





Estuarine amphipods of the western Gulf of Mexico represent mostly eurythermal species. Sampling took place in Summer and Winter on seagrass beds, macroalgae and bare substrates, at 75 localities in Laguna Madre and 34 in Laguna de Tamiahua, Mexico, with a Renfro beam net and an otter trawl. A total of 19,398 specimens of 19 species were collected. Cymadusa compta was dominant in both lagoons with 63.4% of the total abundance. First records are 7 species for Laguna Madre and 11 species for Laguna de Tamiahua. A literature review updated the number of species from the 19 collected to 29. An analysis of the species’ distribution produced 3 zoogeographic patterns: Warm and Caribbean, Warm Temperate and Caribbean, and temperate. A hierarchical cluster analysis of the geographic distribution data of the 29 species produced 2 assemblages: one with an affinity for the warmer regions of the Gulf of Mexico, Caribbean Sea and Brazil, and another associated with the temperate conditions of the northern Gulf of Mexico, the eastern USA, Canada and Argentina, each group indicating a preference for either higher or lower water temperatures.
Los anfípodos estuarinos del golfo de México occidental representan en su mayoría especies euritermas. El muestreo se llevó a cabo en verano e invierno en pastos marinos, macroalgas y sustratos sin vegetación, en 75 localidades de laguna Madre y 34 de laguna de Tamiahua, México, con una red Renfro y una red camaronera. Se recolectaron 19,398 ejemplares de 19 especies. Cymadusa compta fue dominante en ambas lagunas con el 63.4% de la abundancia total. Se anotaron como primeros registros 7 especies en la laguna Madre y 11 en la laguna de Tamiahua. Se actualizó el número de especies de las 19 recolectadas a 29 mediante una revisión de la literatura. El análisis de la distribución de las especies proporcionó 3 patrones zoogeográficos: cálido-caribeño, cálido-templado-caribeño y templado. El análisis de conglomerados de la distribución geográfica de las 29 especies formó 2 grupos: uno con afinidad por las regiones más cálidas del golfo de México, Caribe y Brasil, y otro asociado a las condiciones templadas del norte del golfo de México, el este de EE. UU., Canadá y Argentina, indicando cada grupo una preferencia por temperaturas más cálidas o más frescas.
Amphipods, together with polychaetes and bivalves, make up 77% of the abundance in benthic communities of continental shelves and deep seas (Probert & Grove, 1998). They are a diverse, abundant and species-rich group of peracarid crustaceans. Most are free-living marine species, though some are freshwater, brackish or terrestrial. They are mostly eurythermal and euryhaline. They inhabit a wide variety of habitats and their geographic distribution is mostly cosmopolitan. They influence the structure of estuarine and coastal communities through their high abundance (Baldo, Arias, & Drake, 2001; Saunders, Attrill, Shaw, & Rowden, 2003; Tanner, 2006) and varied reproductive strategies and behaviour (Johnson, Stevens, & Watling, 2000; Thiel, 1997). They also affect community function by playing a part in the biogeochemical cycles of sedimentary organic matter (Gardner, Escobar, Cruz, & Rowe, 1993; Thiel & Hinojosa, 2009), promoting bioturbation and sediment stabilisation in soft substrates, as parasites of some vertebrates and vectors in the transmission of diseases and microorganisms, as well as through their feeding habits (DeBlois & Leggett, 1993; Duffy, 1990; Kamermans et al., 2002), nutrient recycling, linking foraging and detritivore food webs, and being prey of fish, birds and other crustaceans (Escobar & Soto, 1997; Hill & Elmgren, 1992; Zupo & Nelson, 1999). They are also excellent bioindicators of environmental quality (Montagna & Harper, 1996; Thomas, 1993b; Winfield & Ortiz, 2003).
Laguna Madre in Tamaulipas and Laguna de Tamiahua in Veracruz, Mexico, are large coastal lagoon systems that serve as nursery and feeding grounds for freshwater, estuarine and marine species. Their high biodiversity supports artisanal fisheries of oysters, shrimp, swimming crabs and mullet. Laguna Madre also provides feeding and refuge for migratory bird populations (Tunnell & Judd, 2002). The salinity gradients, submerged aquatic vegetation and different substrate types favour a variety of habitats that are available for resident and visiting species. In particular, the part that aquatic vegetation plays in the recruitment of many estuarine invertebrate and fish species has long been recognised (Heck & Crowder, 1991). High values of abundance have been recorded for species of invertebrates and fish that are temporal or permanent community components in these 2 lagoons (Raz-Guzmán & Barba, 2000). However, few studies have focused on the amphipods of these lagoons, and their zoogeography has not been reported. A literature review of studies on amphipods of the Mexican Gulf of Mexico coastal lagoons provided only the papers of Corona, Soto, and Sánchez (2000) and Cházaro, Winfield, Ortiz, and Álvarez (2002) for Laguna de Términos in Campeche, Winfield, Ortiz, Franco, and Bedia (1997), Winfield, Escobar, and Álvarez (2001), Winfield, Cházaro-Olvera, and Álvarez (2007), and Winfield and Ortiz (2011) for Laguna de Alvarado, Cházaro et al. (2002) and Winfield and Ortiz (2011) for Laguna Camaronera, and Winfield and Ortiz (2011) for Laguna de Sontecomapan in Veracruz, and Barba and Sánchez (2005) and Ortega (2013) for Laguna Madre in Tamaulipas. The only study found for Laguna de Tamiahua was that of Winfield and Ortiz (2011).
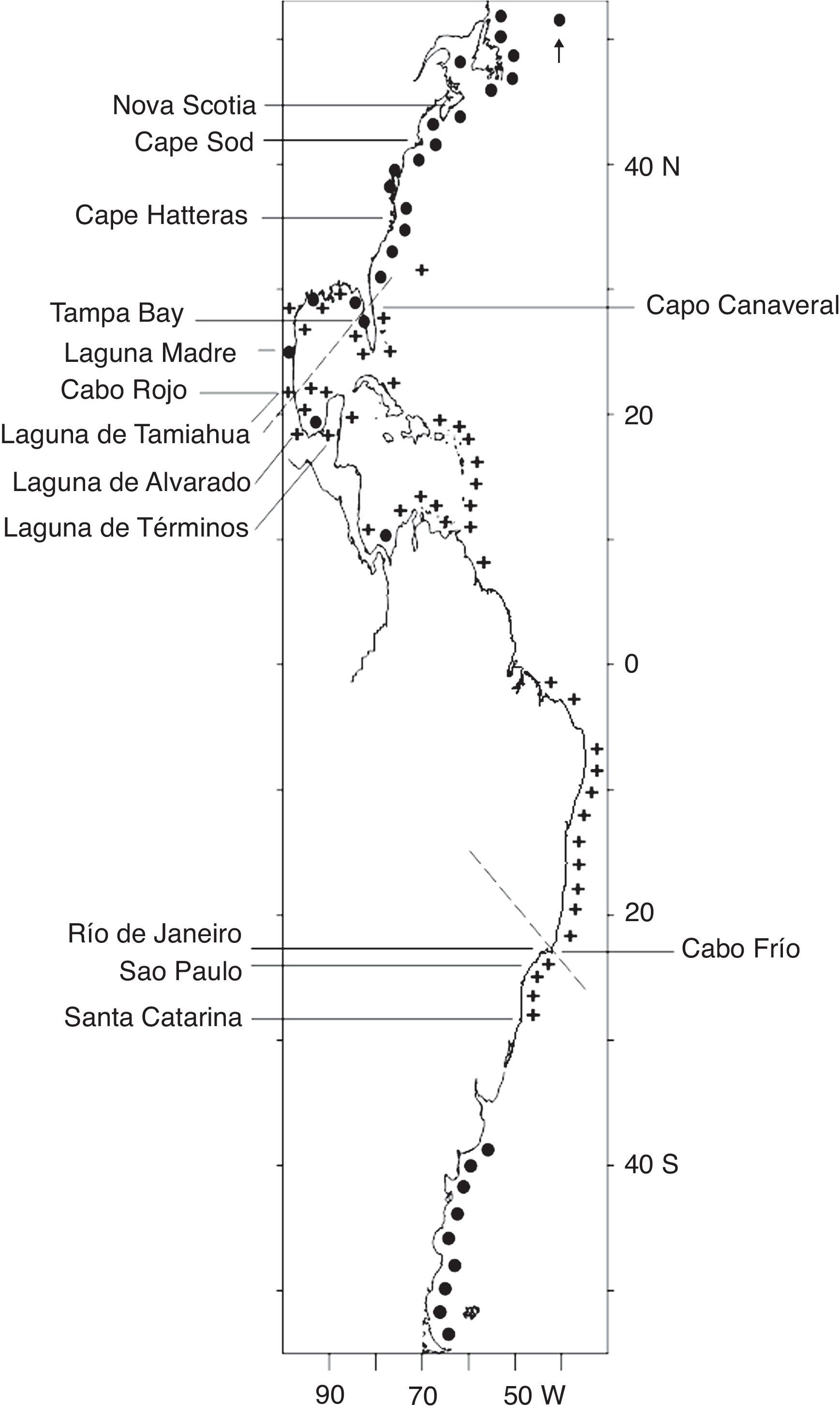
The zoogeographic provinces along the Western Atlantic and the Gulf of Mexico have been described by Hedgpeth (1953), Briggs (1974), and Barnwell and Thurman (1984). Zoogeographic province boundaries are important barriers to the dispersal of tropical shallow marine species (Thurman, 1987). The boundaries along the Western Atlantic include the northern tropical-temperate boundary along an imaginary line that spans from Cabo Rojo in Veracruz to Tampa Bay and Cape Canaveral in Florida and the southern tropical-temperate boundary at Cabo Frío in Brazil. Laguna de Tamiahua is zoogeographically significant as its island barrier of Cabo Rojo lies at the transition between the Caribbean-tropical and the Carolinean-Temperate provinces in the western Gulf of Mexico (Thurman, 1987).
The recording of species along a geographical gradient provides information on the biogeographical and systematic structure of life, and the mapping of the geographic distribution of species contributes to understanding ecological and evolutionary patterns and processes. Cluster analyses assemble species with similar geographic localities, as well as geographic localities with a similar species composition, and support the establishment of zoogeographic distribution patterns (Craw, Grehan, & Heads, 1999). Considering that amphipod studies in Mexican coastal lagoons are few, the purpose of this study was to define the zoogeographic patterns of 29 amphipod species recorded for Laguna Madre and Laguna de Tamiahua, based on updated data of the species’ geographic distribution that includes both published records for the Western Atlantic and the localities sampled in this study.
