La magnetoencefalografía es una técnica de neuroimagen no invasiva que mide, con gran exactitud temporal, los campos magnéticos en la superficie de la cabeza producidos por corrientes neuronales en regiones cerebrales. Esta técnica es sumamente útil en la investigación básica y clínica, porque además permite ubicar el origen de la actividad neural en el cerebro. En esta revisión se abordan aspectos básicos de la biofísica del método y se discuten los hallazgos sobre procesos como la percepción del habla, la atención auditiva y la integración de la información visual y auditiva, que son importantes en la investigación. Igualmente, se ilustran sus ventajas, sus limitaciones y las nuevas tendencias en la investigación con magnetoencefalografía.
Magnetoencephalography is a noninvasive imaging technique that measures the magnetic fields on the surface of the head --produced by neuronal currents in brain regions -- and provides highly accurate temporal information. Magnetoencephalography is extremely useful in basic and clinical research as it can also locate the sources of neural activity in the brain. This review chiefly approaches biophysics-related aspects of the method; findings are also discussed on issues such as speech perception, auditory attention and integration of visual-auditory information, which are quintessential in this type of research. Lastly, this review discusses the benefits and limitations of magnetoencephalography and outlines new trends in research with this technique.
El cerebro humano es un sistema biológico extremadamente complejo que ejecuta funciones sensoriales, motoras, lingüísticas y cognoscitivas mediante procesos neuronales. Por siglos, científicos, médicos y filósofos han mostrado gran interés en estudiar la relación existente entre la actividad cerebral y los eventos mentales, han desarrollado teorías y han hecho descubrimientos fascinantes. Sin embargo, comprender la dinámica espaciotemporal de la actividad cerebral se mantiene como uno de los retos más intrigantes en la ciencia: ¿cómo trabaja el cerebro?, ¿cómo se desarrollan las habilidades?, ¿qué subyace a las fallas sensoriales, motoras, cognoscitivas y comunicativas?, ¿qué métodos pueden aplicarse para la prevención y el tratamiento de ciertos trastornos?
La complejidad de la actividad cerebral involucra una red de billones de neuronas que se comunican entre ellas vía trillones de conexiones sinápticas. Los trabajos de la red neuronal dependen considerablemente de procesos eléctricos mediados químicamente, que pueden medirse en línea usando técnicas invasivas o no invasivas. La magnetoencefalografía (MEG) es una técnica de neuroimagen no invasiva que mide pequeños campos magnéticos producidos por corrientes neuronales en diversas regiones cerebrales (Hämäläinen, Hari, Ilmoniemi, Knuutila & Lounasmaa, 1993). La MEG es una herramienta esencial de investigación para estudiar el cerebro y la cognición, en la que la información temporal de la actividad cerebral evocada es crítica. Otra ventaja es que permite estudiar a bebés que aún no son capaces de dar respuestas conductuales. Las señales MEG se afectan poco por las diferencias anatómicas individuales, tales como la conductividad del tejido. Cuando se combina la señal del MEG con la imagen de resonancia magnética estructural, la MEG proporciona la localización de la actividad cerebral con alta confiabilidad espacial (e.g., una resolución de unos cuantos milímetros; véase Barkley, 2004; para una discusión de las ventajas y desventajas relativas del método, véase Baumgartner, 2004). Sin embargo, la mayor limitación para el uso de la MEG es su alto costo.
Considerando su relevancia para el desarrollo de la neurociencia, en esta revisión se abordan aspectos teóricos básicos, de método y aplicaciones de la MEG, y además se discuten algunos hallazgos de la investigación para ilustrar sus ventajas y limitaciones en la visualización de la representación funcional y la organización de la actividad cerebral.
La señal MEG y su mediciónUna sola neurona en el cerebro trabaja como una pequeña bomba electroquímica con iones cargados eléctricamente que se mueven dentro y fuera de la membrana celular (Kandel, Schwartz, Jessell, Siegelbaum & Hudspeth, 2012). La estimulación de una neurona causa que un flujo de iones cargados positivamente entre a la célula y genere una despolarización. Si la despolarización tiene fuerza suficiente, genera un potencial de acción que causa que la neurona “dispare”. El potencial se propaga a través de la membrana celular hasta la región presináptica, donde libera al espacio sináptico neurotransmisores que llegan hasta la membrana de la célula posináptica e inducen cambios del flujo iónico dentro y fuera de la célula.
