




Se aportan datos de 10 nuevos registros que amplían el área de distribución de la rana ladrona pigmea hacia los estados de Jalisco, Nayarit y Sinaloa.
We provide data on 10 new records that expand the geographic range of the pigmy robber frog into the states of Jalisco, Nayarit, and Sinaloa.
Craugastor pygmaeus (Taylor, 1937) se distribuye discontinuamente desde el sureste de Michoacán hasta el extremo oeste de Guatemala por la vertiente del Pacífico y también en Puebla, Oaxaca, Tabasco y Veracruz por la vertiente del Atlántico, en un intervalo altitudinal desde 0 hasta más de 2 000m snm (Santos-Barrera y Canseco-Márquez, 2004; Stuart et al., 2008). En México se le ha registrado en los estados de Chiapas, México, Guerrero, Michoacán, Oaxaca, Puebla, Tabasco y Veracruz (Lynch, 1965; García-Vázquez et al., 2009). Sin embargo, no existen datos de su presencia en los estados de Jalisco, Nayarit y Sinaloa. En esta nota se presentan los primeros registros de C. pygmaeus para estos 3 estados. La mayoría de estos registros provienen de 15 muestreos realizados entre los años 2008 y 2011 en Nayarit y Jalisco. Otros registros para Nayarit y Sinaloa se obtuvieron de la base de datos en línea de la colección herpetológica de la California Academy of Sciences (CAS, 2013).
El acrónimo UTA-DC corresponde a la University of Texas at Arlington-Digital Collection. Los registros basados en fotografías fueron verificados por J. W. Streicher (Fig. 1). Las coordenadas aparecen en grados decimales con datum de referencia WGS84 y la altitud en m snm. Los registros nuevos para C. pygmaeus se listan a continuación.
. B, sierra del Cuale, Jalisco (UTA-DC 6959). C, camino a Las Guacas, Jalisco (UTA-DC 6960). D, ejido Emiliano Zapata, Jalisco (UTA-DC 6961). E, río Tacubaya, Jalisco (UTA-DC 6962). F, sierra de Vallejo, Nayarit (UTA-DC 6957). G, Santa Cruz de Miramar, Nayarit (UTA-DC 6971). H, Concordia, Sinaloa (CAS 175705). Las divisiones menores en la escala de la Fig. 1H representan 1mm.")
Ejemplares de Craugastor pygmaeus registrados en el presente trabajo. A, sierra de Quila, Jalisco (UTA-DC 6958). B, sierra del Cuale, Jalisco (UTA-DC 6959). C, camino a Las Guacas, Jalisco (UTA-DC 6960). D, ejido Emiliano Zapata, Jalisco (UTA-DC 6961). E, río Tacubaya, Jalisco (UTA-DC 6962). F, sierra de Vallejo, Nayarit (UTA-DC 6957). G, Santa Cruz de Miramar, Nayarit (UTA-DC 6971). H, Concordia, Sinaloa (CAS 175705). Las divisiones menores en la escala de la Fig. 1H representan 1mm.
Jalisco. Municipio de Tecolotlán: Área Natural Protegida Sierra de Quila (UTA-DC 6958; 20.301525°, −104.03462778°; 2 145m snm), 17 de julio de 2010; el ejemplar se observó entre la hojarasca en la ribera de un arroyo, en bosque de pino. Municipio de Talpa de Allende: sierra del Cuale, 22km al sur de Talpa (UTA-DC 6959; 20.226261°, −104.76245000 ° 1 571m snm), 17 de abril de 2011, bosque mesófilo de montaña. Municipio de San Sebastián del Oeste: 7km al oeste de La Estancia (UTA-DC 6960; 20.761639°, −104.95788889°; 834m snm), 19 de abril de 2011; el ejemplar se observá en la ribera de un arroyo perenne que cruza el camino hacia Las Guacas; ecotono entre bosque de pino y bosque tropical subcaducifolio. Municipio de Cabo Corrientes: Ejido Emiliano Zapata (UTA-DC 6961; 20.443169°, − 105.27650000°; 426m snm), 22 de abril de 2011, bosque tropical subcaducifolio. Municipio de Casimiro Castillo: río Tacubaya, 1km al sureste de Casimiro Castillo (UTA-DC 6962; 19.591939°, −104.42641111°; 413m snm), 24 de abril de 2011, bosque tropical subcaducifolio.