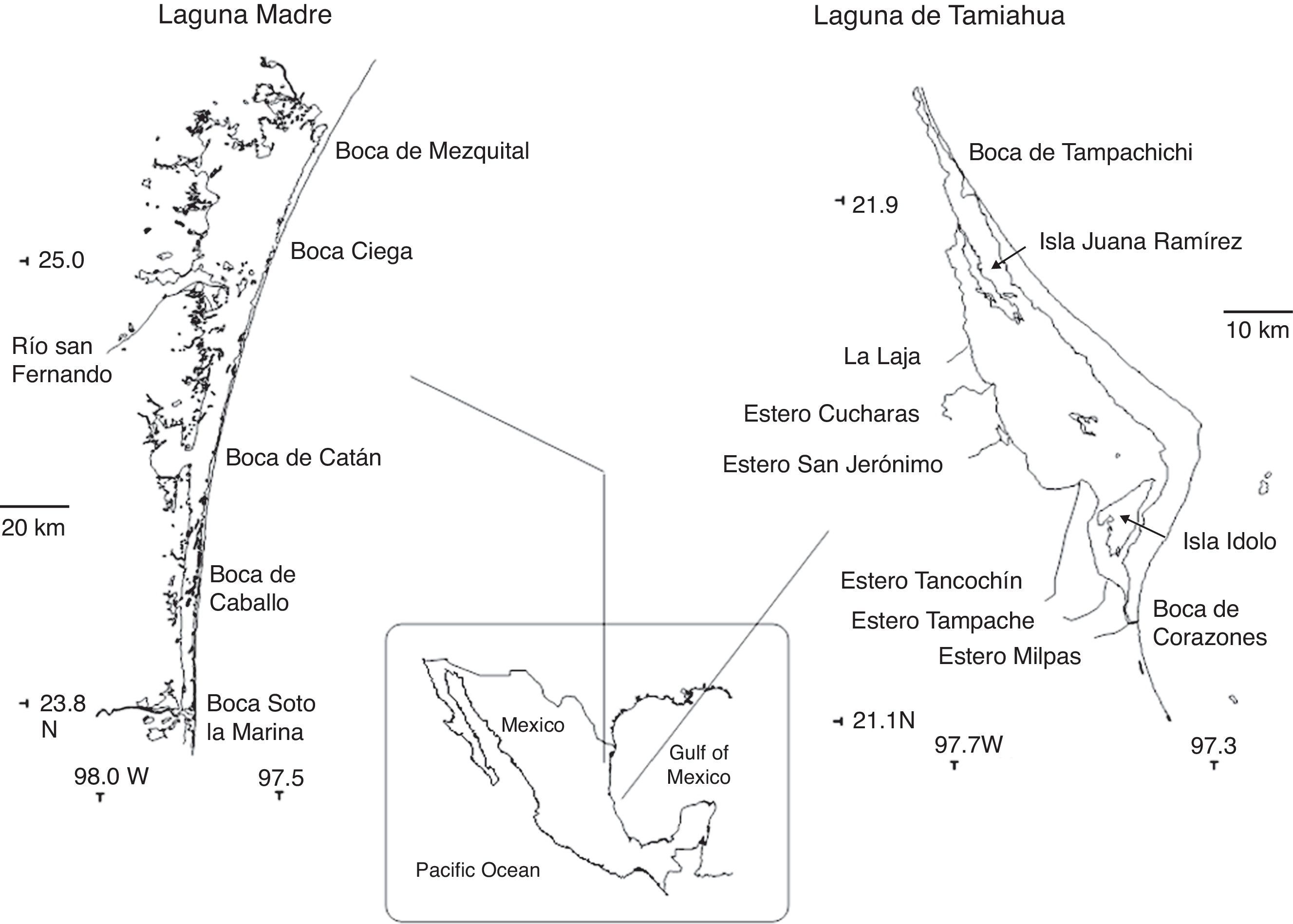
Materials and methodsLaguna Madre is the largest coastal lagoon in Mexico with an area of 200,000ha. It lies at 23°45′–25°27′ N, 97°23′–97°52′ W. It is limited to the north by the delta of the Río Bravo and to the south by the Río Soto la Marina. In 1996, the sand barrier was breached by 5 inlets: Boca de Mezquital, Boca Ciega, Boca de Catán, Boca de Caballo and Boca Soto la Marina. As a result of its location in a semi-arid area, its limited communication with the sea and the reduced input of freshwater from the Río San Fernando, it is a hypersaline system with salinities of 33–62‰, water temperatures of 25–35°C in Summer and 19–25°C in Winter, and a maximum depth of 6m (measured with a rope and dead weight). Further south, Laguna de Tamiahua is the third largest coastal lagoon in the Mexican Gulf of Mexico with 88,000ha. It lies at 21°16′–22°05′ N and 97°23′–97°43′ W. It is limited to the north by the Río Pánuco and to the south by the Río Tuxpan, with the sandy barrier of Cabo Rojo to the east and 2 inlets, Tampachichi to the north and Corazones to the south. Six streams discharge into the lagoon along its western banks and contribute freshwater seasonally. For this reason, salinity varies from 22 to 38‰, water temperatures from 28 to 35°C in Summer and 22 to 27°C in Winter, and the maximum depth is 4m (Fig. 1). Both lagoons have 2 marked climatic seasons, a dry Summer, and a rainy Winter when strong northerly winds (northers) are common (Salcedo, Soto, Estradas-Romero, & Botello, 2017).

The submerged aquatic vegetation in both lagoons is characterised by the shoalgrass Halodule wrightii Aschers that settles along the inner margins of the sand barriers, and macroalgae along the western margins of the lagoons that include the green algae Cladophora sp. and Halimeda incrassata (Ellis) Lamouroux syn. H. tridens, the brown algae Dictyota dichotoma (Hudson) Lamouroux, Padina sp. and Rosenvingea intricata (J. Agardh) Börgensen, and the red algae Hypnea cervicornis J. Agardh, Gracilaria blodgettii Harvey, Gingicithara cylindrica Börgensen, Chondria littoralis Harvey and Digenea simplex (Wulfen) C. Agardh. Fringing mangroves in Laguna Madre are present only in the extreme south and are mostly Avicennia germinans (L.)L., while in Laguna de Tamiahua one finds small mangrove forests of Rhizophora mangle L., A. germinans, Laguncularia racemosa (L.) L. F. Gaertner and Conocarpus erectus L.
Specimens were collected in Summer (August) and Winter (December) of 1996 from 75 localities in August and 32 in December in Laguna Madre, and 34 localities in August and 23 in December in Laguna de Tamiahua (Fig. 1). Regarding the sampling date, it is important to make 3 considerations after having consulted the only 3 more recent studies available: 1) that only 3 species of 13 collected by Barba and Sánchez (2005) and 3 species of 12 collected by Ortega (2013) in Laguna Madre, as well as 5 species of 6 collected by Winfield and Ortiz (2011) in Laguna de Tamiahua, were not collected in our study; 2) that in our study we recorded 9 more species than Barba and Sánchez (2005), 11 more species than Ortega (2013) and 11 more species than Winfield and Ortiz (2011), and 3) that all the species recorded in these 3 papers plus the species collected in our study are included here, in view of which our species list may be considered to be taxonomically updated and valid.
Water temperature and salinity were recorded with a field thermometer and a refractometer respectively. Two nets were used to obtain specimens of different sizes and habitats: a Renfro beam net (1mm mesh size, 50m2 sampling area) and an otter trawl (1cm mesh size, 1.5–3min capture per unit effort). The amphipods were preserved in 10% formalin in the field, after which they were transferred to a solution of glycerin and 70% alcohol for whitening previous to dissection.
The specimens were observed in detail in order to insure a correct identification of the species. The families are listed in alphabetical order. The assignment of genera to families follows Bousfield (1973), Martín and Díaz (2003) and LeCroy (2002, 2004, 2007). Exceptions are the Corophiidae that follow Myers and Lowry (2003) and the talitridan stat. nov. that follow Serejo (2004). The arrangement of genera and families included in the 2 latter works were adopted since they provide more recent phylogenetic evidence of relationships within the corophiidan and talitridan amphipods. The genera and species were identified following Barnard (1969), Bousfield (1973), Myers (1982), Myers and McGrath (1984), Ledoyer (1986), Conlan (1990), Barnard and Karaman (1991), Thomas (1993a), LeCroy (2000, 2002, 2004, 2007), Myers and Lowry (2003), Appadoo and Myers (2004), and Serejo (2004). The species list follows Martin and Davis (2001) for the Suborder Gammaridea, Myers and Lowry (2003) for the Suborder Corophiidea, with an updating following Ahyong et al. (2011) and Horton, Lowry, and De Broyer (2013).
An Olmstead-Tükey analysis was applied to the abundance and spatial frequency data in order to identify dominant species. These and first records of species and families are reported for each lagoon, as well as the species collected in this study together with 3 species reported by Barba and Sánchez (2005) for Laguna Madre, 3 species reported by Ortega (2013) for the same lagoon (his Protohyale sp. B was not included in the analysis), and 5 species reported by Winfield and Ortiz (2011) for Laguna de Tamiahua. An updating was carried out in January 2015 for all species’ names (http://www.itis.gov; Martin & Davis, 2001; Myers & Lowry, 2003) and geographic distributions. The specimens were kept in the Laboratorio de Ecología del Bentos, ICMyL, UNAM.
A map of the Western Atlantic including the important zoogeographic landmarks of Cape Cod, Cape Hatteras, Cape Canaveral and Tampa Bay in the United States, Cabo Rojo in Mexico, and Cabo Frío in Brazil, was prepared for each species considering the published data on the geographic distribution of each species (the geographic localities and the authors that reported them for each species are presented in Appendix 1), as well as the updated records for Laguna Madre and Laguna de Tamiahua. The Primer v.6 programme (Clarke & Gorley, 2006) was used to analyse a presence-absence matrix for species-geographic localities, and group geographic points according to Pearson's average linkage correlation to form clusters. A cluster analysis to group species provided no clear pattern.
ResultsLaguna Madre and Laguna de Tamiahua experience marked seasonal changes in water temperature throughout the year. In Winter, values drop to 19°C in Madre and to 22°C in Tamiahua, while in Summer, there is a maximum of 35°C in both lagoons. Salinity varied from 33 to 62‰ in Madre and from 22 to 38‰ in Tamiahua (Table 1).
Temperature, salinity, seasonal abundance (ind), number of species (spp.) and number of families (fam) of amphipods collected in each lagoon in this study.
Temperature Salinity | Laguna Madre 19–35°C 33–62‰ | Laguna de Tamiahua 22–35°C 22–38‰ |
|---|---|---|
| Abundance-species-families | 13,052 ind – 18 spp. – 14 fam | 6,346 ind – 12 spp. – 9 fam |
| Summer | 5,367 ind – 14 spp. | 4,209 ind – 10 spp. |
| Winter | 7,685 ind – 13 spp. | 2,137 ind – 11 spp. |
A total of 19,398 specimens of 19 amphipod species were collected in the 2 lagoons. Laguna Madre provided 13,052 amphipods of 18 species and 14 families, of which 5,367 specimens and 14 species were collected in August and 7,685 specimens and 13 species in December. In the case of Laguna de Tamiahua, the amphipods collected included 6,346 specimens of 12 species and 9 families, with 4,209 specimens and 10 species recorded in August, and 2,137 specimens and 11 species in December (Table 1). Three species of the 13 recorded by Barba and Sánchez (2005), 3 species of the 12 recorded by Ortega (2013), and 5 species of the 6 recorded by Winfield and Ortiz (2011) that were not collected in the present study were added to the list, providing a total of 24 species for Laguna Madre and 17 species for Laguna de Tamiahua. In total, 29 species were included in the zoogeographic analysis (Table 2).
Amphipod species and abundance in Laguna Madre (LM) and Laguna de Tamiahua (LT) (+ Barba & Sánchez, 2005; ++Winfield & Ortiz, 2011; +++Ortega, 2013).
| Family | Species | LM | LT |
|---|---|---|---|
| Subphylum Crustacea Brünnich, 1772 | |||
| Class Malacostraca Latreille, 1802 | |||
| Subclass Eumalacostraca Grobben, 1892 | |||
| Superorder Peracarida Calman, 1904 | |||
| Order Amphipoda Latreille, 1816 | |||
| Suborder Gammaridea Latreille, 1802 (sensu Martin & Davis, 2001) | |||
| Ampeliscidae Krøyer, 1842 | Ampelisca vadorumMills, 1963 | 2 | 2 |
| Ampelisca venetiensis Shoemaker, 1916 | ++ | ||
| Ampelisca verrilli Mills, 1967 | ++ | ||
| Amphilochidae Boeck, 1871 | Hourstonius laguna (McKinney, 1978) | 589 | 7 |
| Bateidae Stebbing, 1906 | Batea catharinensis F. Müller, 1865 | 794 | |
| Dexaminidae Leach, 1814 | Nototropis minikoi (A.O. Walker, 1905) | 119 | 2 |
| Gammaridae Leach, 1814 | Gammarus mucronatus Say, 1818 | 42 | 847 |
| Hyalidae Bulycheva, 1957 | Apohyale prevostii (H. Milne Edwards, 1830) | 5 | 791 |
| Parhyale hawaiensis (Dana, 1853) | +++ | ||
| Melitidae Bousfield, 1983 | Elasmopus levis (S.I. Smith, 1873) | 2684 | 80 |
| Elasmopus pectenicrus (Bate, 1862) | +++ | ||
| Elasmopus rapax Costa, 1853 | +++ | ||
| Melita nitida S.I. Smith, 1873 | 1 | 54 | |
| Phoxocephalidae G.O. Sars, 1891 | Eobrolgus spinosus (Holmes, 1905) | 1 | ++ |
| Talitridae Rafinesque, 1815 | Orchestia gammarellus (Pallas, 1766) | + | |
| Orchestia grillus (Bosc, 1802) | 13 | ||
| Suborder Corophiidea Leach, 1814 (sensu Myers & Lowry, 2003) | |||
| Infraorder Corophiida Leach, 1814 | |||
| Superfamily Aoroidea Stebbing, 1899 | |||
| Aoridae Stebbing, 1899 | Grandidierella bonnieroidesStephensen, 1948 | 55 | |
| Lembos websteri Bate, 1857 | + | ||
| Unciolidae Myers & Lowry, 2003 | Unciola serrata Shoemaker, 1945 | ++ | |
| Superfamily Corophioidea Leach, 1814 | |||
| Ampithoidae Stebbing, 1899 | Ampithoe longimana S.I. Smith, 1873 | 10 | 1 |
| Ampithoe valida S.I. Smith, 1873 | 4 | 76 | |
| Cymadusa compta (S.I. Smith, 1873) | 8054 | 4240 | |
| Corophiidae Leach, 1814 | Monocorophium acherusicum (Costa, 1851) | 1 | |
| Monocorophium tuberculatum (Shoemaker, 1934) | 202 | ||
| Infraorder Caprellida Leach, 1814 | |||
| Superfamily Microprotopoidea Myers & Lowry, 2003 | |||
| Microprotopidae Myers & Lowry, 2003 | Microprotopus raneyi Wigley, 1966 | 1 | |
| Superfamily Photoidea Boeck, 1871 | |||
| Ischyroceridae Stebbing, 1899 | Cerapus tubularis Say, 1817 | 1 | |
| Ericthonius brasiliensis (Dana, 1853) | 674 | 245 | |
| Jassa falcata (Montagu,1808) | + | ||
| Photidae Boeck 1871 | Photis longicaudata (Bate & Westwood, 1862) | ++ | |
The amphipod species of Laguna Madre and Laguna de Tamiahua, together with the anomuran and brachyuran crabs of the 4 largest estuarine systems of the western and southwestern Gulf of Mexico, contribute to the highly diversified fauna of the Tropical Western Atlantic (Raz-Guzmán & Sánchez, 1992a, 1992b, 1996, 1998; Guzmán, Sánchez, Soto, & Álvarez, 1986; Guzmán, Sánchez, & Soto, 1992; Raz-Guzmán, Sánchez, Peralta, & Florido, 2004). It is interesting to note that in spite of Cabo Rojo being the barrier island of Laguna de Tamiahua and representing a critical boundary for the northern dispersal of tropical shallow water fauna (Thurman, 1987), this lagoon and Laguna Madre further north share 12 of the 29 amphipod species recorded here, a 41% affinity that may indicate that these 2 lagoons constitute habitats of positive population growth from which amphipods can successfully disperse to neighbouring areas.
