


Studies on helminths of chiropterans are relatively uncommon compared to those of other animals, and seasonal changes in helminth load have been rarely examined. We characterized the gastro-intestinal helminth load of 3 bats species to test for the existence of seasonal changes in response to known seasonal environmental and bat prey fluctuations. We did not find seasonal variation in most of the cases. However, the prevalence of 4 endoparasite species was significantly higher during one of the seasons. The highest richness was registered in Pteronotus parnellii during the wet season. The effective number of species was higher during the dry season in the 3 species of Pteronotus. Diet seems to be an important driver of helminth infracommunity structure, but we found heterogeneous patterns in the relationship between diversity and load of helminths and seasonal patterns of bat's diets and abundance of potential intermediate hosts.
Las investigaciones sobre los helmintos de quirópteros son relativamente escasas en comparación con las de otros vertebrados y los cambios estacionales en su carga han sido poco estudiados. En este estudio se caracterizó la carga de helmintos gastrointestinales de 3 especies de murciélagos para probar la existencia de cambios estacionales en respuesta a las fluctuaciones ambientales y de presas. No se encontró variación estacional en la mayoría de los casos. Sin embargo, la prevalencia de 4 especies de endoparásitos fue significativamente mayor durante una de las épocas. La mayor riqueza de especies se registró en P. parnellii durante la época de lluvias. El número efectivo de especies fue mayor durante estaciones secas en las 3 especies de Pteronotus. La dieta parece dirigir la estructura de las infracomunidades de helmintos, aunque se encontraron patrones heterogéneos en la relación entre la diversidad y la carga de helmintos y los patrones estacionales de la dieta y la abundancia de los posibles huéspedes intermediarios.
All organisms, including parasites, are influenced directly or indirectly by environmental variations (Marcogliese, 2001; Pilosof, Dick, Korine, Patterson, & Krasnov, 2012). The transmission, development and distribution of parasites can be regulated by abiotic factors (Brooks & Hoberg, 2007; Gotz, Harf, Sommer, & Matthee, 2010). In particular, dynamics of helminths are regulated by environmental conditions such as ambient temperature, humidity and precipitation (Appleton & Gouws, 1996; Doi & Yurlova, 2011; Hudson, Cattadori, Boag, & Dobson, 2006; Mouritsen & Poulin, 2002; Moyer, Drown, & Clayton, 2002; Tinsley et al., 2011). A few studies have also suggested that seasonal variation in parasite communities is influenced by biotic factors such as abundance, diet, reproductive behavior, and immunocompetence of hosts (Carvalho & Luque, 2011; Esch & Fernández, 1993; Felis & Esch, 2004; Šimková, Jarkovsky, Koubková, Barus, & Prokes, 2005).
Bats are one of the most diverse and widespread of mammal orders (Altringham, 1996; Wilson & Reeder, 2005). Studies of helminths in chiropteran populations are relatively uncommon compared to those of other animals (Kirschbaum, Perkins, & Gannon, 2009; Lord, Parker, Parker, & Brooks, 2012). Chiropterans harbor a great variety of helminths, including trematodes, cestodes, and nematodes (Cuartas-Calles & Muñoz-Arango, 1999; Lord et al., 2012) and most studies consist of checklists of species and new descriptions of host or localities (Guzmán-Cornejo, García-Prieto, Pérez-Ponce de León, & Morales-Malacara, 2003; McAllister, Bursey, & Dowler, 2007; Muñoz et al., 2011; Nahhas, Yang, & Uch, 2005; Nogueira, de Fabio, & Peracchi, 2004; Shimalov, Demyanchik, & Demyanchik, 2002). Nevertheless, a few studies have explored the effect of seasonal variation in intensity and prevalence of bat endoparasites (Blankespoor & Ulmer, 1970; Coggins, Tedesco, & Rupprecht, 1982; Lord et al., 2012; Nickel & Hansen, 1967). For example, studies with insectivorous bats have reported that prevalence and intensity of helminths are low during the spring, increase in the summer and reach a peak in the autumn (Blankespoor & Ulmer, 1970; Nickel & Hansen, 1967). Among other factors intrinsic to host biology, seasonal changes in endoparasite abundance might be related to increased abundance of arthropods that act as intermediate hosts and that are then ingested by bats (Lord et al., 2012).
Trematodes are the most diverse group of helminths found in bats (Coggins, 1988; Ubelaker, 1970). They are found mainly within the gastrointestinal tract and in other body cavities (Coggins, 1988; Ricci, 1995; Shimalov et al., 2002), and their incidence and prevalence are affected by the host's feeding habits (Coggins, 1988; Marshall & Miller, 1979). For example, most digenean species (trematodes) have been collected in insectivorous bats since they are more prone to ingest infected insects (intermediate hosts) than nectar or fruit feeding bats (Coggins, 1988; García-Vargas, Osorio, & Pérez-Ponce de León, 1996; Lord et al., 2012; Ubelaker, 1970). Studies with other vertebrates (e.g., fishes) have reported that the diet of the host determines the abundance and richness of helminths (Bell & Burt, 1991; Poulin & Morand, 2004; Šimková et al., 2005).
