



Graphis robertusii Bárcenas-Peña, Herrera-Campos y Miranda es descrita por primera vez para la selva alta perennifolia de Los Tuxtlas, Veracruz. Es una especie conspicua caracterizada por lirelas erumpentes con margen talino lateral, labios no estriados blancos pruinosos, excípulo completamente carbonizado, himenio insperso, esporas transversalmente septadas y ácidos estíctico, salazínico y conestíctico como metabolitos secundarios. G. robertusii fue encontrada en corteza del dosel en diversos sitios de la Reserva de la Biosfera de Los Tuxtlas, Veracruz y es una de las especies de líquenes costrosos corticícolas con mayor abundancia en la región.
Graphis robertusii (G. robertusii) Bárcenas-Peña, Herrera-Campos and Miranda is described for the first time from the tropical rain forest in Los Tuxtlas, Veracruz. It is a conspicuous species characterized by erumpent lirella with lateral thalline margin separated from the excipulum, labia entire and white pruinose, completely carbonized excipulum, inspersed hymenium, transversally septate spores, and stictic, salazinic and constictic acids as secondary metabolites. G. robertusii was found on cortex at the canopy of several sites within Los Tuxtlas Biosphere Reserve, Veracruz and it is one of the most abundant corticolous crustose lichen species in the region.
La selva alta perennifolia es la más rica y compleja de todas las comunidades vegetales, ubicada en las zonas intertropicales del mundo y cuyo límite boreal de distribución en el continente americano se encuentra en México (Rzedowski, 1978). Hasta hace más de un siglo este tipo de vegetación estaba ampliamente distribuido desde el sur de San Luis Potosí, Veracruz, Hidalgo, Puebla y Oaxaca, norte y noreste de Chiapas, Campeche, Quintana Roo y Yucatán. Sin embargo, debido a su acelerada destrucción, en la actualidad se limita a pequeñas áreas al sureste en la península de Yucatán, la selva Lacandona en el noreste de Chiapas y en la selva El Ocote en la región limítrofe de Chiapas, Oaxaca y Veracruz (Guevara, Laborde y Sánchez-Ríos, 2004; Rzedowski, 1978). En la vertiente del golfo, la sierra de Los Tuxtlas en Veracruz es uno de los últimos reductos de esta vegetación, donde la flora y fauna aún son ricas (Ibarra-Manríquez, Martínez-Ramos, Dirzo y Núñez-Farfán, 1997), zona considerada de gran importancia biogeográfica debido a la presencia de taxa de afinidad austral, boreal y endémicos (Andrle, 1964; Dirzo y Miranda, 1991). Esta región es uno de los sitios neotropicales más estudiados en Latinoamérica y uno de los mejor conocidos en México (Dirzo, González-Soriano y Vogt, 1997; Guevara et al., 2004); entre los principales trabajos sobre diversos aspectos de biología tropical, se cuentan los de Álvarez y Naranjo (2003); Andrle (1964); Dirzo (1991); Gómez-Pompa y Del Amo (1985); Gómez-Pompa, Del Amo, Vázquez-Yáñez y Butanda (1976); González-Soriano, Dirzo y Vogt (1997) y Guevara et al. (2004). Sin embargo, es evidente la carencia de información biológica acerca de distintos procesos ecológicos y de ciertos componentes de la biota (Dirzo et al., 1997). En particular, el estudio de la biota liquénica de esta región, iniciado recientemente, se ha enfocado en la diversidad, ecología y biogeografía de los líquenes foliícolas, los más abundantes y diversos en la selva alta perennifolia (Bárcenas-Peña, 2004, 2007; Herrera-Campos, Huhndorf y Lücking, 2005; Herrera-Campos, Lücking, Pérez, Campos, Martínez-Colín y Bárcenas-Peña, 2004; Herrera-Campos, Martínez-Colín, Bárcenas-Peña y Lücking, 2004; Herrera-Campos y Lücking, 2002). No obstante, el conocimiento sobre los líquenes costrosos corticícolas aún es limitado. Entre los géneros de este último grupo, Graphis es el que presenta la mayor riqueza de especies en los trópicos (Lücking, Archer y Aptroot, 2009; Lücking, Chaves, Sipman, Umaña y Aptroot, 2008), la mayor parte de estas se encuentran a elevaciones medias en vegetación con una ligera estación seca, como las selvas lluviosas de baja altitud, en micrositios semiexpuestos a expuestos (dosel bajo, márgenes del bosque y árboles aislados), así como en vegetación secundaria o parcialmente perturbada. Asimismo, un número menor de especies se puede encontrar en bosques húmedos de montaña, bosque tropical estacionalmente seco y matorral xerófilo (Lücking et al., 2008).