Cuando múltiples sinapsis excitatorias con orientaciones similares, como las encontradas en la capa piramidal de la corteza, se activan simultáneamente, su corriente neta converge en las dendritas apicales y fluye hacia el cuerpo celular perpendicular a la superficie de la corteza. Estas corrientes eléctricas se acompañan de un campo magnético. Se necesitan aproximadamente 50.000 neuronas activas para generar un campo magnético medible fuera del cuero cabelludo. Las sinapsis inhibitorias también pueden producir señales magnéticas medibles, aunque estas señales suelen ser más débiles que las producidas por las sinapsis excitatorias. Cuando las corrientes fluyen en direcciones opuestas, los campos magnéticos se cancelan (Hari & Salmelin, 2012).
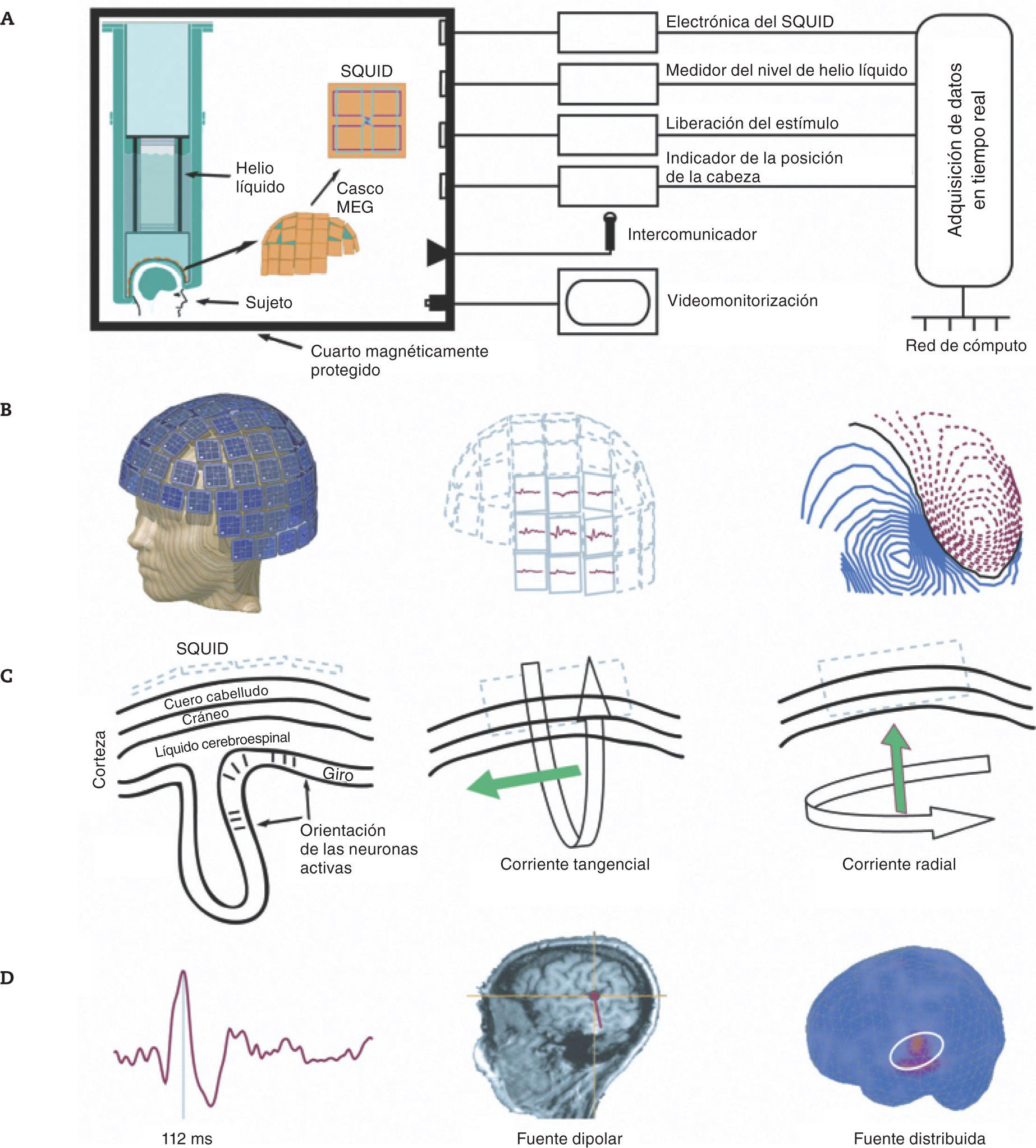
Las corrientes netas se modelan como corrientes dipolares con posición, orientación y magnitud definidas, pero no en extensión espacial (Hämäläinen et al., 1993). Una corriente dipolar da origen a un campo magnético alrededor del eje del vector de corriente. El campo magnético está orientado perpendicularmente al flujo de la corriente eléctrica. Debido a las orientaciones físicas de las neuronas, las bobinas colectoras de los dispositivos de interferencia cuántica superconductora (superconducting quantum interference devices [SQUID]) en un sistema MEG (Zimmerman, Thiene & Harding, 1970), son más sensibles a la actividad en los surcos cerebrales (figs. 1A-C). Los sistemas modernos de registro contienen un arreglo de más de 300 sensores SQUID organizados en forma de casco, que tienen una separación del cuero cabelludo de aproximadamente 2cm (Hari & Salmelin, 2012). Dada la distancia entre un área cerebral y las bobinas colectoras, la MEG es menos precisa para la medición de las fuentes de las estructuras subcorticales que para las de la corteza cerebral.