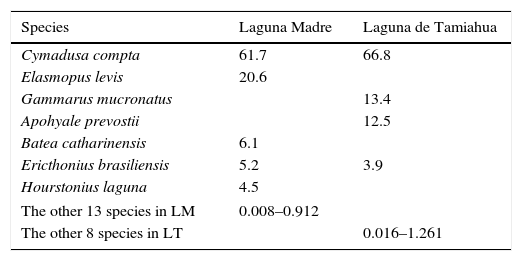
Cymadusa compta (S.I. Smith, 1873) was dominant in both lagoons with 63.4% of the overall total abundance. Of the 18 species collected in this study in Laguna Madre, 2 constitute 82.3% of the total abundance: C. compta (61.7%) and Elasmopus levis (S.I. Smith, 1873) (20.6%), and are followed by Batea catharinensis F. Müller, 1865 (6.1%), Ericthonius brasiliensis (Dana, 1853) (5.2%), Hourstonius laguna (McKinney, 1978) (4.5%), and the other 13 species with 0.008–0.912%. Of the 12 species collected in Laguna de Tamiahua, C. compta is markedly more abundant than the others with 66.8%, and is followed by Gammarus mucronatus Say, 1818 (13.4%), Apohyale prevostii (H. Milne Edwards, 1830) (12.5%), E. brasiliensis (3.9%), and the other 8 species with 0.016–1.261% (Table 3).
Dominant amphipod species in Laguna Madre (LM) and Laguna de Tamiahua (LT) (% of total abundance).
| Species | Laguna Madre | Laguna de Tamiahua |
|---|---|---|
| Cymadusa compta | 61.7 | 66.8 |
| Elasmopus levis | 20.6 | |
| Gammarus mucronatus | 13.4 | |
| Apohyale prevostii | 12.5 | |
| Batea catharinensis | 6.1 | |
| Ericthonius brasiliensis | 5.2 | 3.9 |
| Hourstonius laguna | 4.5 | |
| The other 13 species in LM | 0.008–0.912 | |
| The other 8 species in LT | 0.016–1.261 | |
Seven species reported here for Laguna Madre were not recorded by Barba and Sánchez (2005) or Ortega (2013). Ampelisca vadorumMills, 1963, H. laguna, Ampithoe longimana S.I. Smith, 1873, Ampithoe valida S.I. Smith, 1873, Melita nitida S.I. Smith, 1873, Eobrolgus spinosus (Holmes, 1905) and Orchestia grillus (Bosc, 1802) may thus be considered first records for this lagoon, as well as 2 families, the Amphilochidae and Phoxocephalidae. In the case of Laguna de Tamiahua, with the exception of G. mucronatus which was recorded previously by Winfield and Ortiz (2011), the other 11 species, A. vadorum, H. laguna, A. longimana, A. valida, C. compta, Monocorophium acherusicum (Costa, 1851), Nototropis minikoi (A.O. Walker, 1905), A. prevostii, E. brasiliensis, E. levis and M. nitida, are first records, together with the family Amphilochidae.
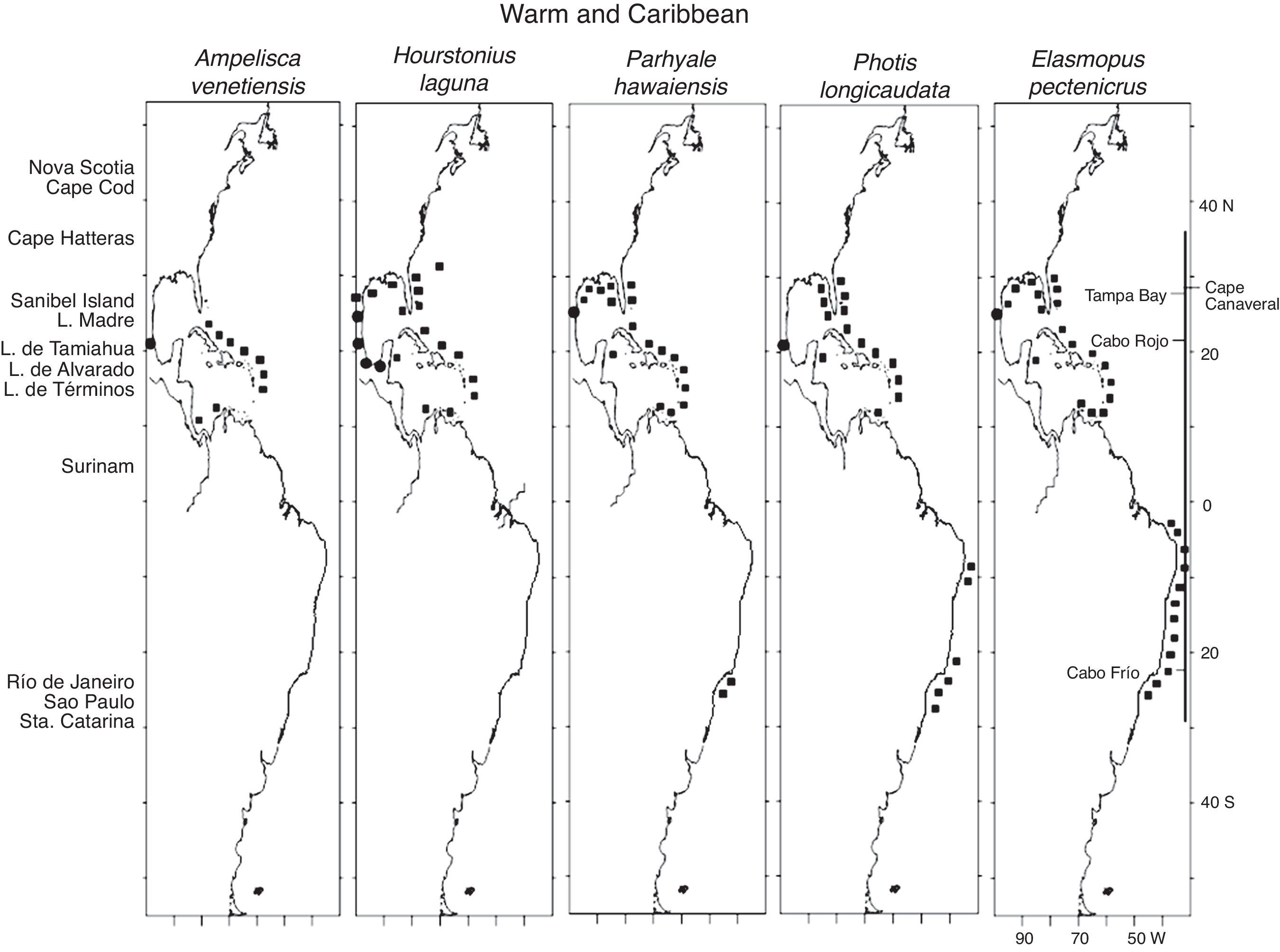
The mapping of the geographic distribution data of each of the 29 species recorded here made it possible to visually define 3 zoogeographic distribution patterns along the Western Atlantic from Newfoundland to Argentina: Warm and Caribbean with 5 species (17%), Warm Temperate and Caribbean with 18 species (62%) and Temperate with 6 species (21%). A hierarchical cluster analysis of the presence-absence matrix of geographic distribution data of the 29 amphipod species produced 2 groups at a correlation level of −0.05 (with other smaller groups at higher correlation levels). One group included the warmer regions of the Gulf of Mexico, Caribbean Sea and Brazil (+), and another group included the colder regions of the northern Gulf of Mexico, the eastern and northeastern United States, Canada, Greenland, and Argentina (●).
DiscussionThe first records reported here for Laguna Madre (A. vadorum, H. laguna, A. longimana, A. valida, M. nitida, E. spinosus and O. grillus) and Laguna de Tamiahua (A. vadorum, H. laguna, A. longimana, A. valida, C. compta, M. acherusicum, N. minikoi, A. prevostii, E. brasiliensis, E. levis and M. nitida) respond to the fact that previous studies on amphipods in the state of Veracruz have focused on Laguna Camaronera (Cházaro et al., 2002; Winfield & Ortiz, 2011), Laguna de Alvarado (Winfield & Ortiz, 2011; Winfield et al., 1997, 2001; Winfield, Cházaro-Olvera, et al., 2007), Laguna de Sontecomapan (Winfield & Ortiz, 2011), the Veracruz Reef System (Winfield & Ortiz, 2011; Winfield, Abarca-Arenas, & Cházaro-Olvera, 2007; Winfield, Cházaro-Olvera, Horta-Puga, Lozano-Aburto, & Arenas-Fuentes, 2010; Winfield, Cházaro-Olvera, Ortiz, & Palomo-Aguayo, 2011; Winfield, 2013) and the Veracruz continental shelf (Winfield & Ortiz, 2011), with only that of Winfield and Ortiz (2011) for Laguna de Tamiahua, while only those of Barba and Sánchez (2005) and Ortega (2013) have dealt with Laguna Madre in the state of Tamaulipas.
The 5 Warm and Caribbean species, Ampelisca venetiensis Shoemaker, 1916, H. laguna, Parhyale hawaiensis (Dana, 1853), Photis longicaudata (Bate & Westwood, 1862) and Elasmopus pectenicrus (Bate, 1862), are mostly distributed along the eastern coast of Florida and the Gulf of Mexico (except for A. venetiensis), throughout the Caribbean Sea, and along the coast of Brazil (except for A. venetiensis and H. laguna). These species include a few records along the Temperate northern Gulf of Mexico which may be explained as the result of dispersal by the shallow coastal currents that flow from Louisiana and Texas (the LATEX current) to Campeche during the Winter and back during the Summer (Fernández, Zavala, & Romero, 1998) (Fig. 2). The capacity to disperse is important in the life history of many species. Benthic invertebrates usually disperse through a swimming larval phase. However, many species with a direct development have wide geographic distributions (Castilla & Guikez, 2000; Thiel, 2003a). Floating and rafting have been identified as highly efficient dispersal mechanisms in these species (Highsmith, 1985; Thiel, 2003b), which include shallow-water isopods and amphipods (Locke & Corey, 1989), many of which brood their young. This has been supported by Havermans, De Broyer, Mallefet, and Zintzen (2007) who recorded how Jassa herdmani (Walker, 1893), a tube-building amphipod, may travel long distances between hard-substrate areas, and by records of brooding benthic peracarids found in plancton nets in areas far from their native habitats (Williams & Bynum, 1972). Most common floaters are small individuals that may remain suspended on the water surface for long periods of time. Small animals are also more likely to live on seagrass and macroalgae, and may become rafters when the plants break (Norton & Mathieson, 1983). However, gravid females have also been observed rendering rafting a potential dispersal mechanism. It is important to note that P. hawaiensis, a species used in developmental and genetic studies, may actually occur as a species complex (Myers, 1985).
 along the Western Atlantic (■) including the Mexican lagoons (●).")
The warm affinity of these species is supported by records from other warm localities that include Hawaii, the Philippines and the South China Sea for P. hawaiensis, and Hawaii, the Red Sea, Madagascar, Mauritius, India, Indonesia, the South China Sea, New Guinea and Australia for E. pectenicrus. P. longicaudata has been recorded in the Indian Ocean, however it is also present in colder localities including Sweden, Ireland, Scotland, England, Belgium, France, Spain, Greece, the Mediterranean Sea, the Black Sea and Japan (see Appendix 1).