El género Graphis fue descrito por Adanson (1763), posteriormente se presentaron varias propuestas de delimitación sin llegar a concretarse. Müller-Argoviensis (1880, 1882) realizó la primera separación universalmente aceptada. Staiger (2002) definió el género con talo corticado blanco-grisáceo, ascomas lireladas con labios bien desarrollados y disco cerrado, excípulo de lateral a completamente carbonizado, himenio de no insperso a insperso, ascosporas hialinas amiloides I+azul-violeta, transversalmente septadas a muriformes. Con esta circunscripción el género incluía más de 300 especies en su mayoría previamente asignadas a Graphis o Graphina sensu Müller; aun cuando muchos taxones fueron transferidos a Acanthothecis, Anomomorpha, Carbacanthographis, Diorygma, Dyplolabia, Fissurina, Glyphis y Hemithecium (Lücking et al., 2009). En los últimos años, Graphis tuvo cambios taxonómicos importantes, Lücking et al. (2008, 2009) publicaron claves para Graphis de Costa Rica y el mundo. Rivas-Plata et al. (2011) a través de filogenia molecular, así como de diferencias morfológicas, principalmente sobre la carbonización del excípulo e inspersión, encontraron que Graphis sensu Staiger representa 2 linajes separados: el primero, Graphis s. str., incluye las especies de los grupos G. scripta y G. subserpentina y G. striatula sensu Lücking; y el segundo, compuesto por los grupos G. acharii, G. dussii, G. marginata, G. nuda y G. symplecta, es incluido en el género Allographa Chevall. Recientemente, Bárcenas-Peña, Lücking, Miranda-González y Herrera-Campos (2014) actualizaron la clave mundial, incluyendo las especies descritas del 2009 al 2013 y reconociendo 330 especies a nivel mundial. En México se estiman 109 especies (Herrera-Campos et al., 2014), este número es aún bajo considerando la extensión y la amplia variedad de tipos de vegetación del país que faltan por explorar; por ejemplo, para Costa Rica, país más pequeño y menos diverso que México, se han documentado hasta la fecha 115 especies (Lücking et al., 2008). En este artículo se describe una nueva especie que se encuentra ampliamente distribuida en el dosel de la selva alta perennifolia de la región de Los Tuxtlas.
Materiales y métodosEl área de estudio se ubica en la selva alta perennifolia de la Reserva de la Biosfera Los Tuxtlas, Veracruz, en la sierra del mismo nombre en el golfo de México, entre las coordenadas 18°10′-18°45′ N, 94°42′-95°27′ O, en un rango altitudinal de 200 a 1,700 m (Ibarra-Manríquez et al., 1997; Miranda y Hernández, 1963). El clima de la región es clasificado como cálido A y semicálido A(C), con temperatura media anual de 22-26°C y precipitación media anual de 4,000mm (García, 1981; Soto y Gama, 1997).
Los ejemplares fueron recolectados a lo largo de 3 transectos establecidos para un estudio ecológico en 3 localidades de la reserva: en la Estación de Biología Tropical Los Tuxtlas: 1) cerro El Vigía; 2) en las inmediaciones de la laguna Zacatal y 3) ejido Adolfo López Mateos, selva del Marinero. La identificación de los ejemplares se realizó usando las claves de Bárcenas-Peña et al. (2014), de Lücking et al. (2009) y de Staiger (2002). Las observaciones morfológicas y anatómicas del talo y las esporas se realizaron bajo un estereoscopio Zeiss Stemi DV4 y un microscopio compuesto Zeiss Axiostar plus. Los cortes transversales de las lirelas se hicieron en un microtomo de congelación Leica CM1850, y se montaron en agua para su observación. Las esporas fueron medidas en agua y posteriormente teñidas con Lugol (0.3% IKI) para observar la reacción amiloide. Los metabolitos secundarios se identificaron con pruebas de tinción (KOH al 10%) en cortes delgados del talo y por cromatografía en capa fina (CCF) de acuerdo con la técnica estándar (Culberson y Ammann, 1979; Culberson y Johnson, 1982; Orange, James y White, 2001). Las fotografías del talo y las lirelas fueron tomadas en un microscopio estereoscópico multifocal Leica Z16 APO A con el programa Leica Application Suite y las fotos del corte del talo y la espora con una cámara Canon PowerShot G6 instalada en el microscopio compuesto antes mencionado.