Nayarit. Municipio de Bahía de Banderas: sierra de Vallejo, 6km al sur de Sayulita por la carretera federal 200 (UTA-DC 6957; 20.819956°, −105.39356944°; 160m snm), 11 abril de 2009, bosque tropical subcaducifolio. Municipio de San Blas: 2.46km al este de Santa Cruz de Miramar (UTA-DC 6971-72; 21.429132°, −105.17345000°; 98m snm), 17 de abril de 2008, bosque tropical subcaducifolio; 5km al noreste de San Blas (CAS 95663; 21.56°, − 105.24°; 20m snm), 9 de junio de 1963, bosque tropical subcaducifolio. Municipio de Santa María del Oro: 6.59km al este sobre el camino a Santa María del Oro desde de la intersección con la carretera federal 15 (CAS 99231; 21.34°, −104.61°; 1 164m snm), 28 de mayo de 1963, bosque tropical subcaducifolio.
Sinaloa. Municipio de Concordia: carretera Durango-Mazatlán, 40km al este de la intersección de las carreteras federales 15 y 40, de ahí 0.48km (en línea recta) al sur de la carretera federal 40 (CAS 175697-716; 23.31°, −106.01°; 186m snm), 9 de junio de 1963, bosque tropical subcaducifolio.
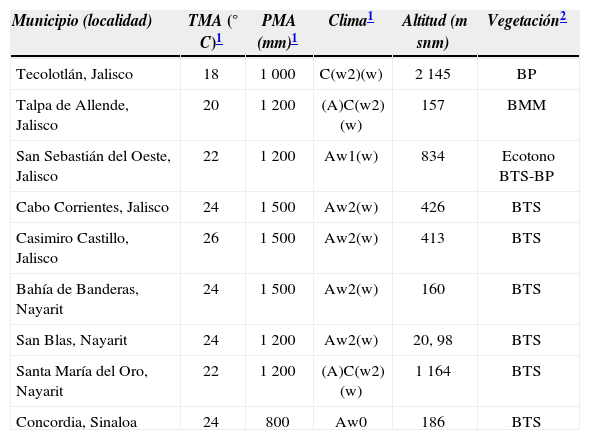
Estas 10 localidades incrementan de 8 a 11 el número de estados de México en los cuales se registra la presencia de C. pygmaeus (Fig. 2). Asimismo, amplían el área de distribución geográfica de la especie alrededor de 671km(en línea recta) hacia el noroeste desde la localidad más cercana, que es Arteaga, Michoacán (Duellman, 1961). Con los registros nuevos, la distribución de C. pygmaeus en el occidente de México incluye ahora 5 municipios de Jalisco, 3 de Nayarit y 1 de Sinaloa, en intervalos de altitud de 20 a 2 145m snm, de precipitacián media anual de 800 a 1 500fmm, y de temperatura media anual de 18 a 26° C (Cuadro 1). El registro del municipio de Concordia, Sinaloa representa la localidad más septentrional para la especie.
.")
Distribucián geográfica de Craugastor pygmaeus en los estados de Jalisco, Nayarit y Sinaloa. Los círculos negros representan los registros de los autores y los cuadros negros representan los ejemplares depositados en la colección de CAS. En rojo se muestra la distribución de C. pygmaeus de acuerdo con la IUCN Red List of Threatened Species (Santos-Barrera y Canseco-Márquez, 2004).
Variables ambientales asociadas a las nuevas localidades donde se reporta Craugastor pygmaeus en el occidente de México
| Municipio (localidad) | TMA (° C)1 | PMA (mm)1 | Clima1 | Altitud (m snm) | Vegetación2 |
|---|---|---|---|---|---|
| Tecolotlán, Jalisco | 18 | 1 000 | C(w2)(w) | 2 145 | BP |
| Talpa de Allende, Jalisco | 20 | 1 200 | (A)C(w2)(w) | 157 | BMM |
| San Sebastián del Oeste, Jalisco | 22 | 1 200 | Aw1(w) | 834 | Ecotono BTS-BP |
| Cabo Corrientes, Jalisco | 24 | 1 500 | Aw2(w) | 426 | BTS |
| Casimiro Castillo, Jalisco | 26 | 1 500 | Aw2(w) | 413 | BTS |
| Bahía de Banderas, Nayarit | 24 | 1 500 | Aw2(w) | 160 | BTS |
| San Blas, Nayarit | 24 | 1 200 | Aw2(w) | 20, 98 | BTS |
| Santa María del Oro, Nayarit | 22 | 1 200 | (A)C(w2)(w) | 1 164 | BTS |
| Concordia, Sinaloa | 24 | 800 | Aw0 | 186 | BTS |
TMA= Temperatura media anual, PMA= Precipitación media anual, BP= Bosque de Pino, BMM= Bosque Mesófilo de Montaña, BTS= Bosque Tropical Subcaducifolio.