. C: representación esquemática de las medidas del MEG en relación con la orientación de las células piramidales en la corteza; solo los componentes tangenciales de una corriente neuronal generan campos magnéticos medibles por el SQUID. D: muestra de ondas MEG de respuestas evocadas auditivas (100 épocas promediadas para la ventana temporal –100–700ms relativa al inicio del estímulo), y localización del componente seleccionado calculado como fuente dipolar usando un modelo esférico de la cabeza basado en las imágenes de resonancia magnética del propio sujeto y como fuente distribuida sobre el cerebro estándar. MEG: magnetoencefalografía.")
A: diagrama de bloques de un sistema típico de MEG. El arreglo de sensores SQUID está bañado en helio líquido. B: casco del MEG sobre un modelo, una muestra de la distribución de la respuesta auditiva evocada y sus mapas de contorno, que muestran un patrón bipolar (las líneas continuas indican el flujo magnético dentro del cerebro y las líneas punteadas, el flujo magnético hacia fuera del cerebro). C: representación esquemática de las medidas del MEG en relación con la orientación de las células piramidales en la corteza; solo los componentes tangenciales de una corriente neuronal generan campos magnéticos medibles por el SQUID. D: muestra de ondas MEG de respuestas evocadas auditivas (100 épocas promediadas para la ventana temporal –100–700ms relativa al inicio del estímulo), y localización del componente seleccionado calculado como fuente dipolar usando un modelo esférico de la cabeza basado en las imágenes de resonancia magnética del propio sujeto y como fuente distribuida sobre el cerebro estándar. MEG: magnetoencefalografía.
Debido a la amplitud extremadamente pequeña de la señal, los sistemas MEG requieren la instalación de un cuarto protegido para evitar la interferencia de las fuertes señales magnéticas del ambiente. Los sensores SQUID están en una gran unidad de enfriamiento bañados en helio líquido a –269°C. A esta temperatura, los pequeños cambios en el campo magnético generados por las neuronas a pocos centímetros de los sensores pueden inducir un flujo de corriente en la bobina colectora.
El campo magnético evocado dentro de la cabeza tiene una amplitud de 100 fT (1 fT = 10−15 T), que es una milmillonésima parte de la fuerza del campo magnético de la Tierra (Tobimatsu, 2005). Para excluir el ruido de fondo, se mide el gradiente del campo magnético en vez del campo mismo. El gradiente se refiere a la diferencia en las mediciones del campo entre dos lugares diferentes, uno cerca del cuero cabelludo y el otro a cierta distancia de este. El gradiente decae rápidamente con la distancia. Dado que la fuente de fondo está muy lejana comparada con la del cerebro, su influencia en la señal MEG es sumamente reducida en estos registros. Para reducir ruidos de alta y baja frecuencia, se emplean típicamente el aluminio y el mu-metal respectivamente, junto con algoritmos activos de cancelación de ruido (Hari, Levänen & Raij, 2000).