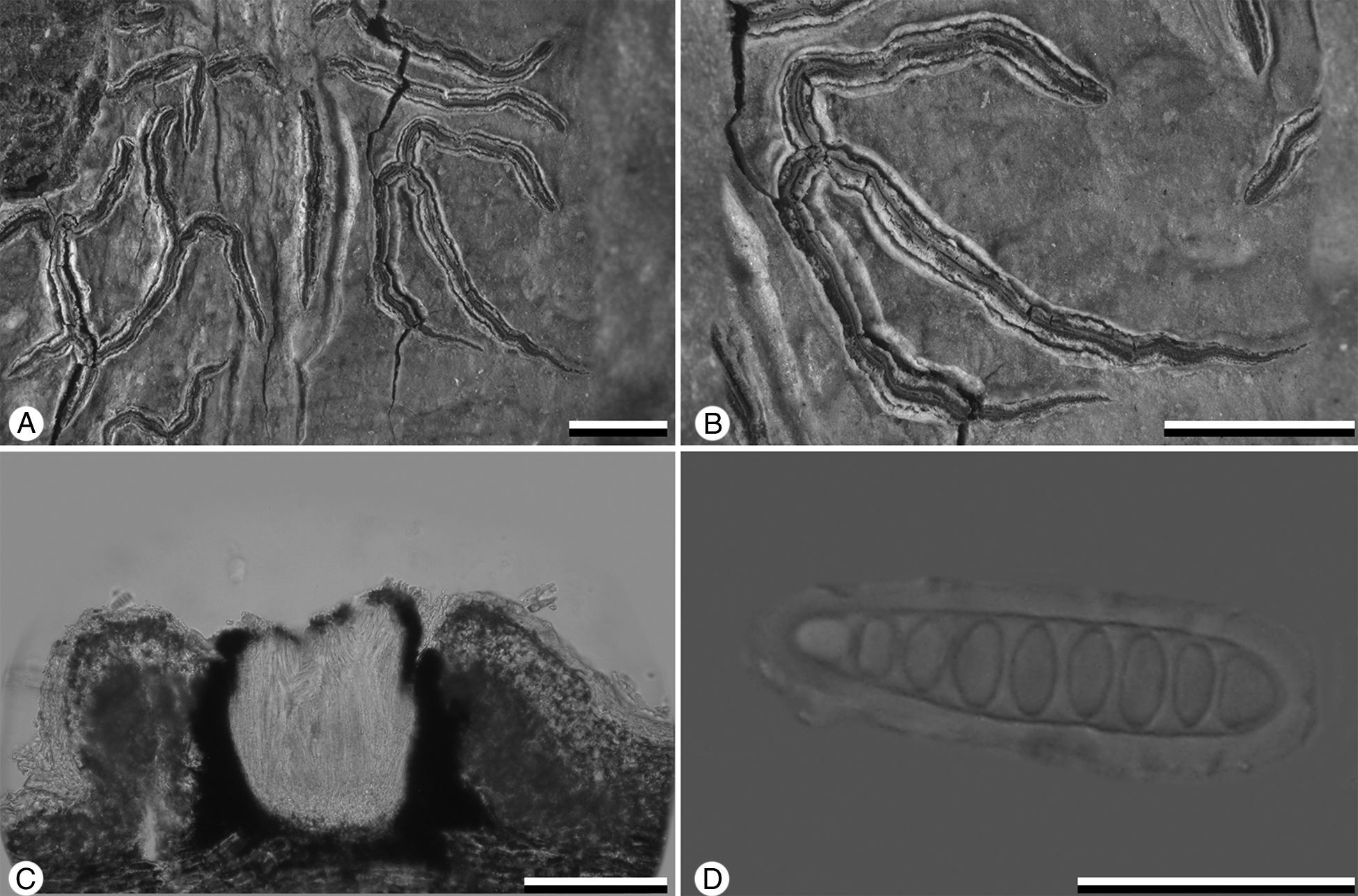
DescripciónGraphis robertusii Bárcenas-Peña, Herrera-Campos y R. Miranda n. sp. MycoBank No: MB 811600 (fig. 1A-D.TipoMéxico. Veracruz, Catemaco, Reserva de la Biosfera Los Tuxtlas, selva del Marinero, ejido Adolfo López Mateos, 18°26′1.4″ N, 94°57′56.8″ O, 300 m, selva alta perennifolia, sobre corteza de una Lauraceae no identificada, abril 2009, Bárcena-Peña 839 (holotipo, MEXU).
