5 a 9m. Los organismos se encontraron ocupando conchas de Anachis scalarina (G.B. Sowerby, 1832) y Pilsbryspira garciacubasi Shasky, 1971.
Phimochirus roseus (Benedict, 1892)
Material examinado: isla Cocinas (19°32'49” N, 105°06'35” O), 28 Noviembre 2011, buceo, 5m, 3 machos (LE 4.15.1mm), 2 ovígeras (LE 3.3–3.5mm), LEMA-CR-46. Isla San Andrés (19°31'22.94” N, 105°5'11.92” O), 26 Noviembre 2008, buceo, 9m, 1 hembra (LE 3.2mm), LEMA-CR-47. Isla Colorada (19°32'47” N, 105°07'29” O), 19 Junio 2008, buceo, 9m, 1 ovígera (LE 5.3mm), LEMA-CR-48; 29 Noviembre 2011, buceo, 10m, 1 macho (LE 4.0mm), 1 ovígera (LE 3.7mm), LEMA-CR-49.
El Novillo (19°33'15” N, 105°07'25” O), 28 Noviembre 2011, buceo, 5m, 1 macho (LE 5.0mm), 2 hembras (LE 2.6mm), 1 ovígera (LE 4.9mm), LEMA-CR-50; mismos datos, 1 macho (LE 5.4mm), 1 hembra (4.5mm), EMU-9694. Pedreguera San Agustín, 29 Noviembre 2011, buceo, 9m, 1 macho (LE 3.0mm), 1 hembra (LE 3.8mm), LEMA-CR-51.
Distribución: de bahía Magdalena, costa oeste de Baja California Sur, isla San Pedro Mártir y bahía La Cholla, Sonora; golfo de California, México y Costa Rica a Ecuador (McLaughlin, 1982; Hendrickx y Harvey, 1999; Lemaitre y McLaughlin, 2003).
Comentarios: los organismos de bahía Chamela representan el primer registro de la especie en México al sur del golfo de California. El material fue recolectado entre los 5 y 10m de profundidad, lo que representa un nuevo registro batimétrico, la especie había sido recolectada previamente de 0 a 3m. Los especímenes ocuparon con mayor frecuencia conchas de C. sanguinolentus y L. cerata, con menor frecuencia, Muricopsis zeteki Hertlein y Strong, 1951, Bursa corrugata corrugata (Perry, 1811), C. maculosum, M. speciosa, C. elegans y Tegula sp.
DiscusiónLas 12 especies de cangrejos ermitaños registrados en este estudio constituyen un 13.3% de las especies conocidas en la costa occidental mexicana y 9 de ellas representan nuevos registros para la localidad. El hallazgo de P. nanodes en Chamela corresponde al segundo registro de la especie en México y una ampliación en su distribución batimétrica a partir del reporte de Ayón-Parente y Hendrickx (2005), mientras que la captura de P. roseus representa el segundo registro en México fuera del golfo de California (Hendrickx y Harvey, 1999). Al igual que en la costa occidental mexicana, la familia Diogenidae fue la mejor representada en el área de estudio, con 7 especies. El género mejor representado fue Pagurus con 3 especies, seguido de Paguristes con 2 y el resto de los géneros tuvieron una sola especie. Coenobita compressus fue la única especie presente en el ambiente supralitoral. En el estero de Pérula, solamente se recolectó a C. lineatus y las especies restantes fueron capturadas en la bahía tanto en la porción intermareal como submareal (Cuadro 1).
Utilización de conchas vacías como hábitat. Los ermitaños de bahía Chamela utilizaron conchas de 29 especies de gasterópodos como habitáculo. Entre las especies de ermitaños más abundantes se pudo observar cierta preferencia hacia algunos tipos de conchas. Los ejemplares de C. compressus ocuparon 10 especies distintas, de éstas N. scabricosta y C. sanguinolentus fueron las más utilizadas (29% cada una), C. californiensis y P. roseus utilizaron 8 conchas distintas cada uno; C. californiensis mostró preferencia por M. speciosa (29%) y R. gemmata (24%), mientras que P. roseus lo hizo por C. sanguinolentus y L. cerata (21% cada una). Por su parte, C. lineatus utilizó solamente conchas de C. cf. pulchra (70%) y Cerithidea californica (30%), comunes en ambientes estuarinos. En realidad solamente 7 de las 29 especies de gasterópodos registrados como habitáculo fueron las mayormente seleccionadas, representando casi el 54%, mientras que 15 gasterópodos fueron elegidos sólo en 1 o 2 ocasiones. Las 7 primeras se encuentran entre las más comunes en los ambientes intermareal y submareal somero de la bahía. Reese (1963) señala algunas de las características que influyen en la selección de la concha, como son: contraste con el fondo, movilidad, textura superficial, forma externa, la presencia de abertura, la abertura libre de obstrucciones, tamaño interno, concha con abertura hacia abajo, configuración interna y el peso. Rendón-Salinas (2000) menciona que los ermitaños seleccionan las conchas con base en su arquitectura y volumen, mientras que otros estudios indican que el uso de conchas está determinado principalmente por la disponibilidad de conchas y no necesariamente por la preferencia (Botelho y Costa, 2000; Barnes, 2005). En la bahía se han registrado al menos 215 especies de gasterópodos (Bastida-Izaguirre, datos no publ.). Sin embargo, un gran número de estas especies son raras, de tamaño muy pequeño o grande no apropiado para los cangrejos ermitaños, por lo que se requieren estudios más detallados que permitan determinar si la disponibilidad de conchas es un factor limitante para su selección por parte de las especies de ermitaños.
Elwood et al. (1979) y Elwood y Kennedy (1988) señalaron que la selección de un determinado tipo y forma de concha por parte de los cangrejos ermitaños tiene como consecuencia modificaciones en la morfología del cangrejo, por ejemplo, moldear y modificar su cefalotórax y abdomen. Algunas alteraciones morfológicas importantes son el resultado de la arquitectura de las conchas utilizadas (McLaughlin y Bailey-Brock, 1975; Vermeij, 1978). Por ejemplo, Glassell (1937, 1938) señala que Paguristes digueti usualmente habita conchas de Strombus y presenta una depresión de la porción precervical en el caparazón y un distendimiento de las regiones branquiales. Esto pudo observarse en Paguristes sp. 1, recolectada en una concha del género Conus que presentó un aplastamiento dorsoventral de cefalotórax y abdomen.
Con los resultados de este trabajo se incrementa el conocimiento de la riqueza de especies de ermitaños de bahía Chamela, lo que refleja la necesidad de aumentar el esfuerzo en el estudio de la biodiversidad del santuario para garantizar su conservación y manejo sustentable.
A los estudiantes y prestadores de servicio social que apoyaron este trabajo, en especial a Arizbeth Alonso y Ana Barragán. A Jorge Vega, jefe de la estación de Biología Chamela del IBUNAM, por todas las facilidades otorgadas durante las salidas de campo. DBI agradece al Conacyt por la beca No. 15477 otorgada para estudios de posgrado; asimismo, MAP agradece al Conacyt-México por el apoyo recibido durante su estancia postdoctoral en el CUCBA, Universidad de Guadalajara. Este proyecto fue financiado por la Universidad de Guadalajara a través del proyecto P3E 2011. A los revisores por los comentarios y sugerencias hechas al manuscrito.