Adicionalmente hay que considerar que las señales de interés están embebidas en actividad cerebral independiente del experimento. Esta actividad de fondo es más fuerte que la señal objetivo. Una solución sencilla es promediar un gran número de respuestas sincronizadas con la presentación de estímulos idénticos. Esto reduce la actividad de fondo, que es variable e independiente de la tarea experimental (Matani, Naruse, Terazono, Fujimaki & Murata, 2011). Hay que considerar también la dificultad del equipo para detectar corrientes radiales con una fuente en un giro de la corteza (fig. 1C). Una tercera restricción es que los sujetos deben permanecer quietos durante la sesión de registro. Los artefactos relacionados con el movimiento pueden afectar considerablemente a la calidad de la medición y hacer difícil el análisis de estimación de fuentes (Hari et al., 2000).
Los datos obtenidos con MEG no dan un indicador directo sobre la localización de la(s) fuente(s) de la actividad cerebral, pero se puede intentar determinarlas utilizando algunos modelos. Se conoce como solución al problema inverso cuando se encuentran la(s) fuente(s) de la actividad neural registrada en su componente magnético o eléctrico (Baillet, Mosher & Leahy, 2001). El problema inverso es lo opuesto al problema “anterógrado”, donde la fuente es conocida y la solución es la modelación del campo medido a una distancia dada de la fuente. En teoría, un mapa MEG registrado puede proceder de muchas configuraciones de fuentes de corriente. Así, podría haber una infinidad de soluciones potencialmente “correctas” al problema inverso. El desafío es determinar qué fuente o configuración de ellas es fisiológicamente plausible y qué solución podría proveer el mayor ajuste estadístico entre los valores predichos del modelo de fuente y las señales observadas (Hari et al., 2000).
Las soluciones no representan una certeza del origen de una fuente, sino una probabilidad estadística de dónde pueda encontrarse. El procedimiento es minimizar el número de posibles soluciones al implementar supuestos que permiten limitar las soluciones a estimaciones razonables de la configuración de la fuente (es decir, número de fuentes, ubicación de estas en el cerebro, dirección de la fuente y su fuerza). Dos aproximaciones generales en la estimación de fuentes permiten obtener una única solución: el modelo de dipolos y el modelo de fuentes distribuidas (fig. 1D).
Modelo de dipolosSe modela una fuente puntual (dipolo de corriente equivalente, equivalent current dipole [ECD]) o un conjunto de estas en cualquier instante y en cualquier sitio. Se identifica un instante a modelar y se presupone la ubicación de la(s) fuente(s). Después de la primera iteración, se genera el campo magnético resultante y se reajusta la ubicación para reducir la diferencia entre el campo estimado y el campo medido. Las iteraciones llegan hasta la convergencia con el mínimo error residual. Un método alternativo consiste en realizar un análisis de componentes independientes (independent component analysis [ICA]) que permite discriminar las fuentes sin usar el modelo anterógrado porque puede localizar las fuentes individualmente. Este método puede mejorar la relación señal-ruido al separar correctamente la señal de las fuentes neurales del ruido de fuentes no neurales.
Las sucesivas muestras en el tiempo se modelan como dipolos móviles (Breuer, Dammers, Roberts & Shah, 2013). Los mapas del campo magnético suelen deberse a fuentes que involucran varias regiones corticales simultáneamente; la estimación de las fuentes puede basarse en dipolos separados espacialmente, pero implica una mayor demanda computacional. Los efectos de las fuentes conocidas pueden quitarse antes de identificar una nueva fuente. Considérese que los dipolos en el tejido neural solo existen en una escala microscópica y la exactitud de la configuración en la corteza no puede determinarse, dados los 3cm de distancia que hay entre la fuente y los sensores de medición (por simplicidad, los patrones de activación focal < 2cm de largo se aproximan a un dipolo; Hari et al., 2000).
Para localizar la fuente, es necesario tener un modelo válido de conductividad macroscópica del tejido cerebral y de la cabeza. Por simplicidad, se emplea el modelo conductor simétrico esférico, que consiste en especificar esferas concéntricas con diferentes conductividades: una para el cuero cabelludo, otra para las caras externa e interna del cráneo y otra para el cerebro. Los cálculos basados en modelos esféricos son bastante precisos. Para una aproximación más exacta, es necesario considerar un modelo realista del cerebro. La geometría realista puede basarse tanto en imágenes de resonancia magnética de un individuo como en una imagen del cerebro estandarizado escalado individualmente.