In this study, we investigated the seasonal variation of the endoparasitic load in 3 insectivorous [Pteronotus davyi (Gray), P. parnellii (Gray), and P. personatus (Wagner)] bat species. Previous studies have reported the helminthological record of these bats species in Mexico (Caballero-Caballero & Zerecero, 1942; Espericueta-Viera, 2012; García-Vargas et al., 1996; Guzmán-Cornejo et al., 2003; Peralta-Rodríguez, Caspeta-Mandujano, & Guerrero, 2012; Pérez-Ponce de León, León-Régagnon, & García-Vargas, 1996), but few have examined seasonal variations of infection patterns. For example, Clarke (2008) found no seasonal variation in endoparasite species composition of P. davyi and P. personatus and no seasonal difference in prevalence and abundance in a related species (Mormoops megalophylla) in a tropical deciduous forest in southern Mexico.
The study was conducted in a highly seasonal dry forest. Tropical dry forests have extreme changes in the physiognomy and availability of food resources during the wet and dry seasons, affecting the composition and diversity of fauna (Castaño-Meneses, 2014). For instance, the abundance of arthropods in tropical dry forests experiences considerable seasonal fluctuations, reaching its highest level during the wet season (Andresen, 2005; Castaño-Meneses, 2014; Güizado & Casas-Andreu, 2011; Leavings & Windsor, 1984), a pattern that has been previously reported for the study region (Pescador-Rubio, Rodríguez-Palafox, & Noguera, 2002).
A previous study using DNA barcodes showed that the diet of the 3 species of Pteronotus considered in our study is more diverse during the dry season (Salinas-Ramos, Montalvo, León-Regagnon, Arrizabalaga-Escudero, & Clare, 2015). For completing their transmission, some species of helminths use arthropods as intermediate hosts (Bush, Fernández, Esch, & Seed, 2001; Clarke, 2008) and insectivorous bats as definite host (Chitwood, 1938; García-Vargas, 1995). Accordingly, we predicted that the endoparasite load in insectivorous bats would exhibit seasonal changes, having the highest richness during the dry season (spring), when their diet is more diverse (Salinas-Ramos et al., 2015). In contrast, we expected that the prevalence, abundance and intensity of helminths would be higher in the rainy season, when the abundance of intermediate hosts peaks (Lord et al., 2012).
Material and methodsThe 3 focal bat species roost in a cave in San Panchito Island, off the Pacific coast in Jalisco, Mexico (19°32′6″N, 105°5′17.9″W). The adjacent continental region is composed of tropical deciduous and tropical semideciduous forest (Rzedowski, 1981), with most of the rainfall occurring from July to November (Bullock, 1995; Méndez-Alonzo, Pineda-García, Paz, Rosell, & Olson, 2013; Pringle, Dirzo, & Gordon, 2012). We carried out 3 collecting trips during the dry season (spring: June 2012, April 2013, May 2014) and 4 in the wet season (summer: July 2013; autumn: November 2012, November 2013 and September 2014). Bats were collected with mist nests at sunset and with sweep nets inside the cave during the morning. All the specimens captured were adults and we held each individual in a cotton bag. Bats were transported to the Estación de Biología Chamela.
Bats were sacrificed with chloroform and deposited as voucher specimens in the National Mammal Collection (CNMA) of the Instituto de Biología and in the Mammal Collection of the Zoology Museum, Universidad Nacional Autónoma de México (MZFC, UNAM). The gastrointestinal tract was dissected from each bat and immersed in phosphate-buffered saline solution in a Petri dish. The stomach and intestine were examined carefully using a stereo microscope (Leica Microsystems, ES2, Wetzlar, Germany). All the helminths were fixed by sudden immersion in hot 4% formalin and preserved in 70% ethanol. Specimens were stained with Mayer's paracarmine, dehydrated, cleared in methyl salicylate and mounted in Canada balsam (Lamothe-Argumedo, 1997). Morphological data were cross-referenced with the available literature on species known to be present in bats (García-Vargas, 1995; Justine, 1989; Moravec, 1982; Vaucher, 1992; Vaucher & Durette-Desset, 1986).
In order to characterize the seasonal variation in endoparasite load, we calculated the following descriptors of parasite populations: prevalence, abundance and intensity in the dry and wet seasons. We performed a chi-squared test with the prevalence data and a bootstrap test with the mean abundance and mean intensity values of each endoparasite species to evaluate significant differences between the seasons. We also calculated for each bat species the proportion of hosts infected considering the total endoparasite records, the average number of total endoparasites per host individual examined, and the average number of total endoparasites per infected host individual. Statistical analyses were conducted in Quantitative Parasitology (Version, 3.0, Reiczigel & Rózsa, 2005), and the significance level was set at p≤0.05.
