


Los receptores nucleares (RNs) constituyen una familia de factores transcripcionales activados por ligando que regulan la expresión de un gran número de genes de forma dependiente del tipo y contexto celular. La localización subcelular de los RNs es altamente dinámica y repercute sobre sus funciones como factores transcripcionales. En presencia de su ligando específico, los RNs se acumulan en el núcleo para modular la expresión de sus genes blanco. Por ende, la salida desde el núcleo a citoplasma de los RNs disminuye su acumulación nuclear y abate su actividad transcripcional. Por lo tanto, la exportación nuclear constituye un importante mecanismo de regulación de la actividad de los RNs. A pesar de su importancia, el proceso de exportación nuclear de los RNs no ha sido completamente explorado, sin embargo, los estudios que se tienen hasta ahora sugieren la participación de las proteínas CRM–1 y la Calreticulina (CRT) como mediadoras de este proceso. En esta revisión se destaca la exportación nuclear como un mecanismo regulador de las funciones de los RNs y se discuten las características estructurales y funcionales de las exportinas CRM–1 y CRT.
Nuclear receptors (RNs) are a family of transcription factors activated by ligand which regulate the expression of many genes dependent on the cellular type and context. The subcellular localization of RNs is highly dynamic and affects its function as transcriptional factors. In the presence of its specific ligand, the RNs increase in the nucleus to modulate the expression of their target genes. Thus, the exit from nucleus to cytoplasm of RNs decreases its nuclear accumulation and its transcriptional activity. Therefore, nuclear export is an important mechanism regulating the activity of RNs. Despite its importance, the process of nuclear export of RNs has not been fully studied. However, the studies made so far suggest the involvement of the CRM–1 and Calreticulin (CRT) proteins as mediators of this process. In this review, we highlight the nuclear export as a regulatory mechanism to control the functions of RNs and the structural and functional characteristics of CRM–1 and CRT exportins are discussed.
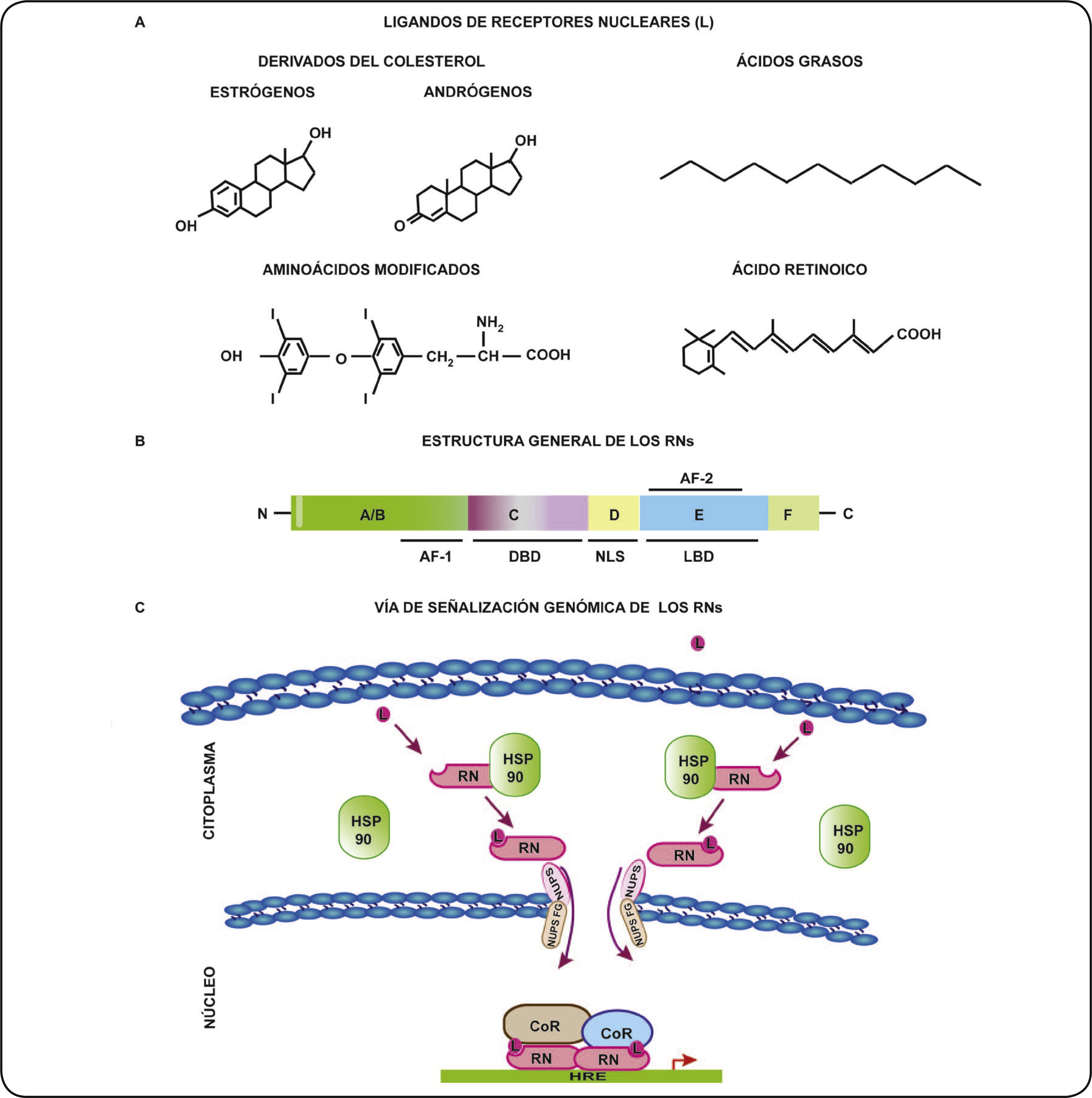
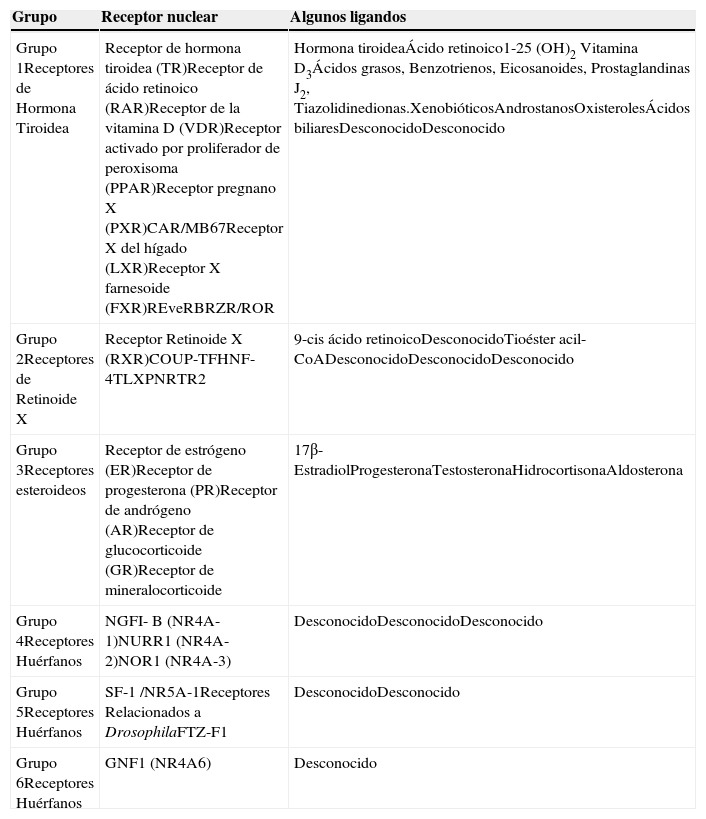
Los receptores nucleares (RNs) son factores de transcripción (FT) activados por ligando, que modulan la expresión de diferentes genes implicados en la diferenciación, apoptosis, crecimiento y el metabolismo, entre otros1,2. La expresión de los RNs es dependiente del tipo de tejido y contexto celular, y la desregulación de su expresión está involucrada en diversas enfermedades como cáncer, alteraciones cardiovasculares y diabetes3,4. Se han identificado 48 miembros que conforman una superfamilia de RNs que transducen señales de ligandos específicos de naturaleza lipofílica, como las