El modelo del elemento frontera (boundary element model [BEM]) usa un sistema de compartimentos homogéneos y requiere una discretización (aproximada por múltiples figuras triangulares) (fig. 1D) de la superficie de los compartimentos del volumen conductor: el cerebro. Para mejorar el modelo, se puede usar la teselización por medio del procedimiento del elemento finito (finite element model [FEM]), que tiene en cuenta detalles finos de la conductividad del tejido cerebral. A diferencia del BEM, en el FEM se puede asignar una conductividad diferente a cada región sólida reconstruida por múltiples tetraedros. Sin embargo, las restricciones adicionales y las especificaciones en los modelos de la cabeza basados en geometría real incrementan la demanda computacional (Hämäläinen et al., 1993).
Modelo de fuentes distribuidasAun cuando los modelos de dipolo son útiles para modelar generadores de las respuestas auditivas, táctiles y visuales, estos proveen una cruda aproximación de la actividad cerebral subyacente, por lo que se han propuesto modelos que no requieren información a priori o algún supuesto específico acerca de la configuración de la fuente para calcular distribuciones tridimensionales de múltiples fuentes. En esta se supone que la actividad cerebral se dispersa sobre todas las posibles localizaciones de las fuentes para un modelo de cabeza dado. Un ejemplo de esta aproximación es la estimación de corriente mínima (minimum current estimates [MCE]; e.g., el método de mínima norma) que presenta una solución distribuida de corriente cuyo máximo supuesto para una única solución es que la suma total de corriente sea la menor para explicar las señales medidas (Uutela, Hämäläinen & Somersalo, 1999). Una aproximación diferente, la magnetometría de apertura sintética (synthetic aperture magnetometry [SAM]), usa una ponderación lineal de los sensores para centrar el arreglo en un lugar dado (Robinson & Vrba, 1999).
Es difícil determinar cómo la extensión de la propagación y la dispersión de la actividad puede capturarse en el análisis de fuentes distribuidas, ya que el supuesto es que la actividad está siempre distribuida (para una aproximación de aglomeración de dipolos en la estimación de la duración y la extensión de las actividades MEG localizadas, véase Zhang, Kuhl, Imada, Kotani & Tohkura, 2005). En general, con el análisis de fuentes distribuidas, el ruido se confunde más fácilmente con las fuentes reales (comparado con el análisis de dipolos).
Aplicaciones de la MEG en la investigación básica y clínicaEl cerebro humano tiene regiones especializadas en el procesamiento de diversos estímulos sensoriales, lingüísticos, cognoscitivos y para el control motor. La comprensión adecuada de las funciones cerebrales requiere la localización y la temporalidad de la activación cerebral. Se ha mostrado una relación entre ventanas temporales y la actividad neural de fenómenos como los efectos de la experiencia lingüística en la percepción del habla (Hari et al., 2000; Hari & Salmelin, 2012) que se asocian a componentes de la respuesta evocada, tales como la respuesta N1m (componente negativo de la señal MEG que aparece aproximadamente 100ms después de la presentación del estímulo) y la respuesta magnética mismatch (MMNm; componente negativo en la señal del MEG, que aparece aproximadamente a los 100–250ms). Algunos de los hallazgos se resumen a continuación para ilustrar cómo los datos MEG permiten evaluar teorías y determinar los mecanismos cerebrales subyacentes de varios procesos cognitivos y conductuales, así como su utilidad en la evaluación clínica.
Percepción del hablaExisten dos teorías opuestas que explican la percepción de categorías fonéticas. Una postula la existencia de mecanismos especializados del habla, existentes desde el nacimiento y modificables mediante procesos de selección. La otra, considera que la percepción del habla surge de las habilidades auditivas y cognitivas generales humanas generadas por los principios del aprendizaje. Algunos modelos animales (modelos en primates no humanos y en otros animales) indican que el sistema auditivo de los mamíferos incorpora mecanismos básicos involucrados en la percepción categórica del habla (Steinschneider, Nourski & Fishman, 2013).
El intervalo de inicio de la sonoridad (voice onset time [VOT]) es una pista acústica efectiva que marca la diferencia entre sílabas que inician con una consonante oclusiva sonora y aquellas que inician con una oclusiva sorda. El VOT es un parámetro fonético útil para dilucidar los eventos neurales asociados con la percepción categórica. Steinschneider et al. (2013) mencionan que algunas habilidades de los animales para discriminar consonantes que difieren en el VOT son similares a las humanas.