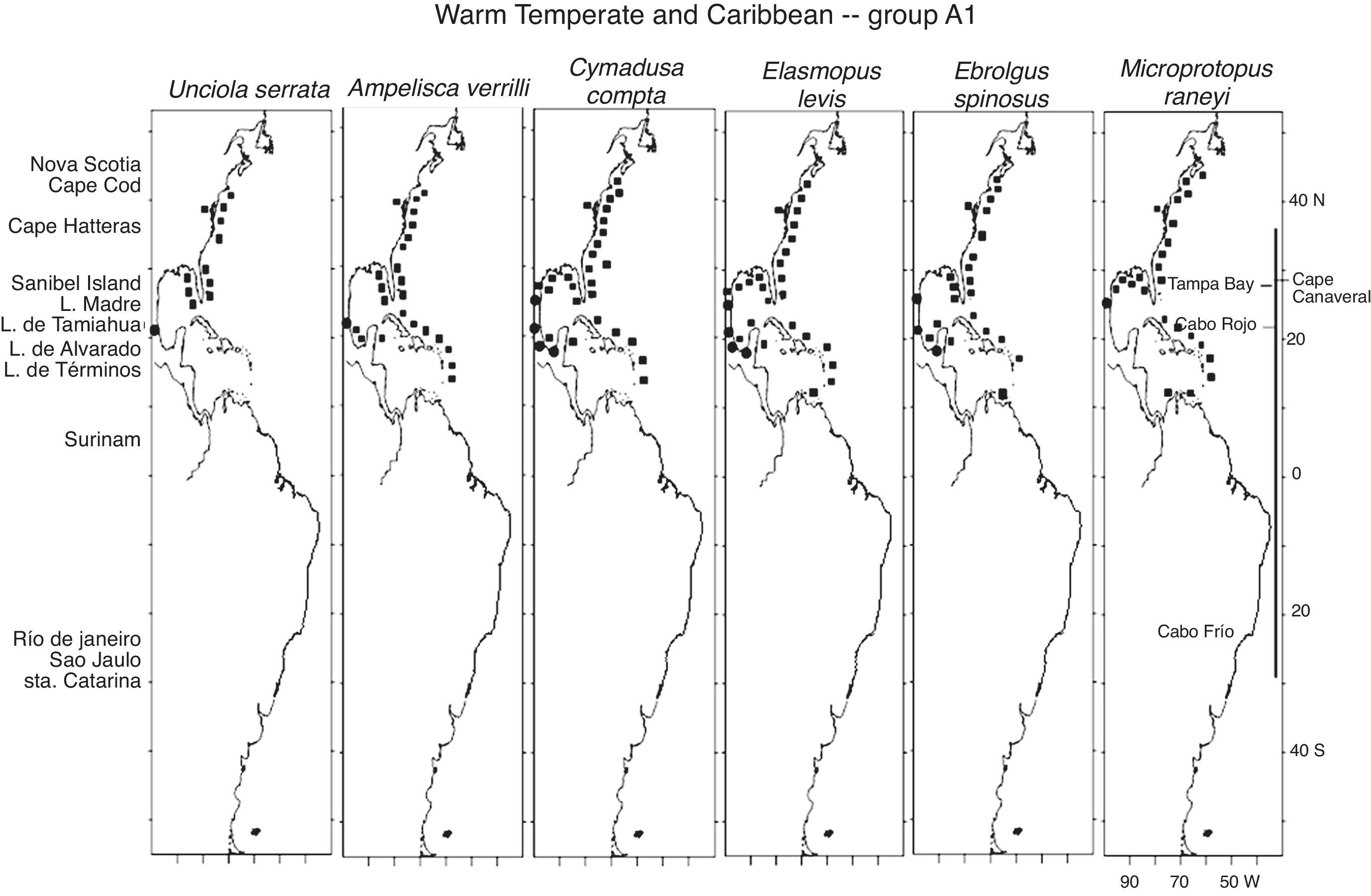
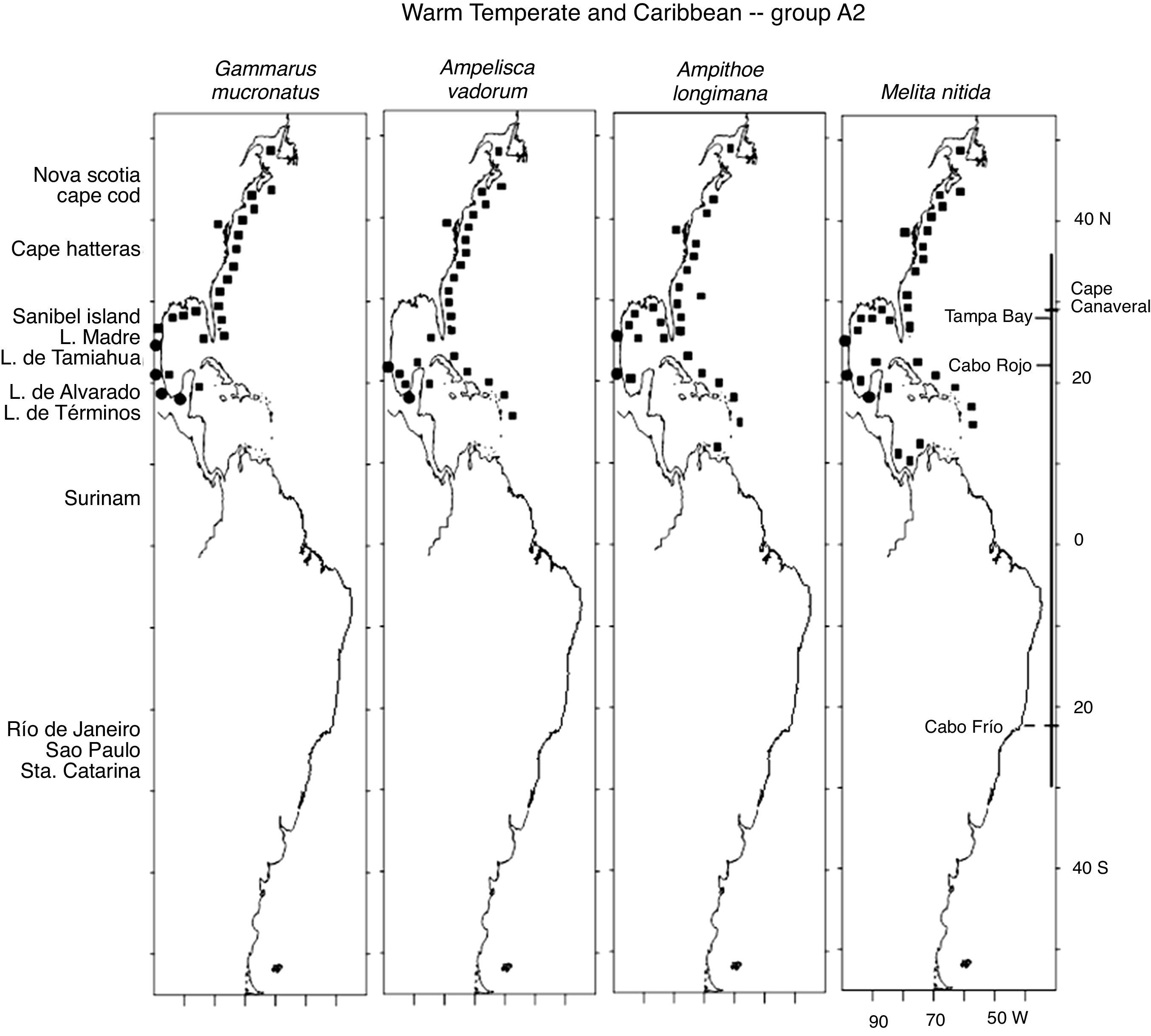
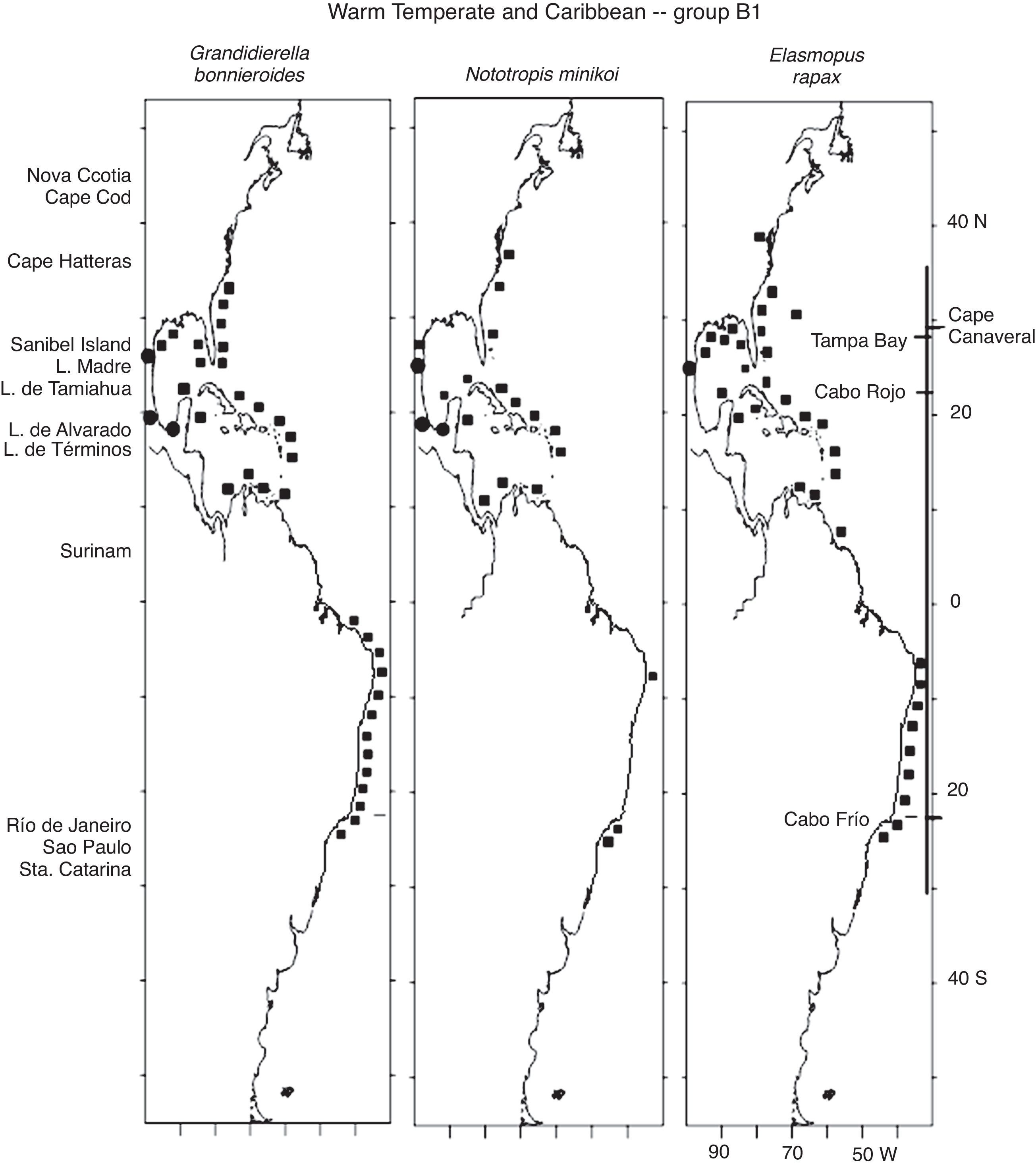
The 18 Warm Temperate and Caribbean species may be grouped into 5 sub-groups. Six species in group A1, Unciola serrata Shoemaker, 1945, Ampelisca verrilli Mills, 1967, C. compta, E. levis, E. spinosus and Microprotopus raneyi Wigley, 1966, present a mostly continuous distribution from Maine, south along the eastern coast of the United States, the Gulf of Mexico and throughout the Caribbean Sea (except for U. serrata which at present has no records in the Caribbean) (Fig. 3). The species in proup A2 follow the same distribution but include temperate habitats in Nova Scotia and a northernmost point in the Gulf of St Lawrence, Canada. These are G. mucronatus, A. vadorum, A. longimana and M. nitida. It is interesting to note that G. mucronatus has 1 single record in the Caribbean, in Quintana Roo, Mexico (Fig. 4).
 along the Western Atlantic (■) including the Mexican lagoons (●).")
 along the Western Atlantic (■) including the Mexican lagoons (●).")