. A) Hábito esc=1mm. B) Lirelas erumpentes con disco cerrado y labios enteros con pruina esc=1mm. C) Corte tranversal de la lirela, excípulo completamente carbonizado, himenio insperso y margen doble esc=100μm; D, ascospora transversalmente septada esc=20μm.")
Graphis robertusii (holotype). A) Hábito esc=1mm. B) Lirelas erumpentes con disco cerrado y labios enteros con pruina esc=1mm. C) Corte tranversal de la lirela, excípulo completamente carbonizado, himenio insperso y margen doble esc=100μm; D, ascospora transversalmente septada esc=20μm.
Thallus corticate, smooth, pale greenish grey. Lirellae erumpent, partly branched, with laterally thalline margin separated from the excipulum (double margin), 3-7mm long, 0.1-0.2mm broad; labia entire, white pruinose. Excipulum completely carbonized. Hymenium inspersed. Ascospores 4-8 per ascus, fusiform, transversely 7-9-septate, 30-40×7-9μm, hyaline. Secondary chemistry: stictic, salazinic and constictic acids.
Talo corticícola, 1-3cm de diámetro, 50-100μm de grosor, continuo, superficie lisa, gris verdoso pálido, protalo ausente. Talo con corteza superior gruesa y cartilaginosa, capa algal irregular y cristales de oxalato de calcio. Apotecio lireliforme, flexuoso, parcialmente ramificado, erumpente, 3-7mm de largo, 0.1-0.2mm de ancho, 0.1-0.2mm de alto; disco cerrado, margen propio grueso; labios enteros, blancos, pruinosos, con margen talino lateral, del mismo color que el talo y separado del excípulo (margen doble); excípulo entero, completamente carbonizado, 50-70μm de ancho, negro, lateralmente cubierto por el talo y cristales de oxalato de calcio, parte basal interna café; hipotecio prosoplectenquimatoso, 20-30μm de alto, de incoloro a pálido amarillento; himenio 90-120μm de alto, incoloro, con inspersión tipo A; paráfisis hialinas, no ramificadas; epitecio granuloso, 10-20μm de alto, café; ascas fusiformes, 80-90×10-20μm; ascosporas 4-8 por asca, fusiformes, con 7-9 septos transversales, 30-40×7-9μm, 5-6 veces más largas que anchas, hialinas. Metabolitos secundarios (CCF) ácidos: estíctico, salazínico y conestíctico (accesorio). Sustrato corteza de ramas de dosel de una Lauraceae no identificada, Clarisia biflora, Pseudolmedia oxyphyllaria, Cojoba arborea, Croton schiedeanus y Pleuranthodendron lindenii.
Resumen taxonómicoGraphis robertusii posee una combinación de caracteres morfológicos: margen doble, pruina blanca en los labios, inspersión tipo A en el himenio, y químicos: ácidos estíctico, salazínico y conestíctico –accesorio– como metabolitos secundarios; que la ubican dentro del grupo de G. scripta. Esta especie únicamente ha sido recolectada en la región de Los Tuxtlas, en selva alta perennifolia, a elevaciones de 200-300 m. Es una especie corticícola distribuida ampliamente en el dosel medio a 30 m de alto, en las ramas de distintas especies de árboles.
Etimología. Esta especie es nombrada en honor al Dr. Robert Lücking, mentor y colaborador, por su constante apoyo y su importante contribución al desarrollo de la liquenología en México.