En estudios con humanos donde se presentan cambios paulatinos que permiten ir del fonema /b/ al /p/, lo que permite estudiar el efecto de frontera fonética que se cree es una consecuencia de grupos específicos de neuronas (Capilla, Belin & Gross, 2013), se encuentra una reducción en la amplitud de N1m en correspondencia con la percepción del cambio categórico. Esta reducción también se extiende a estímulos que no son habla, lo que confirma la fuerte base auditiva de la partición categórica para estos contrastes. Sin embargo, estos datos no llegan a mostrar poblaciones neuronales separadas a los contrastes de vocalización ni a explicar los bien conocidos efectos de la experiencia en el lenguaje.
Contra la postura de la percepción del habla que surge del proceso general auditivo, algunos estudios de MEG muestran propiedades únicas para el procesamiento del habla (Imada et al., 2006). Por ejemplo, se encontró que los cambios en las categorías fonéticas (/a, i, u/) causan cambios significativos de latencia en N1m, sin que esto ocurra en cambios en la frecuencia fundamental de los estímulos (Poeppel et al., 1997). En otro estudio, Phillips et al. (2000) registraron la MMNm a múltiples estímulos auditivos frecuentes e infrecuentes escogidos de un continuo sintético /da-ta/. Para una “condición fonológica”, se usaron múltiples tipos de la misma categoría, mientras en la condición “control acústica” se eliminó el cambio categórico definido entre los frecuentes e infrecuentes manteniendo la misma separación física.
Se encontraron MMNm significativas en la condición fonológica pero no en la acústica, lo que proporciona evidencia de un nivel abstracto de agrupamiento fonológico diferente del simple procesamiento auditivo. Näätänen et al. (1997) examinaron la representación cortical de fonemas específicos del lenguaje y encontraron que la MMN para la detección de un cambio en las vocales nativas era mayor en la corteza auditiva izquierda que en la derecha. Las vocales no nativas tuvieron respuestas bilateralmente equivalentes.
Zhang et al. (2005) usaron un diseño similar y encontraron una gran variabilidad individual con respecto a la lateralidad de la representación del habla. Según Näätänen (2001), existen procesos en paralelo que contribuyen a la MMN en la discriminación del habla. Uno es el proceso de detección del cambio acústico con la activación bilateral y el otro, el procesamiento fonético específico, el cual está lateralizado a la corteza auditiva izquierda. Así, el aprendizaje fonético de un segundo lenguaje, como hablantes de japonés en entrenamiento para aprender el contraste /r-l/ en inglés, muestra un cambio más prominente en el hemisferio izquierdo que en el derecho (Zhang, Kuhl, Imada, Kotani & Pruitt, 2001). Sin embargo, Kasai et al. (2001) reportaron que la MMNm de los japoneses a los tonos puros y a los cambios intracategóricos de una vocal fue predominante en el hemisferio derecho, mientras que la MMN a los cambios en las categorías de vocales fueron bilaterales.
Atención auditiva y visualLos estudios con MEG muestran que la atención auditiva modula la actividad cortical muy temprano en el procesamiento, de 20 a 40ms a partir del inicio del sonido, lo que refleja el rápido análisis de las características en la entrada para su posterior procesamiento (Hari et al., 1989). Se ha reportado que diferentes señales MEG se asocian con tareas auditivas de diferentes demandas atencionales, lo que provee información sobre la actividad en un circuito local y la integración entre regiones (Kaiser & Lutzenberger, 2005).
En un estudio realizado por Xiang, Simon y Elhilali (2010), los participantes atendían a una o dos secuencias de tonos intentando detectar desviaciones en el flujo lento (4Hz) o rápido (7Hz). Los resultados mostraron un aumento en la señal neural a la repetición del blanco atendido. La fuente se localizó en la corteza auditiva, en la circunvolución de Heschl y el giro temporal superior, lo que indica que se dan respuestas en esta región ante las demandas atencionales.
En la modalidad visual, la investigación previa en animales y humanos muestra que la corteza parietal y frontal está involucrada en la atención alternante y en el movimiento ocular para enfocar ciertos objetos en el campo visual (Corbetta, 1998). Vanni y Uutela (2000) usaron el modelo de estimación de corriente mínima para localizar las fuentes en un estudio con MEG, y encontraron claros efectos de modulación atencional en las regiones parietal y frontal. Vidal, Chaumon, O’Regany y Tallon-Baudry (2006) examinaron la hipótesis de que los patrones de oscilación de la sincronía neural son específicos de distintos procesos visuales en tareas de atención. Sus resultados mostraron oscilaciones relacionadas con la integración en la banda gamma alta (70–120Hz) en la región central occipital y oscilaciones relacionadas con la atención en la banda gamma baja (44–66Hz) en áreas parietales.