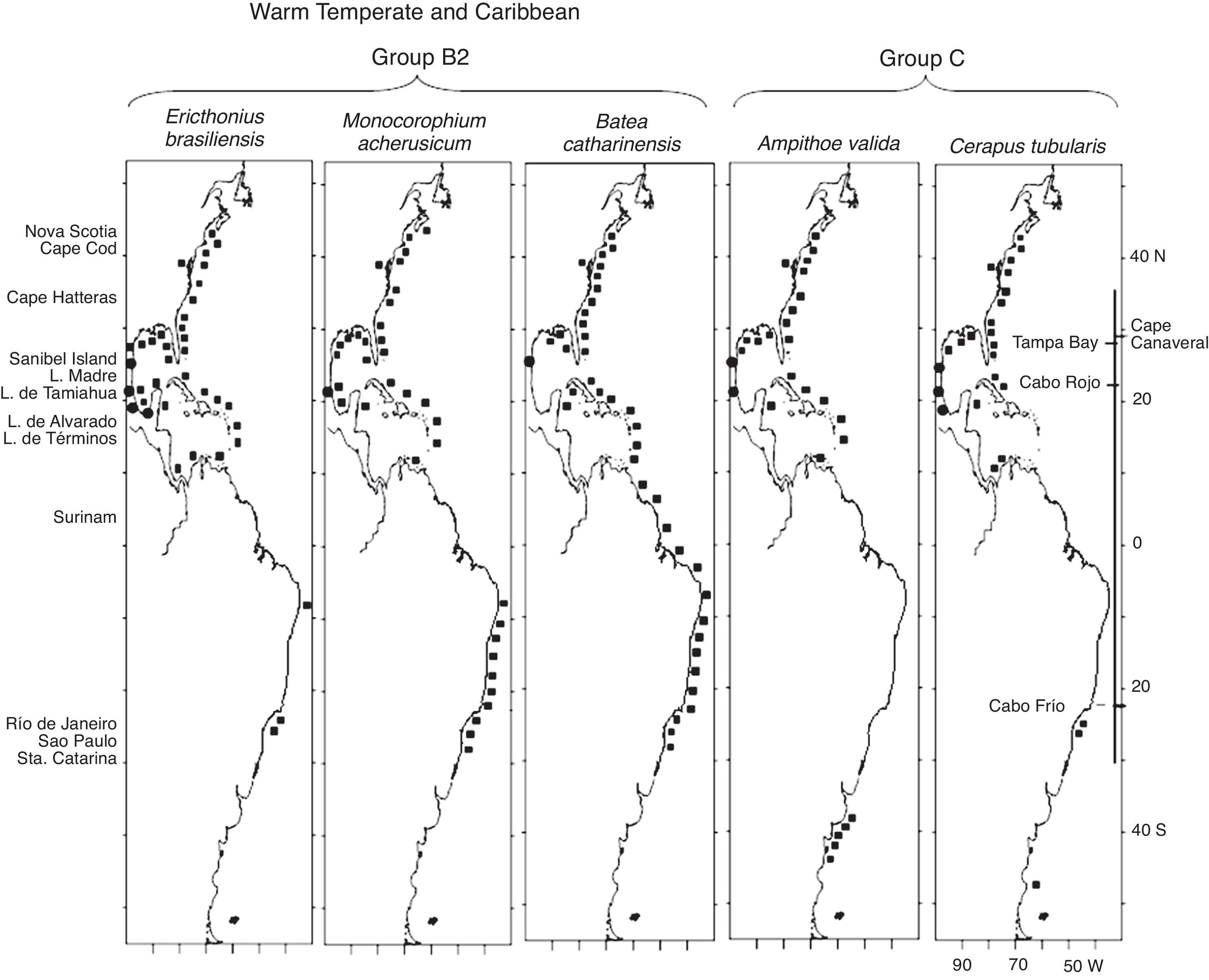
The species in group B1 are distributed along the southern half of the eastern coast of the United States, the Gulf of Mexico, the Caribbean Sea and the coast of Brazil, and include Grandidierella bonnieroidesStephensen, 1948, N. minikoi and Elasmopus rapax Costa, 1853 (Fig. 5). In particular, regarding G. bonnieroides, Myers (1970) reviewed its status and stated that specimens collected in the Gulf of Mexico, Caribbean Sea, Colombia, Venezuela, and Trinidad corresponded to this species. Asari and Myers (1982) later observed variations in specimens collected in the Western Atlantic and concluded that a complex of species was present. However, the 55 specimens from Laguna Madre were identified by A. Corona (pers comm), an expert in amphipod taxonomy, leaving no doubt as to the identity of the species. Three of the Warm Temperate and Caribbean species, C. compta, A. longimana and E. rapax, have also been recorded in Bermuda. The species in group B2, E. brasiliensis, M. acherusicum and B. catharinensis, have the same distribution but extend northwards to Nova Scotia (Fig. 6). The coastline between Venezuela and Brazil with no records in the case of some species most probably indicates a lack of studies throughout the area rather than a disjunct distribution of the species.
 along the Western Atlantic (■) including the Mexican lagoons (●).")
 and group C (Ampithoe valida, Cerapus tubularis) along the Western Atlantic (■) including the Mexican lagoons (●).")
The species in group C, A. valida and Cerapus tubularis Say, 1817, are distributed along the eastern coast of the United States from Maine to Florida, throughout the Gulf of Mexico, the Caribbean Sea, southern Brazil (C. tubularis) and include some southernmost points in Argentina (Fig. 6).
The warm and temperate affinity of these species is reflected in the wide range of geographic locations where they have been recorded and indicates a high ecologic plasticity, as Melo (1990) stated for South American brachyurans. The geographic locations previously recorded for these species, which latitudinally include the Gulf of Mexico, validate their presence in this area. As an example of this, 12 species reported here are amphiamerican or cosmopolitan: E. spinosus (Washington to California, USA; Baja California and Gulf of California, Mexico), G. mucronatus (California, USA), A. longimana (Baja California and Gulf of California, Mexico), M. nitida (British Columbia-Canada, Washington to California, USA, Baja California, Sinaloa and Michoacán, Mexico, Costa Rica, Panama, Ecuador, United Kingdom, Netherlands, Germany), G. bonnieroides (Tanzania, Mozambique, South Africa, Madagascar, Mauritius, India, Australia), N. minikoi (East India, South China Sea, Indo-Pacific), E. rapax (United Kingdom, Italy, Mediterranean Sea, Turkey, Red Sea, Australia), E. brasiliensis (California, USA, Baja California, Mexico, Norway, United Kingdom, France, Spain, Italy, Mediterranean Sea, Mauritius, Indian Ocean, South China Sea), M. acherusicum (Alaska to California, USA, Baja California, Mexico, Hawaii, United Kingdom, Netherlands, Mediterranean Sea, France, Italy, Egypt, Tunisia, Algeria, Senegal, Cape Colony, Dar es Salaam, Mauritius, Indian Ocean, Japan, South China Sea, Australia, New Zealand), B. catharinensis (Baja California, Mexico), A. valida (British Columbia, Canada to California, USA, Japan, Korea), and C. tubularis (California, USA to Baja California, Mexico, Japan, Korea) (Appendix 1).
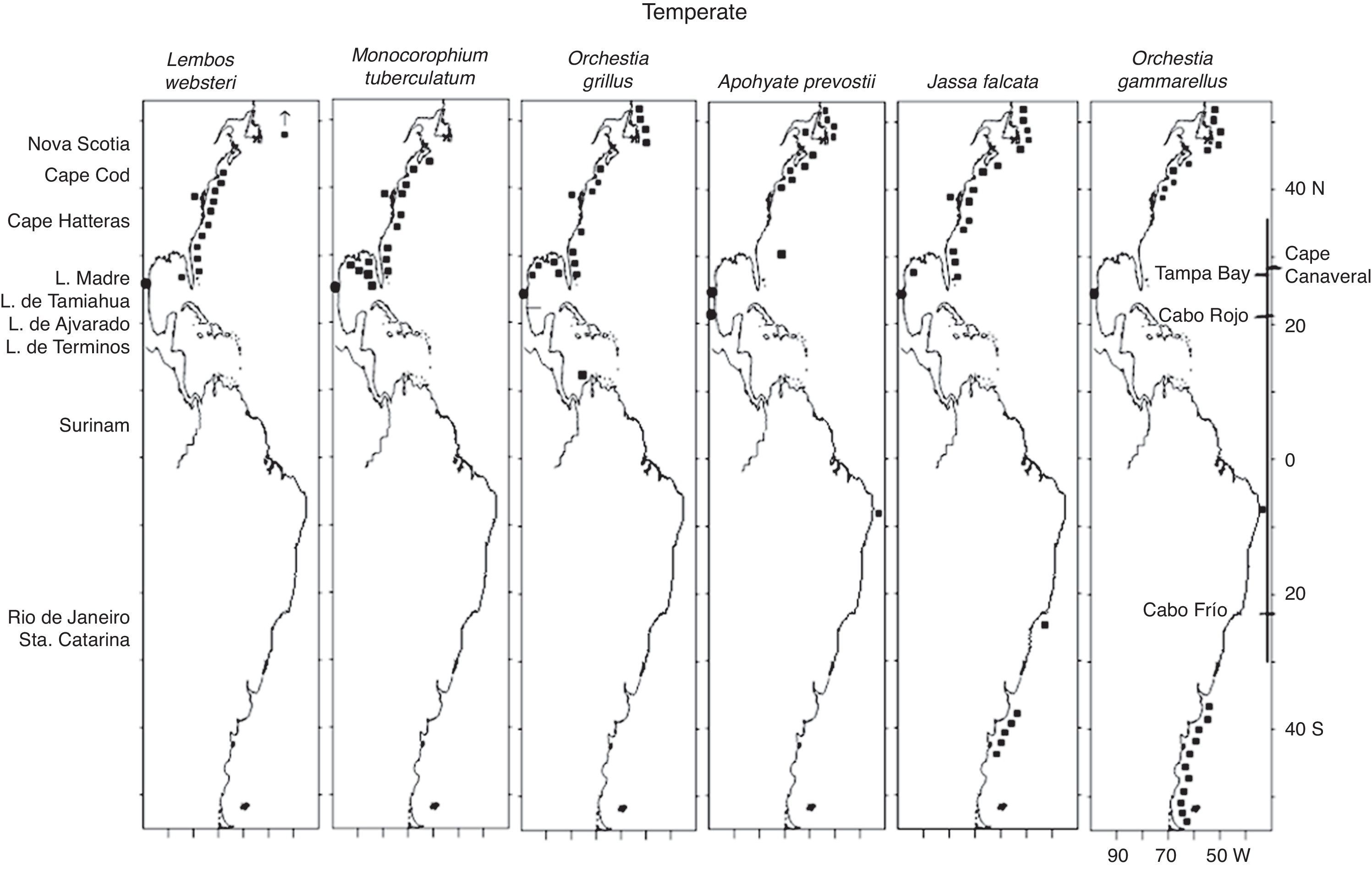
Among the 6 temperate species (Fig. 7), Lembos websteri Bate, 1857, Monocorophium tuberculatum (Shoemaker, 1934) and O. grillus are distributed from Maine to eastern Florida and in the Gulf of Mexico. L. websteri also has records in an extremely northerly point in Greenland, while O. grillus extends northwards to the Newfoundland coast and southwards to Colombia (this last point is considered an isolated record). M. tuberculatum is included in this group in spite of having 1 record in Florida Bay, in consideration of its clearly northern affinity and its absence from typically tropical habitats such as Laguna de Términos, Campeche where Ledoyer (1986) and Corona et al. (2000) did not record it in spite of thorough sampling. These species present a disjunct distribution at the tip of Florida where ocean water masses are more characteristically tropical, as is the case of the Carolinian zoogeographic province (Thurman, 1987).
 along the Western Atlantic (■) including the Mexican lagoons (●).")
The other 3 temperate species, A. prevostii, Jassa falcata (Montagu, 1808) and Orchestia gammarellus (Pallas, 1766), also present a northerly distribution up to Newfoundland. A. prevostii extends south to Laguna de Tamiahua, Veracruz (with 791 specimens collected in this study) at the northern tropical-temperate boundary for shallow water fauna within the western Gulf of Mexico, and includes an isolated record in eastern Brazil. J. falcata and O. gammarellus also present a southern temperate distribution with records in southern Brazil (J. falcata), an isolated point in eastern Brazil (O. gammarellus) and Argentina (both species). The presence of species in widely separated localities such as these generally responds to dispersal processes governed by ocean currents, rafting on plant debris, logs, sargassum and macroalgae, and transportation in ships’ ballast water (Appadoo & Steele, 1998).
L. websteri, M. tuberculatum, O. grillus and J. falcata, previously recorded in the northern Gulf of Mexico, may disperse west and south to the northern tropical-Temperate boundary at Cabo Rojo, Veracruz, by floating and/or rafting on the LATEX coastal current, as is discussed above in the section on the “Warm and Caribbean species” (Castilla & Guikez, 2000; Fernández et al., 1998; Havermans et al., 2007; Highsmith, 1985; Locke & Corey, 1989; Norton & Mathieson, 1983; Thiel, 2003a, 2003b; Williams & Bynum, 1972). The 6 temperate species were recorded in Laguna Madre, Tamaulipas, which lies north of this zoogeographic boundary.
As a confirmation of the temperate affinity of these species, other temperate localities have been reported for 4 of these species: for L. websteri Norway, England, France, Spain, Italy, Yugoslavia, Greece, Turkey, Israel, Egypt, the Mediterranean Sea and West Africa, for A. prevostii the Arctic Ocean, Iceland, Ireland, Norway, the United Kingdom, the Faroe Islands, the Netherlands, Belgium, France and Spain, for J. falcata Ireland, Norway, the United Kingdom, Spain, the Mediterranean Sea, British Columbia, Canada to California-United States and New Zealand, and for O. gammarellus Iceland, Norway, the United Kingdom, Spain, the Mediterranean Sea, the Black Sea, Turkey and SW Africa (Appendix 1).
The area with no records that may be seen for some species along the Central American coasts and the northern and northeastern South American coasts may, in general, be attributed to a lack of local studies. Future studies in these areas should thus contribute to filling the information gaps, particularly in the case of species in the Warm Temperate and Caribbean distribution pattern.