Paratipos. México. Veracruz, San Andrés Tuxtla, Estación de Biología Tropical Los Tuxtlas, cerca del tanque de agua, 18°35′08.6″ N, 95°04′43.1″ O, 228 m, selva alta perennifolia, sobre corteza de Pseudolmedia oxyphyllaria, enero 2009, Bárcenas-Peña 1432; cerro El Vigía, cerca de la vereda 5, 18°35′08.6″ N, 95°04′46.5″ O, 242 m, sobre corteza de Clarisia biflora, enero 2009, Bárcenas-Peña 1389; alrededores de la laguna Zacatal, 18°35′04.7″ N, 95°05′18.9″ O, 252 m, sobre corteza de Pleuranthodendron lindenii y Pseudolmedia oxyphyllaria, mayo 2011, Bárcenas-Peña 1250, 1251, respectivamente; alrededores de la laguna Zacatal, 18°35′03.8″ N, 95°05′17.2″ O, 257 m, sobre corteza de Croton schiedeanus, mayo 2011, Bárcenas-Peña 2079; alrededores de la laguna Zacatal, 18°35′03.7″ N, 95°05′16.1″ O, 263 m, sobre corteza de Cojoba arborea, mayo 2011, Bárcenas-Peña 2080; Catemaco, selva del Marinero, ejido Adolfo López Mateos, 18°26′1.4″ N, 94°57′56.8″ O, 300 m, sobre corteza de una Lauraceae no identificada, abril 2009, Bárcenas-Peña 844 (todos en MEXU).
Comentarios taxonómicosLa combinación de caracteres morfológicos, anatómicos, así como su hábitat ubican a G. robertusii dentro del grupo de G. scripta, cuyas especies se desarrollan en sitios semiexpuestos a baja altitud entre 0-1,000m (Lücking et al., 2008), condiciones ecológicas que concuerdan con aquellas de los sitios donde la nueva especie fue recolectada. Graphis robertusii se caracteriza químicamente por la presencia de ácidos estíctico, salazínico y en ocasiones ácido conestíctico, lo que la convierte en la primera especie con esta combinación química dentro del grupo G. scripta. Actualmente, este grupo está conformado por 42 especies, sin embargo, Lücking (2012) estima que podría alcanzar las 90 especies.
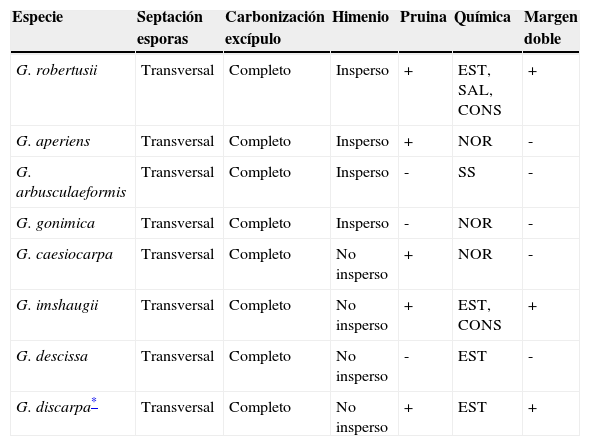
Las especies del grupo mencionado más similares a G. robertusii son: G. aperiens, G. arbusculaeformis, G. descissa, G. discarpa, G. gonimica y G. imshaugii. Estas especies poseen esporas transversalmente septadas y excípulo completamente carbonizado, de la misma manera que la especie aquí descrita, sin embargo, se diferencian de esta en al menos 2 caracteres. El himenio de G. robertusii es insperso, mientras que en G. descissa, G. caesiocarpa y G. imshaugii es no insperso. Los labios de G. arbusculaeformis, G. descissa y G. gonimica carecen de pruina contrastando con los labios de G. robertusii. Por otro lado, el margen talino lateral separado del excípulo (margen doble) es una característica especial que solo presentan 2 especies del grupo: G. imshaugii y G. robertusii (tabla 1).
Comparación de caracteres de G. robertusii, G. discarpa y las especies similares del grupo G. scripta, de acuerdo con la nomenclatura de Lücking et al. (2009).
| Especie | Septación esporas | Carbonización excípulo | Himenio | Pruina | Química | Margen doble |
|---|---|---|---|---|---|---|
| G. robertusii | Transversal | Completo | Insperso | + | EST, SAL, CONS | + |
| G. aperiens | Transversal | Completo | Insperso | + | NOR | - |
| G. arbusculaeformis | Transversal | Completo | Insperso | - | SS | - |
| G. gonimica | Transversal | Completo | Insperso | - | NOR | - |
| G. caesiocarpa | Transversal | Completo | No insperso | + | NOR | - |
| G. imshaugii | Transversal | Completo | No insperso | + | EST, CONS | + |
| G. descissa | Transversal | Completo | No insperso | - | EST | - |
| G. discarpa* | Transversal | Completo | No insperso | + | EST | + |
Fuente: Lücking, 2012.