Richness, Shannon–Wiener and Simpson–Gini indexes were calculated to assess the diversity of endoparasite species during both seasons for each bat species. We transformed index values to the effective number of species (Hill, 1973; MacArthur, 1965) in order to unify an intuitive interpretation of diversity (Jost, 2006). Descriptive analyses and graphics were conducted in GraphPad Prism (Version 6.0 for Windows, GraphPad Software, La Jolla, CA, USA).
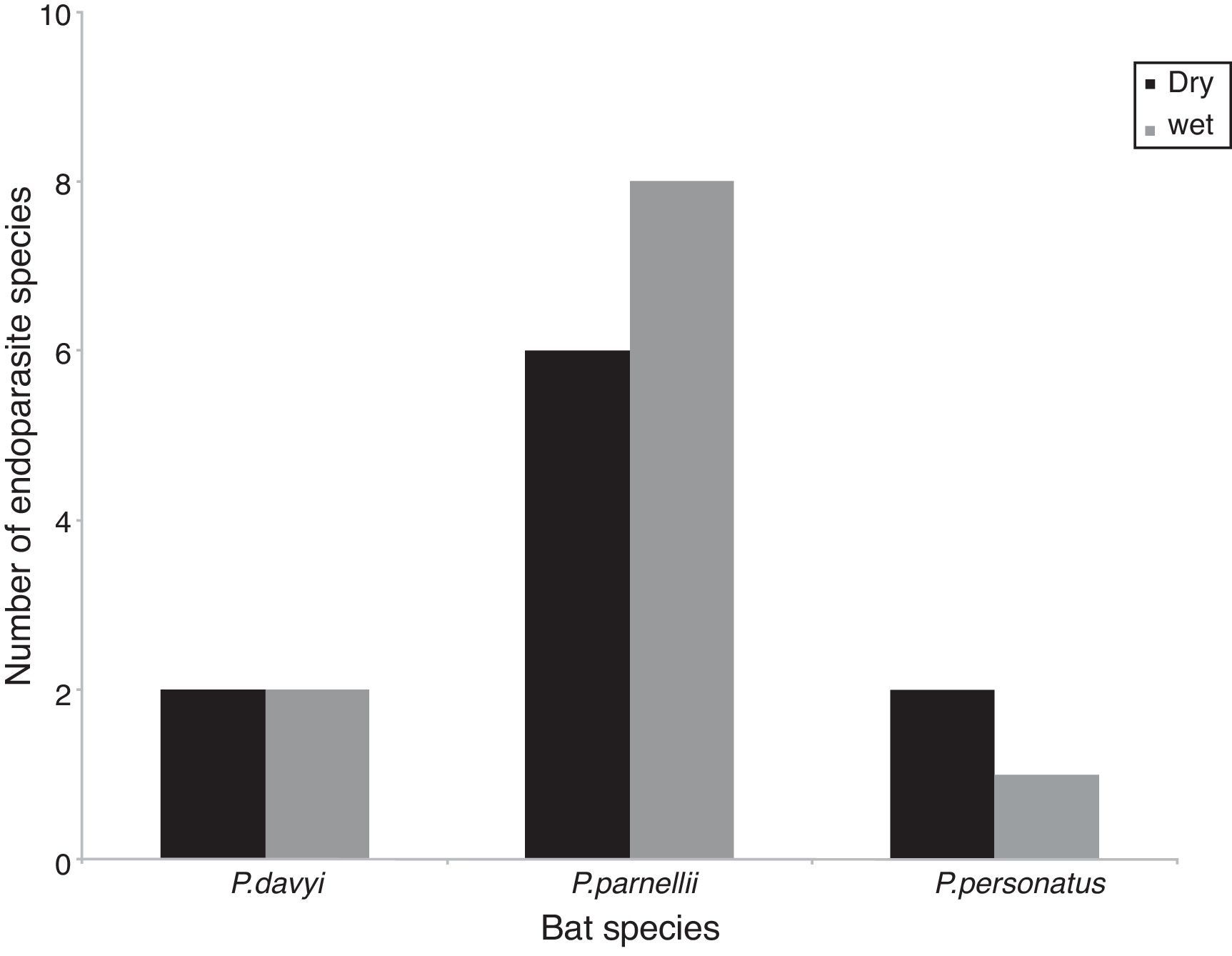
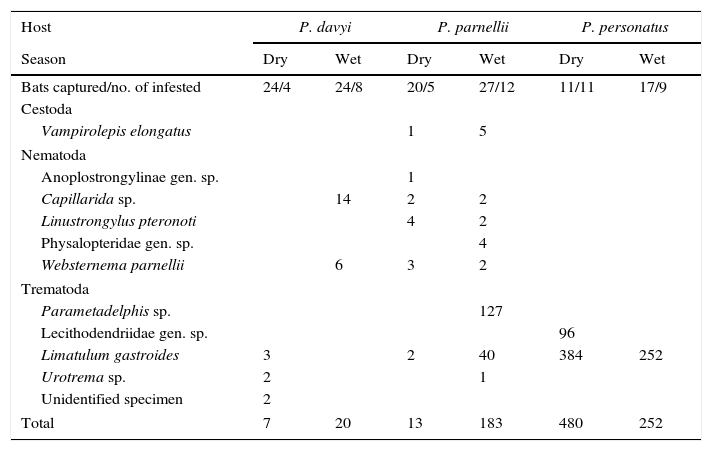
ResultsOne hundred and twenty three bats were collected during 7 fieldtrips, 40% of which had at least 1 helminth species. We found 10 species of parasites, totaling 955 individual helminths, of which 50% belonged to 5 nematode species, followed by 4 trematode species and 1 cestode species (40% and 10% of individuals, respectively). Only 6 of these species were collected in both seasons. Pteronotus personatus had the highest number of helminth individuals from the total of helminths. Two individual helminths collected in P. davyi were in too poor conditions to be identified to species and were not included in measurements of richness and diversity (Table 1). Limatulum gastroides was the only species recorded in the 3 insectivorous bat species and it was particularly abundant in P. personatus (Table 1).