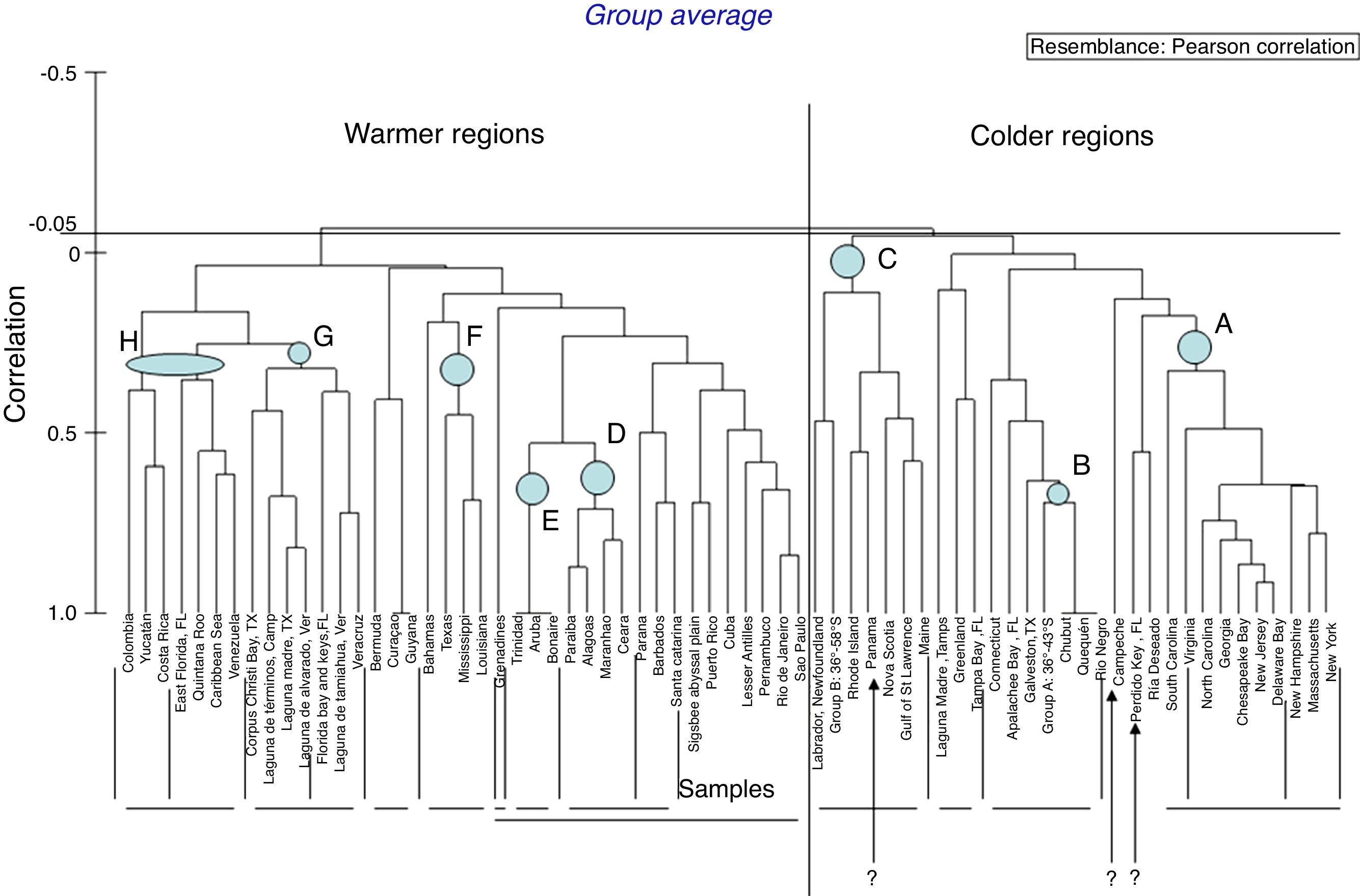
The grouping of warmer (+) and colder (●) geographic localities generated by the hierarchical cluster analysis based on the presence of gammarid and corophiid amphipods along the Western Atlantic (Figs. 8 and 9) indicates that these species, despite most being eurythermal, have a physiological preference for higher or lower water temperatures (Barba & Sánchez, 2005; d’Udekem-d’Acoz, & Verheye, 2013).
; D, northeastern Brazil; E, Caribbean; F, northwestern Gulf of Mexico; G, western, southern and eastern Gulf of Mexico, and H, Caribbean. Three interrogation marks (?) indicate odd localities within groups.")
Cluster diagram of geographic points recorded for the 29 amphipod species of Laguna Madre, Tamaulipas and Laguna de Tamiahua, Veracruz, Mexico, showing one group of warmer regions and another group of colder regions. Eight smaller groups may be defined as: A, eastern and northeastern United States; B, Argentina; C, northeastern United States, Canada and Argentina (and ? Panama); D, northeastern Brazil; E, Caribbean; F, northwestern Gulf of Mexico; G, western, southern and eastern Gulf of Mexico, and H, Caribbean. Three interrogation marks (?) indicate odd localities within groups.
 and colder (●) regions along the Western Atlantic based on the geographic points recorded for the 29 amphipod species of Laguna Madre, Tamaulipas and Laguna de Tamiahua, Veracruz, Mexico. The 2 slotted lines represent the zoogeographic boundaries that separate the warmer tropical regions from the colder Temperate regions along the Western Atlantic. The northern boundary spans along an imaginary line from Cabo Rojo in Veracruz to Tampa Bay and Cape Canaveral in Florida, and the southern boundary lies at Cabo Frío in Brazil.")
Grouping of the warmer (+) and colder (●) regions along the Western Atlantic based on the geographic points recorded for the 29 amphipod species of Laguna Madre, Tamaulipas and Laguna de Tamiahua, Veracruz, Mexico. The 2 slotted lines represent the zoogeographic boundaries that separate the warmer tropical regions from the colder Temperate regions along the Western Atlantic. The northern boundary spans along an imaginary line from Cabo Rojo in Veracruz to Tampa Bay and Cape Canaveral in Florida, and the southern boundary lies at Cabo Frío in Brazil.
This is the first comprehensive study of gammarid and corophiid amphipods of 2 major coastal lagoons in the western Gulf of Mexico that are characterised by their unique environmental conditions and are located at the northern tropical-temperate zoogeographical boundary that restricts the dispersion of tropical shallow water fauna. It may be stated that these estuarine amphipods may be further analysed, not only due to their abundance and diversity but because some species may be considered good indicators of zoogeographic provinces along the Western Atlantic.
To A. Corona for sample processing, identification of species and the note on G. bonnieroides; N. López for identifying the macroalgae; L. Huidobro and D. Salcedo for running the cluster analyses; M. Thiel for reviewing an earlier version of the manuscript, and especially I. Winfield for reviewing the final draft. Conabio provided the funds to carry out this research (project H258).
A=www.geocities.com/ajuarrero/amphipods.html
B=www.bdt.fat.org.br/workshop/costa/praias/tabela3
C=www.acd.ofrj.br/mndi/Carcinologia/hp/Text/Amphipoda1.htm
| Geographic distribution | References |
|---|---|
| Warm and Caribbean | |
| Ampelisca venetiensis ++ Figure 2 Laguna de Tamiahua in Veracruz, Mexico; Costa Rica; Caribbean Sea; Colombia. | Escobar, Winfield, Ortiz, Gasca, & Suárez, 2002; Escobar & Winfield, 2003; Ortiz, Martín, & Díaz, 2007; Miloslavich et al., 2010; Winfield & Ortiz, 2011; Martín et al., 2013 |
| Hourstonius lagunaFigure 2 St John's River, Indian River Lagoon, Biscayne Bay, Florida Bay in Florida, Mississippi, and Galveston, San Antonio Bay, Corpus Christi Bay, Laguna Madre in Texas, US; Laguna Madre in Tamaulipas, Laguna de Tamiahua, Laguna Camaronera and Laguna de Alvarado in Veracruz, Laguna de Términos in Campeche, Quintana Roo, Mexico; Bermuda; Caribbean Sea; Cuba; Colombia; Venezuela. – California, US. | McKinney, 1978; Ledoyer, 1986; Barnard & Karaman, 1991; Ortiz & Lalana, 1993; Nelson, 1995; Camp, Lyons, & Perkins, 1998; Corona et al., 2000; Cházaro et al., 2002; LeCroy, 2002; Ortiz et al., 2007; Winfield, Cházaro-Olvera, et al., 2007; Miloslavich et al., 2010; Winfield & Ortiz, 2011; Martín et al., 2013 |
| Parhyale hawaiensis +++ Figure 2 Florida, Louisiana, US; Gulf of Mexico; Soto la Marina in Tamaulipas, Quintana Roo, Mexico; Caribbean Sea; Cuba; Haiti; Dominican Republic; Puerto Rico; Venezuela; Aruba; Curaçao; Bonaire; Lesser Antilles; Rio de Janeiro to Paraná, Brazil. – Circumtropical; California, US; Atlantic and Pacific oceans; Hawaii; Fiji; Philippines; South China Sea; Mediterranean Sea. | Shoemaker, 1956; Thomas, 1976; Myers, 1985; Camp et al., 1998; Wakabara & Serejo, 1998; Serejo, 1999; Lowry, 2000; Ortiz et al., 2007; Somaio & Moreira, 2008; LeCroy, Gasca, Winfield, Ortiz, & Escobar-Briones, 2009; Miloslavich et al., 2010; Ortiz & Lalana, 2010; Ortega, 2013 |
| Photis longicaudata ++ Figure 2 Florida, US; Laguna de Tamiahua in Veracruz, Quintana Roo, Mexico; Caribbean Sea; Venezuela; Pernambuco, Alagoas, Espirito Santo, Rio de Janeiro, São Paulo, Rio Grande do Sul, Brazil. – Atlantic and Indian oceans; Sweden; Ireland; Scotland; England; Belgium; France; Spain; Greece; Mediterranean Sea; Black Sea; Japan. | Wakabara, Tararam, Valério-Berardo, Duleba, & Pereira, 1991; Camp et al., 1998; Wakabara & Serejo, 1998; Escobar et al., 2002; Ortiz et al., 2007; Miloslavich et al., 2010; Winfield & Ortiz, 2011; Martín et al., 2013; www.marinespecies.org |
| Elasmopus pectenicrus +++ Figure 2 Florida, Florida Keys, US; Gulf of Mexico; Laguna Madre in Tamaulipas, Quintana Roo, Mexico; Caribbean Sea; Cuba; Lesser Antilles; west, central and east coasts of Venezuela; Ceara to Paraná, Brazil. – Circumtropical; Atlantic, Pacific and Indian oceans; Hawaii; Mediterranean Sea; Turkey; Red Sea; western and southern Africa; Madagascar; Mauritius; India; Indonesia; South China Sea; New Guinea; Australia. | Barnard, 1970; McKinney, 1977; Ortiz, 1978; Karaman, 1982; Wakabara, Tararam, & Takeda, 1983; Hay, Duffy, Fenical, & Gustafson, 1988; Appadoo & Steele, 1998; Camp et al., 1998; Wakabara & Serejo, 1998; Lowry, 2000; Díaz & Martín, 2001; Escobar et al., 2002; Appadoo & Myers, 2003; Martín & Díaz, 2003; Ortiz et al., 2007; LeCroy et al., 2009; Bakir, Sezgin, & Katagan, 2010; Miloslavich et al., 2010; Ortiz & Lalana, 2010; Martín et al., 2013; Ortega, 2013; Ortega & Martín, 2013 |
| Warm Temperate and Caribbean | |
| Unciola serrata ++ Figure 3 Cape Cod in Massachusetts to Delaware Bay and Virginia, Florida, US; Laguna de Tamiahua in Veracruz, Mexico. | Dickinson, Wigley, Brodeur, & Brown-Leger, 1980; Camp et al., 1998; Winfield & Ortiz, 2011; www.marinespecies.org |
| Ampelisca verrilli ++ Figure 3 Cape Cod in Massachusetts to Delaware Bay and Chesapeake Bay, Virginia to Cape Hatteras in North Carolina, Indian River Lagoon in Florida, US; Laguna de Tamiahua and Veracruz reef system in Veracruz, Campeche, Quintana Roo, Mexico; Caribbean Sea. | Dickinson et al., 1980; Nelson, 1995; Camp et al., 1998; Eggleston, Elis, Etherington, Dahlgren, & Posey, 1999; Escobar et al., 2002; Escobar & Winfield, 2003; Winfield, Escobar, & Morrone, 2006; Ortiz et al., 2007; Winfield & Escobar, 2007; Cerrato et al., 2010; Miloslavich et al., 2010; Winfield & Ortiz, 2011; Martín et al., 2013; www.marinespecies.org |
| Cymadusa comptaFigure 3 Maine, New Hampshire to Cape Cod, Pleasant Bay, Nantucket, Martha's Vineyard and Buzzards Bay in Massachusetts, Rhode Island, Connecticut, New Jersey, Delaware Bay, Chesapeake Bay, North Carolina to Indian River Lagoon, Riviera Beach, Biscayne Bay in Florida and Florida Keys to Texas, Laguna Madre in Texas, US; Laguna Madre and Soto la Marina in Tamaulipas, Laguna de Tamiahua, Laguna de Alvarado and Veracruz reef system in Veracruz, Laguna de Términos in Campeche, Quintana Roo, Mexico; Bermuda; Caribbean Sea; Cuba. | Watling & Maurer, 1972; Bousfield, 1973; Ledoyer, 1986; Ortiz & Lalana, 1993; Thomas, 1993a; Nelson, 1995; Camp et al., 1998; Eggleston et al., 1999; Corona et al., 2000; Cházaro et al., 2002; LeCroy, 2002; Varela, Manuel, & Rogelio, 2003; Barba & Sánchez, 2005; Ortiz et al., 2007; Miloslavich et al., 2010; Ortiz & Lalana, 2010; Martín et al., 2013; Ortega, 2013 |
| Elasmopus levisFigure 3 Maine, New Hampshire, Cape Cod, Vineyard Sound, Woods Hole, Buzzards Bay in Massachusetts, Rhode Island, Connecticut, New York, New Jersey, Delaware Bay, Chesapeake Bay, Virginia, North Carolina to Georgia, Biscayne Bay and Florida Keys, Port Isabel, San Antonio Bay, Corpus Christi Bay, Laguna Madre in Texas, US; Gulf of Mexico; Laguna Madre and Soto la Marina in Tamaulipas, Laguna de Tamiahua, Laguna de Alvarado and Veracruz reef system in Veracruz, Laguna de Términos in Campeche, Yucatán, Quintana Roo, Mexico; Caribbean Sea; Venezuela. | Watling & Maurer, 1972; Bousfield, 1973; McKinney, 1977; Dickinson et al., 1980; Ledoyer, 1986; Ortiz & Lalana, 1993; Camp et al., 1998; Eggleston et al., 1999; Corona et al., 2000; LeCroy, 2000; Escobar et al., 2002; Barba & Sánchez, 2005; Ortiz et al., 2007; Winfield, Abarca-Arenas, et al., 2007; Cerrato et al., 2010; Miloslavich et al., 2010; Winfield & Ortiz, 2011; Martín et al., 2013; Ortega, 2013; Paz-Ríos & Ardisson, 2013 |
