




Calliphoridae flies allow estimating the postmortem interval (PMI), as they are the first to arrive at a decomposing body. Its larvae feed on the carcass and then move away to pupate (post-feeding dispersal). Since the environment alters the physiology and behaviour of insects, the dispersal of larvae and the final location of pupae could be related to the place where the carcass is deposited. The aim of this work was to study the dispersal of post-feeding larvae in relation to environmental variables in northern Patagonia.
Materials and methodsThe distribution of pupae in response to different illumination, temperature, and soil inclination conditions was evaluated. A circular box divided into quadrants was used for each treatment and a control. Post-feeding larvae were placed in the centre of each box, allowing them to disperse until they pupate. Differences in the number of pupae collected in each quadrant were evaluated by ×2 tests.
ResultsAccording to the control, the dispersion of post-feeding larvae occurs randomly (p=.098). Regarding temperature, the larvae eluded the heat source (p<.01). The inclination treatments showed that larvae avoided the raised surface (p<.01). Concerning illumination conditions, no trend was observed (p=.41).
ConclusionsKnowing the dispersal patterns of larvae, considering environmental variables, reduces search times and let infers the sectors with the highest probability of finding pupae. In this way, it is likely to estimate a more precise PMI in the framework of medico-legal investigations.
Las moscas de Calliphoridae permiten estimar el intervalo postmortem (IPM), por ser las primeras en arribar a un cuerpo en descomposición. Sus larvas se alimentan del cadáver y luego se alejan para empupar (dispersión postalimentaria). Dado que el ambiente altera la fisiología y el comportamiento de los insectos, la dispersión de larvas y la ubicación final de las pupas podrían relacionarse con el lugar donde se deposita el cadáver. El objetivo de este trabajo fue estudiar la dispersión de larvas postalimentarias en relación a variables ambientales en norpatagonia.
Materiales y métodosSe evaluó la distribución de pupas en respuesta a condiciones de iluminación, temperatura e inclinación del suelo. Se utilizó una caja circular dividida en cuadrantes para cada tratamiento y un control. En el centro de cada caja se colocaron larvas postalimentarias, permitiendo que se dispersen hasta empupar. Las diferencias en la cantidad de pupas colectadas en cada cuadrante fueron evaluadas mediante pruebas de X2.
ResultadosSegún el control, la dispersión de larvas postalimentarias se produce de manera azarosa (p = 0,098). Con respecto a la temperatura, las larvas eludieron la fuente de calor (p < 0,01). En cuanto a la inclinación, las larvas evitaron la superficie elevada (p < 0,01). Respecto a la iluminación, no se observó una tendencia (p = 0,41).
ConclusionesConocer los patrones de dispersión de larvas, considerando variables ambientales, permite disminuir los tiempos de búsqueda e inferir los sectores con mayor probabilidad de encontrar pupas. De esta manera, es posible estimar un IPM más preciso en el marco de investigaciones médico-legales.
Forensic entomology, for medical–forensic purposes, studies insects and other arthropods that arrive on a corpse.1 The primary aim of this discipline is to study the time elapsed since the time of death and the discovery of the body, otherwise known as the postmortem interval (PMI).
To this end, it relies on 2 methods, the sequence of different species and the estimation of the age of the insects. The first is based on the known order of appearance of the entomofauna on a decomposing body.2 The second method involves determining the age of the various developmental stages of the insects, particularly in the case of the oldest individuals.3 Thus, with the information available concerning the development of insects that are forensically relevant, their age can be calculated and from that, the time at which the insects colonised the body at the onset of decomposition can be inferred.
Flies (Order: Diptera) are the first insects to colonise and lay eggs on a corpse. Once the eggs have matured, the larvae hatch and then consume the decomposing body tissues. Sometime thereafter, they begin to abandon the substrate in a process known as post-feeding dispersal,4 during which they search for a suitable site to proceed to the next stage, the pupal stage. During this last stage, pupae bury themselves and remain immobile until metamorphosis is complete and, finally, the adult emerges.5 The final arrangement of the pupae will therefore depend on where the larvae prefer to go once they leave the cadaver. Understanding post-feeding larval dispersal can assist in estimating the MPI, not least because this time frame can be underestimated if the more advanced stages are not collected.