Frequencies of helminths in 3 species of bats during the dry and wet seasons.
| Host | P. davyi | P. parnellii | P. personatus | |||
|---|---|---|---|---|---|---|
| Season | Dry | Wet | Dry | Wet | Dry | Wet |
| Bats captured/no. of infested | 24/4 | 24/8 | 20/5 | 27/12 | 11/11 | 17/9 |
| Cestoda | ||||||
| Vampirolepis elongatus | 1 | 5 | ||||
| Nematoda | ||||||
| Anoplostrongylinae gen. sp. | 1 | |||||
| Capillarida sp. | 14 | 2 | 2 | |||
| Linustrongylus pteronoti | 4 | 2 | ||||
| Physalopteridae gen. sp. | 4 | |||||
| Websternema parnellii | 6 | 3 | 2 | |||
| Trematoda | ||||||
| Parametadelphis sp. | 127 | |||||
| Lecithodendriidae gen. sp. | 96 | |||||
| Limatulum gastroides | 3 | 2 | 40 | 384 | 252 | |
| Urotrema sp. | 2 | 1 | ||||
| Unidentified specimen | 2 | |||||
| Total | 7 | 20 | 13 | 183 | 480 | 252 |
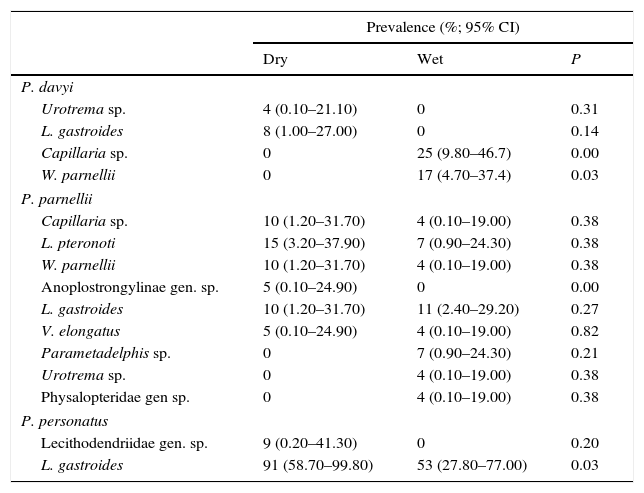
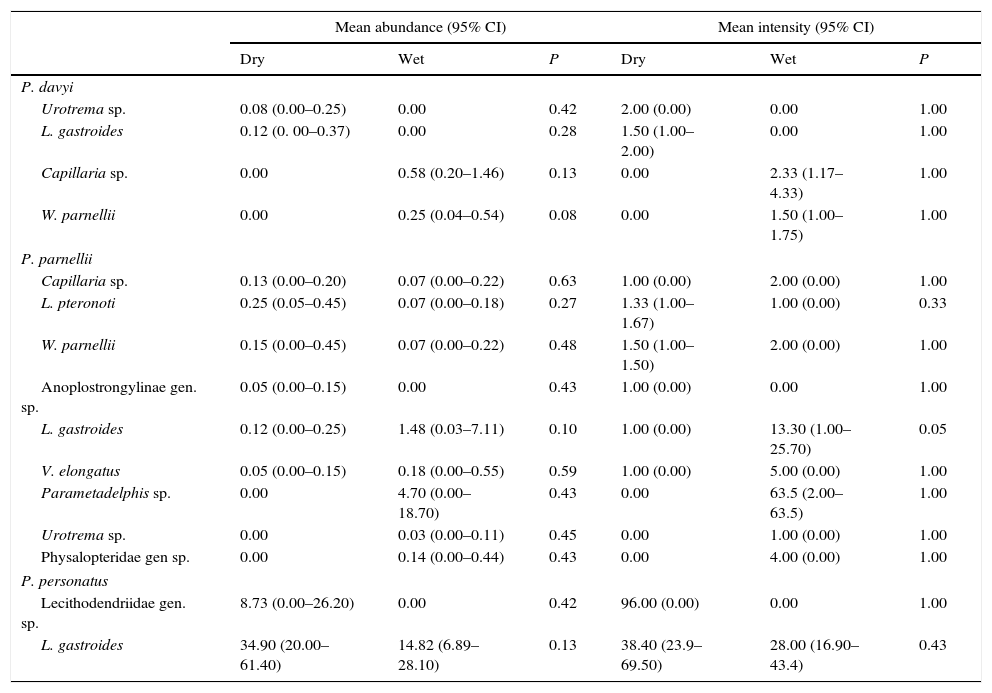
Prevalence was similar for both seasons in most endoparasite species except for Capillaria sp. and Websternema parnellii, which had a higher prevalence in the wet season in P. davyi, and Anoplostrongylinae gen. sp. and L. gastroides, in which prevalence was higher in the dry season in P. parnellii and P. personatus, respectively (Table 2). Mean abundance and intensity did not differ significantly between seasons for any endoparasite species (Table 3).