Los individuos de C. pygmaeus de Jalisco y Nayarit comparten similitudes en el microhábitat que utilizan. Estos individuos se observaron en las márgenes de arroyos perennes donde existía acumulacián de materia orgánica; entre la hojarasca, rocas y troncos en descomposición. Tales características de su microhábitat fueron previamente señaladas por Taylor (1937), Duellman (1961) y Davis y Dixon (1965) para las poblaciones de Veracruz, Michoacán y Guerrero, respectivamente. Cabe mencionar que los ejemplares CAS 95663, 99231 y 175697-716 fueron recolectados por A. E. Greer y F. W. Burley durante expediciones realizadas a los estados de Nayarit y Sinaloa e identificados en la CAS por J. D. Lynch como Eleutherodactylus pygmaeus, actualmente C. pygmaeus (Crawford y Smith, 2005).
De acuerdo con Hedges et al. (2008), C. pygmaeus es una de las 7 especies que pertenecen a la serie mexicanus del género Craugastor. Esta serie de especies se caracteriza porque todos los representantes carecen de tubérculos plantares y también de ranuras o sacos vocales (aunque están presentes en C. occidentalis). Lynch (1965, 1970) indica que C. pygmaeus difiere de todos los miembros de la serie mexicanus, exceptuando a C. hobartsmithi, en poseer 2 tubérculos palmares (3 tubérculos palmares en las otras especies). A su vez Taylor (1940) y Lynch (1965) señalan que C. pygmaeus difiere de C. hobartsmithi, ya que la primera especie carece de una serie de tubérculos a lo largo del borde externo de ambos tarsos (uno o más tubérculos tarsales en C. hobartsmithi). Además, porque C. hobartsmithi presenta glándulas parotoides y carece de manchas inguinales oscuras, mientras que C. pygmaeus carece de las glándulas parotoides, pero en las ingles sí presenta manchas. Tanto la presencia de 2 tubérculos palmares, ausencia de tubérculos tarsales, ausencia de glándulas parotoides y presencia de manchas oscuras en las ingles fueron verificadas en cada uno de los ejemplares observados en el campo.
Aunque todos los ejemplares fotografiados concuerdan con la diagnosis de Taylor (1937), la mayoría presentan una mancha interescapular en forma de “V' invertida (Fig. 1; exceptuando Fig. 1G), es importante señalar el policromatismo entre las poblaciones del occidente de México. Las variaciones cromáticas inter e intrapoblacionales de C. pygmaeus fueron analizadas por Duellman (1961) en una serie de 492 ejemplares provenientes de Chiapas, Michoacán, Oaxaca, Tabasco y Veracruz, en los cuales encontró al menos 6 patrones distintivos que se presentan aleatoriamente entre las poblaciones del sureste mexicano. Los individuos de las Figs. 1B, E, F y H concuerdan con las características del patrán tipo “A” descrito por Duellman (1961), mientras que los individuos de las Figs. 1A, C y D concuerdan con el patrán tipo “B” y el ejemplar de la Fig. 1G presenta el patrán tipo “D”. Las observaciones de Duellman (1961) sirvieron como evidencia para que Lynch (1965) invalidara algunos taxones morfolágicamente relacionados con C. pygmaeus, englobando todos en una sola especie polimárfica (i. e. Craugastor C. pygmaeus).
Sin embargo, un análisis molecular reciente, sugiere la existencia de diversidad críptica en C. pygmaeus (Streicher, 2012). Pero este análisis no incluyá muestras de 7 de los estados de México donde se distribuye C. pygmaeus; entre ellos Nayarit y Sinaloa y sálo incluyá un par de muestras de una misma poblacián (Talpa de Allende) de Jalisco. Si bien es posible que C. pygmaeus represente un complejo de especies, los individuos que se muestran en este estudio son diagnosticables morfolágicamente como C. pygmaeus (sensuTaylor 1937; Lynch, 1965) por un lado, y por el otro, hacen falta análisis moleculares y morfológicos con un muestreo mucho más extensivo para reevaluar los límites de especies en C. pygmaeus. Ya que la evidencia de Streicher (2012) es mínima como para elucidar completamente las relaciones filogenéticas y la taxonomía de esta especie polimórfica.
Agradecemos a D. Carrasco-Fernández, D. Estrada, O. Reyna y H. Leal por su apoyo en campo; a R. C. Drewes por facilitarnos los datos de los ejemplares depositados en CAS y enviarnos la fotografía de la Fig. 1H; a J. Reyes-Velasco por el apoyo en la catalogación de las fotografías en UTA-DC; a J. W. Streicher por verificar la identidad de los ejemplares y a U. O. García-Vázquez por sus sugerencias a una versión previa de esta nota.