Because of the ectothermy characteristic of insects, larval dispersal is influenced by a number of environmental variables that influence their physiology and behaviour.6 Earlier studies have explored post-feeding larval dispersal and pupal development.7 For instance, Robinson et al.8 looked how different indoor surfaces impact the dispersal process and larval development. They suggest that uncarpeted environments, in contrast to carpeted surroundings, result in delayed pupation, which is likely due to the fact that larvae require greater energy expenditure during the dispersal process. Singh and Bala9 conducted a study on the dispersal distance of Calliphorid larvae (Diptera: Calliphoridae) relative to the weight of each pupa, and found that lighter larvae can cover greater distances. Recently, several authors have suggested that during the feeding and dispersal stage, larvae remain clustered together, even among individuals of different species.10–12
On the other hand, only a few studies have investigated the final distribution of pupae taking into account environmental variables. Tessmer and Meek13 evaluated the spatial distribution of calliphorid pupae under field conditions in the southern United States and reported differences between seasons and soil types. Nevertheless, the effect of specific environmental variables on the final disposition of larvae once they leave the corpse remains unknown. Hence, the this work seeks to examine post-feeding dispersal of larvae as a function of temperature, lighting, and slope of the surface underneath the body, with the hypothesis that larvae disperse in a cluster, depending on environmental variables, thereby leading to a distinctive pattern of pupal location. The ultimate goal of this study is to provide relevant information that can optimise the collection of entomological evidence at the crime scene by forensic investigators and increase the precision of the MPI estimation.
Materials and methodsTo study post-feeding dispersal, larvae in the dispersal phase were tested, as well as the final distribution of pupae. Using baited traps, adults of Calliphoridae species (Order: Diptera) were collected in the city of Cipolletti, Río Negro province, Argentina (38° S, 68° W), in February 2018. After the flies oviposited in the traps, approximately 300 eggs were collected per treatment and placed in a 300 ml container with 200 g of beef. This ensured a sample size of at least 50 post-feeding larvae for each experiment. All the larvae that hatched and matured to the dispersal stage were placed in the centre of a circular box filled with soil, where they were allowed to disperse freely. After 1 week, all the pupae were collected. Treatments and control were conducted in parallel, better 19 and 26 February 2018. The response variable was the number of pupae collected per quadrant.
The box protocol for each test consisted of a circular expanded polystyrene base with a diameter of 50 cm, divided into 4 quadrants (C1, C2, C3, and C4). The divisions were made with acetate sheets. Natural soil with no added chemicals was used as substrate to avoid altering the treatment conditions by unknown variables (Fig. 1). To evaluate the effect of environmental variables on the dispersal of post-feeding larvae, four boxes were used; 1 control box and 3 boxes with the following treatments: lighting, temperature, and the slant of the soil (Fig. 2).

 and B) temperature treatment, C) and D) tilt treatment, E) lighting treatment, and F) control box.")
For the temperature treatment condition, the temperature in C1 was increased by 5 °C relative to the remaining quadrants. A halogen lamp with a thermostat was used, covered with aluminium foil to prevent the box from being influenced by light (Fig. 2A and B). The temperature recorded on the day of the experiment was 25 °C, so the set cut-off temperature for the thermostat was 30 °C. As for the slope, the treatment consisted of raising the C1 box, until a 20° slope with respect to the other quadrants (Fig. 2C and D) was achieved. As for lighting treatment, C1 was illuminated, leaving the other quadrants in total darkness, covered with a cardboard lid (Fig. 2E). A white LED spotlight was used to ensure that no heat was generated. Finally, no modifications were made to the control box (Fig. 2F).
Data analysisDifferences in the number of pupae per quadrant for each of the treatment conditions were analysed using a goodness-of-fit test X2.14 The null hypothesis was that the data conform to a uniform pattern of distribution, i.e., pupae are found in similar numbers in all quadrants of the box.