Prevalence registered in helminth parasites of Pteronotus species during the dry and wet seasons. P=values of chi-squared test with prevalence data and bootstrap test with the mean abundance and intensity data. Confidence intervals (CI) were set to 95% probability.
| Prevalence (%; 95% CI) | |||
|---|---|---|---|
| Dry | Wet | P | |
| P. davyi | |||
| Urotrema sp. | 4 (0.10–21.10) | 0 | 0.31 |
| L. gastroides | 8 (1.00–27.00) | 0 | 0.14 |
| Capillaria sp. | 0 | 25 (9.80–46.7) | 0.00 |
| W. parnellii | 0 | 17 (4.70–37.4) | 0.03 |
| P. parnellii | |||
| Capillaria sp. | 10 (1.20–31.70) | 4 (0.10–19.00) | 0.38 |
| L. pteronoti | 15 (3.20–37.90) | 7 (0.90–24.30) | 0.38 |
| W. parnellii | 10 (1.20–31.70) | 4 (0.10–19.00) | 0.38 |
| Anoplostrongylinae gen. sp. | 5 (0.10–24.90) | 0 | 0.00 |
| L. gastroides | 10 (1.20–31.70) | 11 (2.40–29.20) | 0.27 |
| V. elongatus | 5 (0.10–24.90) | 4 (0.10–19.00) | 0.82 |
| Parametadelphis sp. | 0 | 7 (0.90–24.30) | 0.21 |
| Urotrema sp. | 0 | 4 (0.10–19.00) | 0.38 |
| Physalopteridae gen sp. | 0 | 4 (0.10–19.00) | 0.38 |
| P. personatus | |||
| Lecithodendriidae gen. sp. | 9 (0.20–41.30) | 0 | 0.20 |
| L. gastroides | 91 (58.70–99.80) | 53 (27.80–77.00) | 0.03 |
Mean abundance and intensity registered in helminth parasites of Pteronotus species during the dry and wet seasons. P=values of chi-squared test with prevalence data and bootstrap test with the mean abundance and intensity data. Confidence intervals (CI) were set to 95% probability.