Integración audiovisualLa percepción del habla se puede facilitar con pistas visuales; por ejemplo, cuando se conversa con una persona en un ambiente ruidoso, la lectura de labios puede favorecer la comprensión. La investigación conductual (McGurk & MacDonald, 1976) ilustra que, cuando no hay relación entre la información visual de la articulación y el sonido escuchado, la información visual puede fusionarse en una señal auditiva y se puede percibir algo diferente del sonido presentado. Los datos obtenidos con MEG sobre el efecto McGurck indican que la entrada visual desde la observación de los movimientos de la boca sincronizados con la articulación del habla afecta al procesamiento del habla en la corteza auditiva aproximadamente 180ms después del inicio del estímulo (Sams et al., 1991). Incluso la información visual puede modular los juicios de la distancia de sonidos. Altmann et al. (2012) encontraron que las respuestas a N1m en la corteza auditiva izquierda fueron más fuertes ante las pistas visuales más distantes que ante las más próximas.
Neuronas “espejo”El descubrimiento de las neuronas “espejo” en monos sustenta teorías acerca del aprendizaje de la imitación y las bases neurobiológicas y evolutivas del lenguaje (Hauser, Chomsky & Fitch, 2002). En humanos adultos se piensa que la activación del área motora izquierda (área de Broca) refleja la codificación articulatoria y/o fonológica durante la percepción del habla. En monos, un área homóloga en la corteza premotora descarga durante el movimiento de la mano, así como durante la observación de una acción similar pero hecha por otro mono o el experimentador. A este sistema de neuronas que se dispara por ver a “otro” hacer la acción se lo llama neuronas espejo.
Estudios con MEG confirman la existencia de este sistema en humanos que involucra la corteza motora primaria bilateral, la corteza visual-occipital izquierda y al área de Broca (Hari et al., 1998). Según Rizzolatti y Arbib (1998), las neuronas espejo facilitan un “vínculo neural faltante” entre gestos orofaciales y manuales y el control del habla formando las bases gestuales y articulares en la evolución del lenguaje.
Un estudio con MEG en infantes examinó la hipótesis de neuronas espejo en una condición de escucha pasiva en el primer año de vida (Imada et al., 2006). Los autores midieron las respuestas del hemisferio izquierdo a tonos armónicos y sílabas en un grupo de neonatos, un grupo de infantes de 6 meses de edad y otro de 12. Se observaron activaciones del giro temporal superior en todas las edades para todos los estímulos. Mientras los neonatos no mostraron activación frontal inferior, los bebés de 6 meses tuvieron una actividad temprana para los armónicos y una actividad tardía para las sílabas, y los niños de 12 meses tuvieron una activación temprana para los tonos y armónicos, así como tardía para los estímulos armónicos y del habla.
Los patrones de activación proveen evidencia inicial para un vínculo perceptual motor emergente de la percepción del habla. Además, indican que las áreas motoras del habla no se activan por la percepción del habla inicialmente, sino que requieren experiencia para vincular percepción y acción.
Guía quirúrgica, mapeo y evaluación prequirúrgica y posquirúrgicaEl uso clínico de la MEG es para detección y localización de zonas epileptógenas en la evaluación prequirúrgica, y el mapeo de la corteza circundante elocuente para intervenciones en pacientes con tumores cerebrales o epilepsia intratable (Stufflebeam, Tanaka & Ahlfors, 2009). La corteza elocuente se refiere a las áreas intactas de la corteza que son críticas para el procesamiento sensorial, motor y lingüístico y, por lo tanto, no deben removerse quirúrgicamente. Las áreas de corteza elocuente son: para lenguaje, lóbulos frontales y temporal izquierdo; visión, lóbulos occipitales; sensación, lóbulos parietales, y movimiento, corteza motora. El uso de esta técnica en evaluaciones prequirúrgicas se relaciona con mayores posibilidades de recuperación posquirúrgica (Stufflebeam et al., 2009).
Como las lesiones cerebrales pueden alterar estructuralmente el tejido cerebral circundante al moverlo, presionarlo, por infiltración o edema, los signos fisiológicos de la función deteriorada en estas zonas se reflejan en ondas focales lentas y rápidas y actividad paroxística en la MEG. Otras posibles aplicaciones clínicas de la MEG incluyen la estimación no invasiva de la lateralización del lenguaje y el seguimiento de la plasticidad cerebral por lesiones neurales centrales o periféricas, o durante el tratamiento del dolor crónico (Breier, Maher, Novak & Papanicolaou, 2006).