| Eobrolgus spinosusFigure 3 Maine, New Hampshire, Cape Cod in Massachusetts, Rhode Island, Connecticut, New York to Delaware Bay, Chesapeake Bay, North Carolina, Georgia, Florida, Florida Bay, Florida Keys, Charlotte Harbor, Tampa Bay to Apalachee Bay in Florida, US; Laguna Madre in Tamaulipas, Laguna de Tamiahua in Veracruz, southwestern Gulf of Mexico, Laguna de Términos and Campeche Bay in Campeche, Arrecife Alacranes in Yucatán, Quintana Roo, Mexico; Caribbean Sea; Cuba; Venezuela. – Washington to California, US; Bahía San Quintín, Baja California and Gulf of California, Mexico. | Barnard, 1970; Watling & Maurer, 1972; Bousfield, 1973; Barnard & Barnard, 1981; Ledoyer, 1986; Camp et al., 1998; Eggleston et al., 1999; Corona et al., 2000; Díaz & Martín, 2001; Martín, Ortiz, & Díaz, 2002; Escobar & Winfield, 2003; Martín & Díaz, 2003; Winfield et al., 2006; Ortiz et al., 2007; Cerrato et al., 2010; Miloslavich et al., 2010; Ortiz & Lalana, 2010; LeCroy, 2011; Winfield & Ortiz, 2011; Martín et al., 2013; Paz-Ríos, Simões, & Ardisson, 2013 |
| Microprotopus raneyiFigure 3 Canada; Maine, New Hampshire, Cape Cod Bay, Martha's Vineyard, Vineyard Sound, Buzzards Bay, Woods Hole in Massachusetts, Rhode Island, Connecticut, New York, New Jersey, Delaware Bay, Chesapeake Bay, Virginia, North Carolina, Edisto Island in South Carolina, St Simon's Island in Georgia, north Florida, Sarasota Bay and Perdido Key in Florida, Louisiana, Texas, US; northern Gulf of Mexico; Laguna Madre in Tamaulipas, Mexico; Caribbean Sea; Colombia; Venezuela. | Lowry, 1972; Watling & Maurer, 1972; Bousfield, 1973; Dickinson et al., 1980; Barnard & Karaman, 1991; Rakocinski, Heard, LeCroy, McLelland, & Simons, 1993; Camp et al., 1998; Eggleston et al., 1999; Díaz & Martín, 2001; Barba & Sánchez, 2005; Ortiz et al., 2007; Dubois, Gelpi, Condrey, Grippo, & Fleeger, 2009; Miloslavich et al., 2010; Martín et al., 2013; Ortega & Martín, 2013 |
| Gammarus mucronatusFigure 4 Gulf of St Lawrence and Nova Scotia, Canada; Maine, New Hampshire, Massachusetts, Rhode Island, Connecticut, New York, New Jersey, Delaware Bay, Chesapeake Bay, Virginia, North Carolina to Florida and Florida Keys, Louisiana, Laguna Madre in Texas, US; northern Gulf of Mexico; Laguna Madre and Soto la Marina in Tamaulipas, Laguna de Tamiahua, Laguna de Alvarado and continental shelf of Veracruz, southwestern Gulf of Mexico, Laguna de Términos in Campeche, Quintana Roo, Mexico. – California, US. | Watling & Maurer, 1972; Bousfield, 1973; Thomas, 1976; Lincoln, 1979; Dickinson et al., 1980; Heard, 1982; Ledoyer, 1986; Ortiz & Lalana, 1993; Nelson, 1995; Winfield et al., 1997; Camp et al., 1998; Eggleston et al., 1999; Cházaro et al., 2002; Escobar & Winfield, 2003; Barba & Sánchez, 2005; Ortiz et al., 2007; Winfield, Cházaro-Olvera, et al., 2007; Miloslavich et al., 2010; Winfield & Ortiz, 2011; Martín et al., 2013; Ortega, 2013 |
| Ampelisca vadorumFigure 4 SW Gulf of St Lawrence, Nova Scotia, Canada; Maine, New Hampshire, Cape Cod in Massachusetts, Rhode Island, Connecticut, New York, New Jersey, Delaware Bay, Chesapeake Bay, Virginia, North Carolina, South Carolina, Georgia, Florida and Florida Keys, US; Laguna Madre in Tamaulipas, Laguna de Tamiahua and continental shelf off Alvarado in Veracruz, southwestern Gulf of Mexico, Laguna de Términos and Bay of Campeche in Campeche, Yucatán, Quintana Roo, Mexico; Caribbean Sea; Cuba. | Mills, 1963; Watling & Maurer, 1972; Bousfield, 1973; Dickinson et al., 1980; Ledoyer, 1986; Barnard & Karaman, 1991; Ortiz & Lalana, 1993; Nelson, 1995; Camp et al., 1998; Eggleston et al., 1999; Corona et al., 2000; LeCroy, 2002; Escobar & Winfield, 2003; Winfield et al., 2006; Ortiz et al., 2007; Winfield & Escobar, 2007; Cerrato et al., 2010; Miloslavich et al., 2010; Ortiz & Lalana, 2010; Martín et al., 2013; Paz-Ríos & Ardisson, 2013 |
| Ampithoe longimanaFigure 4 SW Gulf of St Lawrence, Canada; Maine, Cape Cod in Massachusetts, Chesapeake Bay to North Carolina and Indian River Lagoon, Biscayne Bay, Florida Bay, Florida Keys, Tampa Bay in Florida to South Padre Island in Texas, US; Laguna Madre in Tamaulipas, Laguna de Tamiahua and Veracruz reef system in Veracruz, Mexico; Bermuda; Bahamas; Caribbean Sea; Venezuela – West Baja California, Gulf of California, Mexico. | Barnard, 1963; Bousfield, 1973; Ortiz & Lalana, 1993; Nelson, 1995; Camp et al., 1998; Eggleston et al., 1999; LeCroy, 2002; Martín & Díaz, 2003; Ortiz et al., 2007; Miloslavich et al., 2010; Winfield & Ortiz, 2011; Winfield et al., 2011; Martín et al., 2013; Winfield, 2013 |
| Melita nitidaFigure 4 SW Gulf of St Lawrence, Nova Scotia, Canada; Maine, New Hampshire, Cape Cod in Massachusetts, Rhode Island, Connecticut, New York, New Jersey, Delaware Bay, Chesapeake Bay, Virginia, North Carolina, Georgia, Cape Romano in Florida, Louisiana, US; Gulf of Mexico; Laguna Madre in Tamaulipas, Laguna de Tamiahua and Veracruz reef system in Veracruz, Laguna de Términos in Campeche, Yucatán, Quintana Roo, Mexico; Costa Rica; Caribbean Sea; Cuba; Panama; Colombia.–British Columbia, Canada; Washington to California, US; Baja California, Mazatlán in Sinaloa, Laguna Salinas del Padre and Coahuayana River in Michoacán, Mexico; Costa Rica; Panama; Ecuador; United Kingdom; Netherlands; Germany. | Watling & Maurer, 1972; Bousfield, 1973; Thomas, 1976; Lincoln, 1979; Dickinson et al., 1980; Heard, 1982; Ortiz, 1983; Chapman, 1988; Ortiz & Lalana, 1993; Nelson, 1995; Camp et al., 1998; Eggleston et al., 1999; Corona et al., 2000; LeCroy, 2000; Corona & Raz-Guzmán, 2003; Faasse & van Moorsel, 2003; Ortiz et al., 2007; Cerrato et al., 2010; Miloslavich et al., 2010; Winfield et al., 2011; Martín et al., 2013; Winfield, 2013; A |
| Grandidierella bonnieroidesFigure 5 North Carolina, Georgia, east Florida, Florida Keys, west Florida, Louisiana, Texas, US; Laguna Madre and Soto la Marina in Tamaulipas, Laguna de Alvarado in Veracruz, Laguna de Términos in Campeche, Yucatán, Quintana Roo, Mexico; Caribbean Sea; Cuba; Colombia; Venezuela; Trinidad and Tobago; Aruba; Curaçao; Bonaire; Maranhao to São Paulo, Brazil. – Tanzania; Mozambique; South Africa; Madagascar; Mauritius; India; Australia. | Fox & Bynum, 1975; Thomas, 1976; Leite, Tararam, & Wakabara, 1980; Asari & Myers, 1982; Heard, 1982; Ledoyer, 1986; Wakabara et al., 1991; Ortiz & Lalana, 1993; Nelson, 1995; Winfield et al., 1997; Camp et al., 1998; Wakabara & Serejo, 1998; Corona et al., 2000; Ortiz, 2001; Winfield et al., 2001; Cházaro et al., 2002; LeCroy, 2002; Martín & Díaz, 2003; Appadoo & Myers, 2004; Ortiz et al., 2007; Winfield, Cházaro-Olvera, et al., 2007; Miloslavich et al., 2010; Ortiz & Lalana, 2010; Winfield & Ortiz, 2011; Martín et al., 2013; Ortega, 2013; Paz-Ríos & Ardisson, 2013; A, B, C |
| Nototropis minikoiFigure 5 Virginia, North Carolina, Florida, Laguna Madre in Texas, US; Laguna Madre and Soto la Marina in Tamaulipas, Laguna de Tamiahua, Laguna Camaronera and Laguna de Alvarado in Veracruz, Laguna de Términos in Campeche, Arrecife Alacranes in Yucatán, Quintana Roo, Mexico; Costa Rica; Caribbean Sea; Cuba; Colombia; Barlovento, Isla Margarita in Venezuela; Pernambuco, Rio de Janeiro and São Paulo, Brazil. – Karwar Bay, East India; South China Sea; Indo Pacific. | Stephensen, 1948; Oliveira, 1953; Bousfield, 1973; Bynum & Fox, 1977; Naomi, 1979; Leite et al., 1980; Ledoyer, 1986; Lagarde, 1987; Barnard & Karaman, 1991; Wakabara et al., 1991; Ortiz & Lalana, 1993; Wakabara, Nicoletti, & Tararam, 1996; Wakabara & Serejo, 1998; Lowry, 2000; Cházaro et al., 2002; Lowry & Stoddart, 2003; Martín & Díaz, 2003; Barba & Sánchez, 2005; McLaughlin et al., 2005; Ortiz et al., 2007; LeCroy et al., 2009; Miloslavich et al., 2010; Ortiz & Lalana, 2010; Winfield & Ortiz, 2011; Martín et al., 2013; Ortega & Martín, 2013; Paz-Ríos et al., 2013; A, B, C |
| Elasmopus rapax +++ Figure 5 Delaware Bay, South Carolina, Georgia to Florida Keys, west Florida, Louisiana, Texas, US; Gulf of Mexico; Laguna Madre in Tamaulipas, Cayo Arcas in Yucatán, Quintana Roo, Mexico; Bermuda; Caribbean Sea; Cuba; Lesser Antilles; Curaçao; east coast of Venezuela; Guyana; Paraiba to São Paulo, Brazil. – Cosmopolitan tropical and temperate; Atlantic, Pacific and Indian oceans; United Kingdom; Italy; Mediterranean Sea; Turkey; Red Sea; eastern Australia. | Shoemaker, 1933; Barnard, 1962; Bousfield, 1973; McKinney, 1977; Ortiz, 1978; Lincoln, 1979; Ortiz, 1979; Karaman, 1982; Ruffo, 1982; Galan, 1984; González, 1991; Wakabara et al., 1991; Procaccini & Scipione, 1992; Camp et al., 1998; Wakabara & Serejo, 1998; LeCroy, 2000; Escobar et al., 2002; LeCroy, 2002; Martín & Díaz, 2003; Ortiz et al., 2007; Winfield & Escobar, 2007; LeCroy et al., 2009; Miloslavich et al., 2010; Ortiz & Lalana, 2010; Martín et al., 2013; Ortega, 2013; Paz-Ríos & Ardisson, 2013; http://www.skaphandrus.com |