| Mean abundance (95% CI) | Mean intensity (95% CI) | |||||
|---|---|---|---|---|---|---|
| Dry | Wet | P | Dry | Wet | P | |
| P. davyi | ||||||
| Urotrema sp. | 0.08 (0.00–0.25) | 0.00 | 0.42 | 2.00 (0.00) | 0.00 | 1.00 |
| L. gastroides | 0.12 (0. 00–0.37) | 0.00 | 0.28 | 1.50 (1.00–2.00) | 0.00 | 1.00 |
| Capillaria sp. | 0.00 | 0.58 (0.20–1.46) | 0.13 | 0.00 | 2.33 (1.17–4.33) | 1.00 |
| W. parnellii | 0.00 | 0.25 (0.04–0.54) | 0.08 | 0.00 | 1.50 (1.00–1.75) | 1.00 |
| P. parnellii | ||||||
| Capillaria sp. | 0.13 (0.00–0.20) | 0.07 (0.00–0.22) | 0.63 | 1.00 (0.00) | 2.00 (0.00) | 1.00 |
| L. pteronoti | 0.25 (0.05–0.45) | 0.07 (0.00–0.18) | 0.27 | 1.33 (1.00–1.67) | 1.00 (0.00) | 0.33 |
| W. parnellii | 0.15 (0.00–0.45) | 0.07 (0.00–0.22) | 0.48 | 1.50 (1.00–1.50) | 2.00 (0.00) | 1.00 |
| Anoplostrongylinae gen. sp. | 0.05 (0.00–0.15) | 0.00 | 0.43 | 1.00 (0.00) | 0.00 | 1.00 |
| L. gastroides | 0.12 (0.00–0.25) | 1.48 (0.03–7.11) | 0.10 | 1.00 (0.00) | 13.30 (1.00–25.70) | 0.05 |
| V. elongatus | 0.05 (0.00–0.15) | 0.18 (0.00–0.55) | 0.59 | 1.00 (0.00) | 5.00 (0.00) | 1.00 |
| Parametadelphis sp. | 0.00 | 4.70 (0.00–18.70) | 0.43 | 0.00 | 63.5 (2.00–63.5) | 1.00 |
| Urotrema sp. | 0.00 | 0.03 (0.00–0.11) | 0.45 | 0.00 | 1.00 (0.00) | 1.00 |
| Physalopteridae gen sp. | 0.00 | 0.14 (0.00–0.44) | 0.43 | 0.00 | 4.00 (0.00) | 1.00 |
| P. personatus | ||||||
| Lecithodendriidae gen. sp. | 8.73 (0.00–26.20) | 0.00 | 0.42 | 96.00 (0.00) | 0.00 | 1.00 |
| L. gastroides | 34.90 (20.00–61.40) | 14.82 (6.89–28.10) | 0.13 | 38.40 (23.9–69.50) | 28.00 (16.90–43.4) | 0.43 |
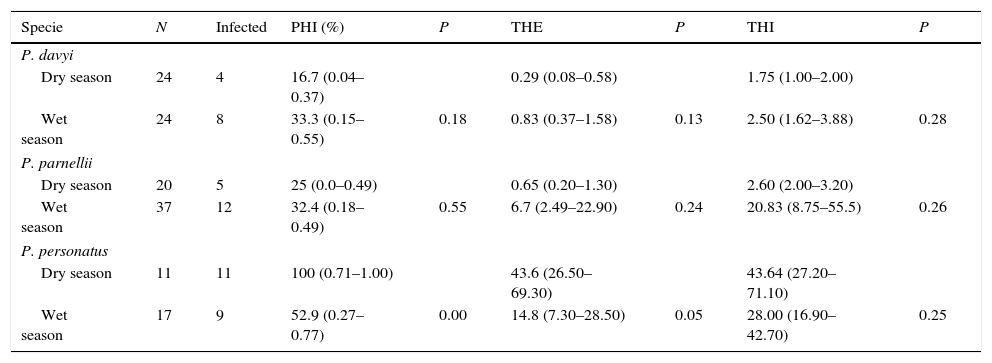
Percent of bats infected, average number of endoparasites per examined individual host and average number of endoparasites per infected individual host did not differ between seasons in most cases (Table 4). The only exception to this pattern was P. personatus, in which there was a higher proportion of infected bats in the dry season (Table 4).
Comparative of endoparasite load between the dry and wet season in 3 species of bats. N=number of bats collected; PHI=proportion of hosts infected considering the total endoparasite records; THE=average number of total endoparasites per host individual examined; THI=average number of total endoparasites per infected host individuals. P=values of chi-squared test with PHI data and bootstrap test with THE and THI data. Confidence intervals (CI) were set to 95% probability.