Nuevas tendencias en la investigación con MEG y futuras implicacionesEn las neurociencias cognoscitivas, la norma es conocer qué áreas del cerebro están involucradas en una función cognoscitiva, cómo están conectadas a través de redes generales y cómo se reclutan diferentes sistemas neurales en los cerebros de individuos sanos y enfermos. Es clara la utilidad de la MEG para seguir la actividad cerebral en tiempo real en comparación con la resolución de la resonancia magnética funcional (Hari & Salmelin, 2012). A pesar del gran desarrollo en las últimas décadas, aún requiere esfuerzo integrar el conocimiento concerniente a la mente y el cerebro en términos de la especialización funcional y la integración. Teniendo en cuenta las características, los alcances y las limitaciones de la MEG, consideramos que la tendencia será el desarrollo en cinco grandes áreas:
- 1.
El desarrollo de un mejor procesamiento de señal y de técnicas de recolección de datos para encontrar la mejor estimación de las fuentes de corriente distribuidas en el cerebro (Breuer et al., 2013; Matani et al., 2011).
- 2.
El estudio de los ritmos cerebrales y su relación con eventos sensoriales, cognitivos y motores (Wang et al., 2012). Los estudios de los ritmos cerebrales abastecerán nuevas ideas sobre el papel de la actividad oscilatoria cerebral en atención, afecto, memoria y aprendizaje.
- 3.
En el desarrollo de la neuroimagen multimodal, la meta es colectar una variedad de datos para visualizar y desarrollar nuevos métodos y estrategias que combinen técnicas de imagen óptica, eléctrica, magnética y radiactiva (Toga, Thompson, Mori, Amunts & Zilles, 2006). La integración de MEG y resonancia magnética funcional facilita la identificación precisa de las áreas cerebrales activas y la identificación temporal precisa de las respuestas cerebrales. Los estudios en neuroimagen multimodal permitirán hallar similitudes y diferencias entre las medidas hemodinámicas y neurofisiológicas para una mejor comprensión de la representación neuronal de las funciones lingüística, afectiva, cognoscitiva y social (Iwaki, Bonmassar & Belliveau, 2013).
- 4.
Avance en el estudio del desarrollo humano. Estudios recientes han mostrado la mayor utilidad de la MEG comparada con otras técnicas para el estudio de infantes y fetos en el procesamiento somatosensorial (Rahkonen et al., 2013), la discriminación auditiva (Lutter, Maier & Wakai, 2006) y la percepción del habla (Imada et al., 2006). Próximos estudios serán capaces de mapear los periodos esenciales del desarrollo en la maduración cerebral que marcan los periodos críticos del aprendizaje infantil e investigar la plasticidad cerebral y los mecanismos del cambio en el desarrollo.
- 5.
Avance en la investigación sobre los trastornos cerebrales. Un número cada vez mayor de médicos e investigadores han comenzado a usar la MEG para estudiar una amplia gama de poblaciones de pacientes con daño cerebral, deficiencias y trastornos, entre otros, la epilepsia (Amo et al., 2005), el autismo (Muñoz-Yunta et al., 2006), trastornos del lenguaje como la dislexia, el tartamudeo y la afasia (Hämäläinen, Rupp, Soltész, Szücs & Goswami, 2012), la enfermedad de Parkinson (Oswal, Brown & Litvak, 2013), la enfermedad de Alzheimer (de Haan et al., 2012), la demencia (Franciotti et al., 2006), la depresión (Méndez et al., 2012) y la ceguera (Osaki et al., 2004). Estas investigaciones básicas sobre poblaciones clínicas guiarán el desarrollo de herramientas objetivas que mejoren el diagnóstico y los tratamientos.
Nuestra habilidad para entender el cerebro será mayor cuando haya la posibilidad de mapear la actividad en el nivel del sistema neural para varios estadios del desarrollo humano con una aproximación de neuroimagen multimodal, todo ello aunado con la exposición detallada de las explicaciones de las bases moleculares, genéticas y neuroquímicas. Cabe destacar que en Iberoamérica existe poca producción científica con la MEG como herramienta (Maestú, 2010; Santiuste-Bermejo & Santiuste-Díaz, 2008).