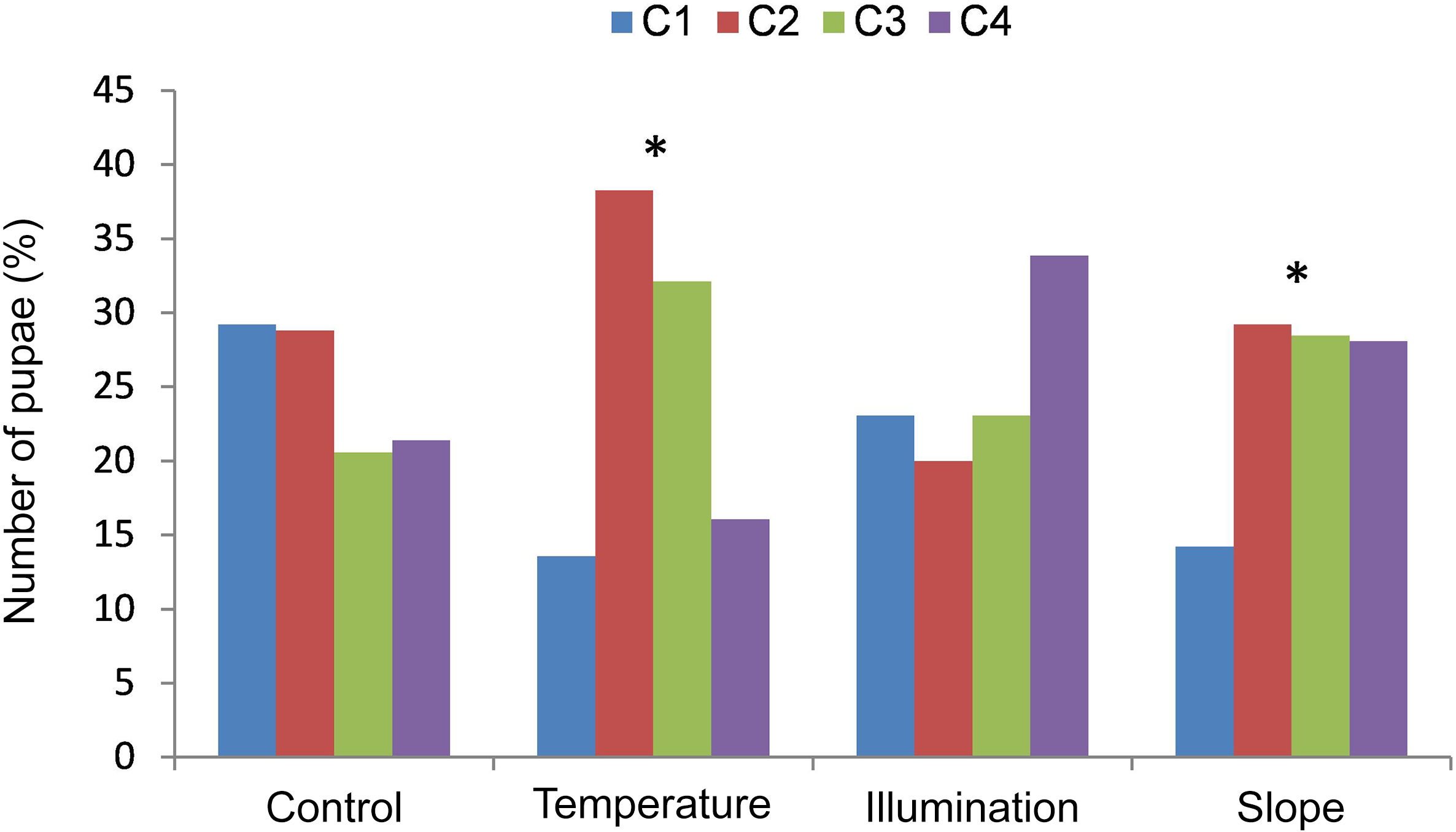
ResultsControlA total of 243 pupae were collected in the control box, distributed as depicted in Fig. 3. After allowing for random dispersion, significant differences were detected between the actual dispersion pattern and the one expected in the quadrants of the control box (p=.098).
 in the treatment boxes. *Significant differences between the frequencies observed and the ones expected. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)")
Relative frequency of pupae per quadrant in the control box and in the various treatment conditions. The variable is applied to quadrant 1 (C1, blue bar) in the treatment boxes. *Significant differences between the frequencies observed and the ones expected. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
All-in-all, 81 pupae were collected (Fig. 3). Significant differences were noted between the observed and expected frequencies. Larvae mostly chose quadrants 2 and 3 (p<.01). There were fewer pupae in both C1 and C4, which could be attributable to an effect caused by the heat source on the adjacent quadrant. This would also explain the higher number of pupae in C2 and C3.
SlopeA total of 267 pupae were collected (Fig. 3). The results reveal significant differences between the number of pupae observed in C1 and those in the other quadrants (p<.01).