| Specie | N | Infected | PHI (%) | P | THE | P | THI | P |
|---|---|---|---|---|---|---|---|---|
| P. davyi | ||||||||
| Dry season | 24 | 4 | 16.7 (0.04–0.37) | 0.29 (0.08–0.58) | 1.75 (1.00–2.00) | |||
| Wet season | 24 | 8 | 33.3 (0.15–0.55) | 0.18 | 0.83 (0.37–1.58) | 0.13 | 2.50 (1.62–3.88) | 0.28 |
| P. parnellii | ||||||||
| Dry season | 20 | 5 | 25 (0.0–0.49) | 0.65 (0.20–1.30) | 2.60 (2.00–3.20) | |||
| Wet season | 37 | 12 | 32.4 (0.18–0.49) | 0.55 | 6.7 (2.49–22.90) | 0.24 | 20.83 (8.75–55.5) | 0.26 |
| P. personatus | ||||||||
| Dry season | 11 | 11 | 100 (0.71–1.00) | 43.6 (26.50–69.30) | 43.64 (27.20–71.10) | |||
| Wet season | 17 | 9 | 52.9 (0.27–0.77) | 0.00 | 14.8 (7.30–28.50) | 0.05 | 28.00 (16.90–42.70) | 0.25 |
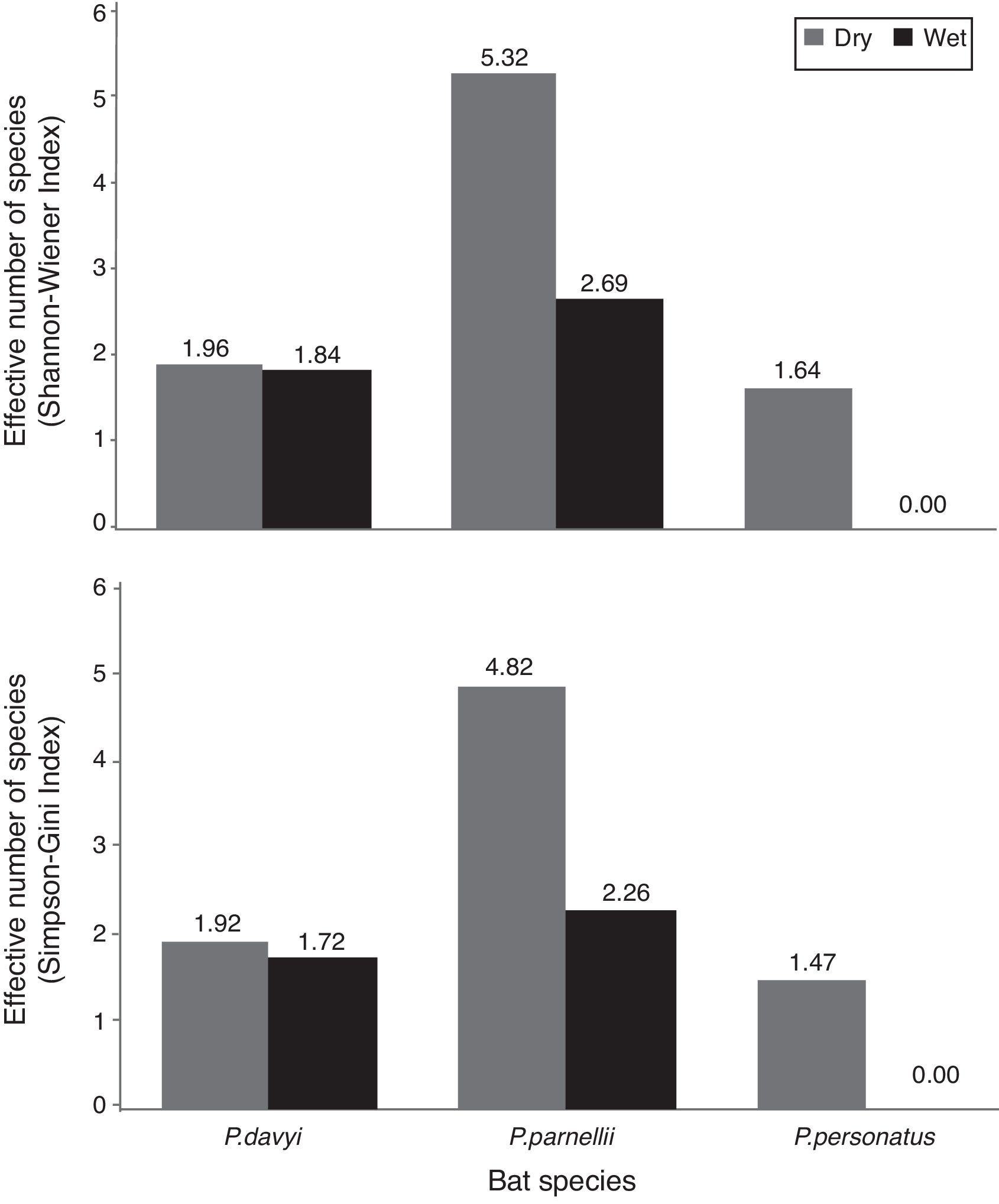
The highest helminth richness was recorded in P. parnellii in both seasons (Fig. 1). In terms of diversity, effective number of species calculated from Shannon–Wiener and Simpson–Gini Indexes were greater for all Pteronotus species during the dry season (Fig. 2).

 during the dry and wet seasons in 3 species of insectivorous bats (Pteronotus).")
We characterized the endoparasite load of 3 insectivorous bat species to examine the existence of seasonal changes in response to known seasonal ambient and prey fluctuations. Below, we discuss how our findings fitted to our predictions.
Our hypothesis of seasonal changes in parasite load was rejected in most cases either when comparing helminth species or the total endoparasite species. Mean abundance and intensity did not differ significantly between seasons for any endoparasite species. Our prediction of higher parasite load in the wet season was supported by the finding of higher prevalence in this season in Capillaria sp. and W. parnellii in P. davyi. In contrast, prevalence in Anoplostrongylinae gen. sp. and L. gastroides was higher in the dry season in P. parnellii and P. personatus, respectively. Percent of bats infected, average number of endoparasites per examined host individual and average number of endoparasites per infected host individual did not differ between seasons in most cases. The only exception to this pattern was P. personatus in which there was a higher proportion of infected bats in the dry season in contrast to our prediction.