| Ericthonius brasiliensisFigure 6 Maine, New Hampshire, Cape Cod, Vineyard Sound in Massachusetts, Rhode Island, Connecticut, New York, New Jersey, Delaware Bay, Chesapeake Bay, Virginia, North Carolina, Georgia, Florida and Florida Keys, Perdido Key, Louisiana, Texas, Port Isabel and Laguna Madre in Texas, US; Laguna Madre and Soto la Marina in Tamaulipas, Laguna de Tamiahua, Laguna Camaronera, Laguna de Alvarado and Veracruz reef system in Veracruz, southwestern Gulf of Mexico, Laguna de Términos in Campeche, Yucatán, Quintana Roo, Mexico; Sigsbee abyssal plain; Costa Rica; Caribbean Sea; Cuba; Lesser Antilles; Colombia; Venezuela; Pernambuco, Rio de Janeiro and São Paulo, Brazil. – Circumtropical cosmopolitan; Atlantic, Pacific and Indian oceans; California, US; Bahía San Quintín, Baja California, Mexico; Norway; United Kingdom; France; Spain; Italy; Mediterranean Sea; Mauritius; South China Sea. | Barnard, 1970; Watling & Maurer, 1972; Bousfield, 1973; McKinney, 1977; Lincoln, 1979; Dickinson et al., 1980; Leite et al., 1980; Ruffo, 1982; Myers & McGrath, 1984; Wakabara et al., 1991; Procaccini & Scipione, 1992; Ortiz & Lalana, 1993; Rakocinski et al., 1993; Thomas, 1993a; Nelson, 1995; Appadoo & Steele, 1998; Camp et al., 1998; Wakabara & Serejo, 1998; Eggleston et al., 1999; Lowry, 2000; Cházaro et al., 2002; Escobar & Winfield, 2003; Martín & Díaz, 2003; Varela et al., 2003; Appadoo & Myers, 2004; Barba & Sánchez, 2005; Winfield et al., 2006; Ortiz et al., 2007; Winfield, Abarca-Arenas, et al., 2007; Dubois et al., 2009; Cerrato et al., 2010; Miloslavich et al., 2010; Ortiz & Lalana, 2010; Winfield et al., 2010; Winfield & Ortiz, 2011; Martín et al., 2013; Ortega, 2013; Paz-Ríos & Ardisson, 2013; A, B, C |
| Monocorophium acherusicumFigure 6 Bay of Fundy and Nova Scotia, Canada; Maine, New Hampshire, Massachusetts, Rhode Island, Connecticut, New Jersey, Delaware Bay, Chesapeake Bay, North Carolina, Georgia, Florida and Florida Keys, west Florida to Texas, US; Laguna de Tamiahua and Veracruz reef system in Veracruz, southwestern Gulf of Mexico, Quintana Roo, Mexico; Sigsbee abyssal plain; Cuba; Puerto Rico; Lesser Antilles; Venezuela; Pernambuco to Santa Catarina, Brazil. – Alaska to California, US; Bahía San Quintín, Baja California, Mexico; Hawaii; United Kingdom and the Netherlands south to the Mediterranean Sea including France, Italy, Egypt, Tunisia and Algeria; Senegal; Cape Colony; Dar es Salaam; Mauritius; Indian Ocean; Japan; South China Sea; Australia; New Zealand. | Shoemaker, 1934; Barnard, 1970, 1971, 1972; Watling & Maurer, 1972; Bousfield, 1973; Lincoln, 1979; Myers, 1982; Ruffo, 1982; Barnard & Karaman, 1991; Wakabara et al., 1991; Procaccini & Scipione, 1992; Ortiz & Lalana, 1993; Nelson, 1995; Appadoo & Steele, 1998; Camp et al., 1998; Wakabara & Serejo, 1998; Eggleston et al., 1999; Lowry, 2000; Appadoo & Myers, 2004; LeCroy, 2004; Winfield et al., 2006; Ortiz et al., 2007; Somaio & Moreira, 2008; Miloslavich et al., 2010; Ortiz & Lalana, 2010; Winfield et al., 2011; Martín et al., 2013; Ortega & Martín, 2013; Winfield, 2013; A, B |
| Batea catharinensisFigure 6 Maine, New Hampshire, Cape Cod in Massachusetts, Rhode Island, Connecticut, Long Island Sound in New York, New Jersey, Delaware Bay, Chesapeake Bay, Virginia, North Carolina, South Carolina, Georgia, Florida, Tampa Bay, Mississippi Sound, Louisiana, US; Laguna Madre in Tamaulipas, Quintana Roo, Mexico; Caribbean Sea; Cuba; Barbados to São Paulo and Santa Catarina, Brazil. – Cedros Island and Magdalena Bay in Baja California, Mexico. | Barnard, 1972; Watling & Maurer, 1972; Bousfield, 1973; Dickinson et al., 1980; Wakabara et al., 1991; Ortiz & Lalana, 1993; Nelson, 1995; Camp et al., 1998; Wakabara & Serejo, 1998; Eggleston et al., 1999; Escobar & Winfield, 2003; LeCroy, 2004; Barba & Sánchez, 2005; Ortiz et al., 2007; Cerrato et al., 2010; Miloslavich et al., 2010; Ortiz & Lalana, 2010; Martín et al., 2013; Ortega, 2013; A, B, C |
| Ampithoe validaFigure 6 Maine, New Hampshire, Cape Cod in Massachusetts, Rhode Island, Connecticut, Long Island Sound in New York, New Jersey, Delaware Bay, Chesapeake Bay, North Carolina, South Carolina, Georgia, Cape Canaveral in Florida, Apalachee Bay in Florida to Galveston Bay in Texas, US; Laguna Madre in Tamaulipas, Laguna de Tamiahua in Veracruz, Quintana Roo, Mexico; Caribbean Sea; Venezuela; Quequén, Rio Negro, Chubut=Group A: 36°–43°S, Argentina. – British Columbia and Vancouver, Canada to Newport Bay in California, US; Japan; Korea. | Watling & Maurer, 1972; Bousfield, 1973; Fox & Bynum, 1975; Alonso, Tablado, López-Gappa, & Magaldi, 1995; Camp et al., 1998; Eggleston et al., 1999; LeCroy, 2002; Martín & Díaz, 2003; López Gappa et al., 2006; Ortiz et al., 2007; Miloslavich et al., 2010; Martín et al., 2013 |
| Cerapus tubularisFigure 6 Maine, New Hampshire, Cape Cod in Massachusetts, Rhode Island, Connecticut, New York, New Jersey, Delaware Bay, Chesapeake Bay, Virginia, North Carolina, Georgia, east Florida, Perdido Key, Louisiana, Texas, US; Laguna Madre in Tamaulipas, Laguna de Tamiahua and Laguna de Alvarado in Veracruz, Quintana Roo, Mexico; Cuba; Colombia; São Paulo and Paraná, Brazil; Ría Deseado, Argentina. – San Diego in California, US to Ensenada in Baja California, Mexico; Japan; Korea. | Watling & Maurer, 1972; Bousfield, 1973; Dickinson et al., 1980; Barnard & Karaman, 1991; Wakabara et al., 1991; Rakocinski et al., 1993; Nelson, 1995; Camp et al., 1998; Wakabara & Serejo, 1998; Barba & Sánchez, 2005; LeCroy, 2007; Ortiz et al., 2007; Dubois et al., 2009; Miloslavich et al., 2010; A, B, C |
| Temperate | |
| Lembos websteri + Figure 7 Greenland; Maine, New Hampshire, Cape Cod, Vineyard Sound in Massachusetts, Rhode Island, Connecticut, New York, New Jersey, Delaware Bay, Chesapeake Bay, Virginia, North Carolina to Georgia and north Florida, west Florida, US; Laguna Madre in Tamaulipas, Mexico. – Norway; England; France; Spain; Italy; Yugoslavia; Greece; Turkey; Israel; Egypt; Algeria in the Mediterranean Sea; west Africa. | Bousfield, 1973; Dickinson et al., 1980; Ruffo, 1982; Barnard & Karaman, 1991; Ortiz & Lalana, 1993; Camp et al., 1998; Eggleston et al., 1999; LeCroy, 2002; Escobar & Winfield, 2003; Barba & Sánchez, 2005; Ortiz et al., 2007 |
| Monocorophium tuberculatumFigure 7 Minas Basin, Bay of Fundy and Nova Scotia, Canada; Maine, New Hampshire, Cape Cod in Massachusetts, Rhode Island, Connecticut, New York, New Jersey, Delaware Bay, Chesapeake Bay, Virginia, North Carolina, Georgia, Sebastian inlet in Florida, Florida Bay to Chandeleur Sound in Louisiana, US; Laguna Madre in Tamaulipas, Mexico. | Shoemaker, 1934, 1947; Watling & Maurer, 1972; Bousfield, 1973; Dickinson et al., 1980; Bousfield & Hoover, 1997; Camp et al., 1998; LeCroy, 2004; Ortiz et al., 2007; Dubois et al., 2009; Ortega, 2013 |
| Orchestia grillusFigure 7 Newfoundland, Canada; Maine, New Hampshire, Massachusetts, Rhode Island, Connecticut, New Jersey, Delaware Bay, Chesapeake Bay, North Carolina, Georgia, Florida, Louisiana, Texas, US; Laguna Madre in Tamaulipas, Mexico; Colombia. | Watling & Maurer, 1972; Bousfield, 1973; Thomas, 1976; Heard, 1982; Nelson, 1995; Camp et al., 1998; Ortiz et al., 2007; Miloslavich et al., 2010; Martín et al., 2013 |
| Apohyale prevostiiFigure 7 Labrador, Gulf of St Lawrence, Nova Scotia, Canada; Maine, New Hampshire, Massachusetts, Rhode Island, Connecticut, US; Laguna Madre in Tamaulipas, Laguna de Tamiahua in Veracruz, Mexico; Bermuda; Pernambuco, Brazil. – Arctic Ocean; Iceland; Ireland; United Kingdom; Faroe Islands; Norway; Netherlands; Belgium; France; Spain. | Bousfield, 1973; Lincoln, 1979; Wakabara & Serejo, 1998; Chavanich & Wilson, 2000; Barba & Sánchez, 2005; Lowry, 2014 |
| Jassa falcata + Figure 7 Newfoundland and Nova Scotia, Canada; Maine, New Hampshire, Massachusetts, Rhode Island, Connecticut, New York, New Jersey, Delaware Bay, Chesapeake Bay, Virginia, North Carolina, Georgia, Florida, Texas, US; Laguna Madre in Tamaulipas, Mexico; São Paulo, Brazil; Group A: 36°–43°S, Argentina. – Ireland; United Kingdom; Norway; Spain; Mediterranean Sea; British Columbia, Canada to California, US; South China Sea; New Zealand. | Barnard, 1972; Watling & Maurer, 1972; Bousfield, 1973; Dickinson et al., 1980; Leite et al., 1980; Barnard & Karaman, 1991; Wakabara et al., 1991; Nelson, 1995; Camp et al., 1998; Wakabara & Serejo, 1998; Lowry, 2000; Barba & Sánchez, 2005; López Gappa et al., 2006; Ortiz et al., 2007; Cerrato et al., 2010; C |
| Orchestia gammarellus + Figure 7 Newfoundland to Cape Breton Island, Nova Scotia, Bay of Fundy, Canada to Maine, New Hampshire, Massachusetts, Rhode Island, Connecticut, US; Laguna Madre in Tamaulipas, Mexico; Pernambuco, Brazil; Group B: 36°–58°S, Argentina. – Iceland; Norway; United Kingdom; Spain; Mediterranean Sea; Black Sea, Turkey; southwest Africa. | Bousfield, 1973; Soares, 1979; Wakabara & Serejo, 1998; Barba & Sánchez, 2005; López Gappa et al., 2006; Ortega, 2013; B, C |
Peer Review under the responsibility of Universidad Nacional Autónoma de México.