CONS:conestíctico; EST:estíctico; NOR:norestíctico; SAL:salazínico; SS:sin sustancias.
Las especies comparadas tienen 5 combinaciones químicas distintas, siendo G. robertusii la que presenta un mayor número de metabolitos. Graphis imshaugii tiene los ácidos estíctico y conestíctico; G. aperiens, G. caesiocarpa y G. gonimica solo presentan ácido norestíctico, mientras que en G. descissa el metabolito secundario principal es el ácido estíctico; G. arbusculaeformis carece de metabolitos secundarios (tabla 1). Finalmente, G. discarpa no pertenece al grupo G. scripta (Lücking, 2012), no obstante, es parecida a G. robertusii, entre otras cosas, por la presencia de lirelas con margen doble, mientras que los caracteres que las separan son el himenio, no insperso en G. discarpa e insperso G. robertusii y la diferente combinación química (tabla 1).
| Clave de identificación para las especies del grupo G. scripta más similares a G. robertusii. Se incluye G. discarpa, que no pertenece al grupo mencionado. |
| 1 Himenio no insperso........................................................................................................2 |
| Himenio insperso….........................................................................................................5 |
| 2(1) Lirelas erumpentes, alargadas, irregularmente ramificadas, margen talino lateral, disco cerrado, labios sin pruina, ácido estíctico..…...............................G. descissa Müll Arg. |
| Lirelas inmersas a erumpentes, alargadas, ramificadas a raramente ramificadas, margen talino lateral, disco cerrado, labios con pruina y ácido estíctico, conestíctico o norestíctico………………………...….........…………………………………………..3 |
| 3(2) Lirelas inmersas a erumpentes, alargadas, ramificadas a raramente ramificadas, margen talino lateral separado del excípulo (doble), ácido estíctico y/o conestíctico (K+amarillo)......………………………………………………………………………4 |
| Lirelas erumpentes, alargadas, ramificadas, margen talino lateral a completo delgado, ácido norestíctico (K+amarillo formando cristales rojos)…..G. caesiocarpa Redinger. |
| 4(3) Lirelas inmersas, alargadas, raramente ramificadas, ácido estíctico….................................................................................................G. discarpa A. W. Archer. |
| Lirelas erumpentes, alargadas, ramificadas, ácidos estíctico y conestíctico..............….…………………………………………...G. imshaugii M. Wirth y Hale. |
| 5(1) Lirelas erumpentes, labios sin pruina y disco cerrado...…………………………….…6 |
| Lirelas erumpentes, labios con pruina y disco abierto o cerrado...…………...………..7 |
| 6 (5) Lirelas de cortas a alargadas, escasa a irregularmente ramificadas, margen talino lateral, ácido norestíctico………………….……...........................G. gonimica Zahlbr. |
| Lirelas muy largas y radialmente ramificadas, margen talino lateral, sin sustancias secundarias……….....……….….......................G. arbusculaeformis (Vain.) Lücking. |
| 7 (5) Lirelas cortas, escasamente ramificadas, margen talino lateral, disco abierto, ácido norestíctico.…..………..………………………………..............G. aperiens Müll Arg. |
| Lirelas alargadas, parcialmente ramificadas, margen talino lateral separado del excípulo (doble), disco cerrado, ácido estíctico, salazínico y conestíctico.… |
| ……………………..G. robertusii Bárcenas-Peña, Herrera-Campos y R. Miranda. |
A la Bióloga Rosamond-Coates, Jefa de la Estación de Biología Tropical Los Tuxtlas, por el apoyo logístico dentro de la reserva. Al Maestro en Ciencias Álvaro Campos por su ayuda en el campo y en la identificación de la especies de forofitos. A la Bióloga Susana Guzmán por su asistencia en el Laboratorio de Microscopía y Fotografía de la Biodiversidad IBUNAM. Al Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica-UNAM (proyecto N.°. IN225808). Al Instituto de Biología, UNAM por la infraestructura requerida para realizar el presente estudio. El primer autor agradece al Posgrado en Ciencias Biológicas-UNAM por la formación recibida durante sus estudios de doctorado y al Conacyt por la beca otorgada (Núm.165048).
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.