Epidemiological models predict a positive relationship between host population density and abundance of macroparasite populations (Altizer, Harvell, & Friedle, 2003; Anderson & May, 1978, 1991; May & Anderson, 1978). When the host density is high, the transmission stages (e.g., eggs, larvae) of the parasite increase their probability of finding a permanent or intermediate host (Anderson & May, 1978; Arneberg, Skorping, Grenfell, & Read, 1998; Lafferty, 1997; May & Anderson, 1978). Our findings partly coincide with the increase in helminth abundance found in temperate insectivorous bats when abundance of intermediate hosts is presumably higher (Blankespoor & Ulmer, 1970; Lord et al., 2012; Nickel & Hansen, 1967), and with a study in which no seasonal patterns in helminth abundance were found in tropical insectivorous bats (Clarke, 2008). The lack of a uniform pattern in the cases when seasonal changes of parasite load were detected suggests that seasonal changes in the abundance of arthropod species that serve as intermediate hosts also might not be uniform. We based our prediction of a higher infestation rate in the wet season on the large increase in arthropod abundance during this season in the study region (Andresen, 2005; Güizado & Casas-Andreu, 2011). The variation in diet of Pteronotus species in Chamela is driven by the abundance and availability of insect prey (Salinas-Ramos et al., 2015), which are highly variable in time and space (Aldridge & Rautenbach, 1987; Whitaker, 1994). More than 1,423 species of arthropods have been recorded in Chamela, of which 570 species are found throughout the year while 622 and 231 species are found only during the wet or dry season, respectively (Pescador-Rubio et al., 2002). Seasonal changes in the abundance of some species of helminths detected in our study might reflect changes in the abundance of some arthropod species. For example, Trichoptera collected in rivers are more abundant in the dry season (Pescador-Rubio et al., 2002). Further studies focused on the examination of seasonal abundance of arthropod species and their endoparasites are warranted to understand seasonal dynamics of infection rate in Pteronotus species. An alternative explanation to the finparding of higher rates of infection when the abundance of intermediate hosts is lower is the “dilution effect” (Hamilton, 1971). The increase in the number of individuals of intermediate hosts leads to a decrease in an individual's probability to be parasitized (Krasnov, Stanko, & Morand, 2007; Ostfeld & Keesing, 2000). In other words, the probability of bats being infected by helminths decreases when the abundance of the intermediate host increases.
Our hypothesis of seasonal changes in endoparasite diversity was partly supported. Species richness was slightly higher in P. parnellii in the wet season, but it remained the same in P. davyi and P. personatus. However, following our prediction, the effective number of species was higher during the dry season for the 3 Pteronotus species. Dietary diversity of P. davyi and P. personatus increases in the dry season (Salinas-Ramos et al., 2015), when arthropod abundance is lower in the area (Andresen, 2005; Güizado & Casas-Andreu, 2011). It has been suggested that these bat species adopt a more generalist strategy when prey are limited and they become more selective when the insect abundance increases during the wet season (Salinas-Ramos et al., 2015). In the case of P. parnellii, it had the highest endoparasite richness and effective number of species of all insectivorous bats, which matches the more diverse diet reported in Chamela for P. parnellii compared to the diets of P. davyi and P. personatus all year long (Salinas-Ramos et al., 2015).
Host–parasite interactions could impact food webs and community structures (Mouritsen & Poulin, 2002; Sukhdeo, 2010) and the studies of the factors regulating helminth communities are complex (Lord et al., 2012). Our study examined the relationship between diversity and load of helminths and seasonal patterns of bat and abundance of potential intermediate hosts previously measured. We found heterogeneous patterns of this relationship that warrant further examination of seasonal abundance of intermediate hosts used as food by our focal bat species and of their helminth infection rates. These kinds of studies might be facilitated by the recent development of DNA libraries of arthropods and helminths in the study region (Fernández-Flores, Fernández-Triana, Martínez, & Zaldívar-Riverón, 2013; Prosser, Velarde-Aguilar, León-Règagnon, & Hebert, 2013).
This work was supported by grants given by Consejo Nacional de Ciencia y Tecnología (Conacyt, Red temática del código de barras de la vida, 2013–2015) to VLR; by Dirección General de Asuntos del Personal Académico (IN202113) to LGHM. VBSR thanks Programa de Doctorado en Ciencias Biológicas, Universidad Nacional Autónoma de México and Conacyt for the scholarship received. We also thank Carlos A. González-Castro, Andrea Rebollo-Hernández and Alejandro Zaldívar-Riverón for their assistance in the field and the staff at the Estación de Biología Chamela, UNAM for hosting this work.
Peer Review under the responsibility of Universidad Nacional Autónoma de México.