LightingAll together, 65 pupae were collected and distributed as per Fig. 3. The results obtained failed to evidence significant differences in pupae distribution in the various quadrants (p<.41).
DiscussionThe effect of environmental factors on the dispersal of post-feeding larvae was studied in the Norpatagonian region, Argentina, where there is no history of this type of study. We found that both temperature and soil slope effect the final distribution of pupae. On the other hand, lighting made no significant differences. In addition, we did not detect any pattern of larval dispersal under control conditions.
The analysis of larval dispersion with respect to temperature indicated that larvae prefer less warm quadrants. This distribution may be a consequence of an avoidant behaviour on the part of the larvae to prevent possible dehydration, given that the cut-off temperature set for the lamp in the test was 30 °C. Based on earlier work, the optimal temperature for development (i.e., the ratio of growth rate to survival) of many species of Calliphoridae is between 20 and 25 °C.15–19 For instance, Diaz Martín et al.17 analysed the influence of temperature on Calliphora vicina (Diptera: Calliphoridae) under controlled conditions. The authors found that from 30 °C onwards, development slows and the larvae do not complete their development, dying several weeks after they begin to migrate and becoming dehydrated due to the heat prior to reaching the pupal stage. However, inasmuch as the optimum temperature may vary from one species to another, future trials should factor this variable in. In any event, an experimental design that explores the behaviour of the species assemblage makes it possible for the results to be a more representative reflection of the diversity of entomofauna collected at the scene of an event.13
The results of the slope experiment suggest that larvae avoided the quadrant with the steepest slope. This could be interpreted as a way to decrease the energy expenditure of the larvae prior to burial and to avoid reducing their reserve, which would be a benefit at the time of pupation. Consistent with this, Sharma et al.20 analysed burial depth under 3 controlled conditions (5, 25, and 45 cm), and found more pupae within the first 5 cm of soil in both upward and downward dispersal. Furthermore, they discovered that adult emergence decreases as depth increases. Still, there is a paucity of information concerning the effect of slope on the process of post-feeding dispersal. Therefore, such tests should be repeated in the future, using varying degrees of slope so as to cover a broader range of situations compatible with actual practice. Current protocols dictate that soil samples be randomly collected around the body. Thus, by complementing the data from the work by Sharma et al.20 with those generated in the present study, extremely useful information is obtained as to where and at what depth to look for entomological evidence at the scene, bearing in mind the topographical characteristics of the site.
The analysis of the dispersal of larvae with changes in light revealed that there was no preference for brighter or darker areas. In this case, larval dispersal behaviour appeared to be unaffected by the stimulus of artificial light under controlled conditions. Kocarek21 investigated the dispersal of post-feeding larvae of Calliphora vomitoria and Lucilia caesar under natural conditions and found that they dispersed exclusively at night, thereby minimising interactions with daytime and twilight predators such as parasitoid insects, and insectivorous birds or mammals. This apparent discrepancy with our findings may be due to differences in experimental conditions. Given that Kocarek21 performed his tests under natural conditions, it is impossible to discriminate whether the effect depends on illumination or temperature. Consequently, dispersal at night might also stem from a preference for dispersal at lower temperatures. Based on our results, the lighting of a site should not be taken into account when collecting entomological traces.
The generation of knowledge about the process of postmortem dispersal contributes substantially to the development of protocols for action. Such protocols should take into account as many variables as possible that might come into play when collecting evidence at the scene of a suspected crime. Being familiar with dispersal patterns based on certain variables enables investigators to reduce search times and to infer more precisely the areas around a corpse where postmortem larvae or pupae are most likely to be found. The PMI may be underestimated if only larvae are lifted from the body, or if pupae or dispersing larvae are not collected because of lack of time or because the wrong places are searched. Therefore, the study of Calliphoridae larval dispersal has far-reaching implications for medico-criminal investigations.
FundingThis work has been funded by the National Interuniversity Council [Grant No. CIN EVC3-UNRN1925] and the National University of Río Negro [PI (2015) 40-A-463].
Please cite this article as: Lavezzo JC, Pereira AJ, Béguelin M. Spatial dispersion of post-feeding larvae of forensically important flies: Implications for medicolegal investigation. Revista Española de Medicina Legal. 2024. https://doi.org/10.1016/j.remle.2023.09.002.