hormonas esteroideas, tiroideas y ácidos grasos o ligandos no naturales, como los fármacos tiazolidinedionas1,5 (Figura 1A). De acuerdo a las características de sus ligandos, los RNs se subdividen en seis grupos (Tabla I), dentro de los cuales sobresalen por su relevancia en la homeostasis, el primer grupo de receptores para hormonas tiroideas y el tercer grupo integrado por receptores para las hormonas esteroideas. Existe también un grupo de RNs denominados huérfanos, debido a que sus ligandos aún no se identifican2,4,5.
 y su vía de señalización genómica. A. Ligandos representativos de los RNs. Los ligandos de los RNs son de diversa naturaleza, algunos ejemplos de ellos son los derivados del colesterol (estrógenos, progesterona, andrógenos, glucocorticoides), ácidos grasos (como las prostaglandinas), aminoácidos modificados (hormonas tiroideas) y el ácido retinoico. B. Dominios funcionales de los RNs. Los RNs poseen de cinco a seis dominios funcionales denominados con letras de la A-F. La región N-terminal o A/B contiene el dominio de activación transcripcional AF-1. La región C consiste en el dominio de unión al ADN (DBD) que es altamente conservado entre los RNs. La región D o “bisagra” conecta al DBD con la región E y además contiene la secuencia de localización nuclear (NLS). La región E conforma el dominio de unión al ligando o LBD y el segundo dominio de transactivación AF-2. El dominio F está presente sólo en algunos RNs. C. Vía se señalización genómica de los RNs. Los RNs son activados por su ligando específico en el citoplasma. La activación de los RNs conduce a su cambio conformacional, su disociación con proteínas chaperonas (HSP90), dimerización y traslocación al núcleo. En este compartimento, los RNs se unen a sus elementos de respuesta a la hormona (HRE) e interactúan con correguladores (CoR) (coactivadores o correpresores) para modular la expresión de sus genes blanco.")
Receptores nucleares (RNs) y su vía de señalización genómica. A. Ligandos representativos de los RNs. Los ligandos de los RNs son de diversa naturaleza, algunos ejemplos de ellos son los derivados del colesterol (estrógenos, progesterona, andrógenos, glucocorticoides), ácidos grasos (como las prostaglandinas), aminoácidos modificados (hormonas tiroideas) y el ácido retinoico. B. Dominios funcionales de los RNs. Los RNs poseen de cinco a seis dominios funcionales denominados con letras de la A-F. La región N-terminal o A/B contiene el dominio de activación transcripcional AF-1. La región C consiste en el dominio de unión al ADN (DBD) que es altamente conservado entre los RNs. La región D o “bisagra” conecta al DBD con la región E y además contiene la secuencia de localización nuclear (NLS). La región E conforma el dominio de unión al ligando o LBD y el segundo dominio de transactivación AF-2. El dominio F está presente sólo en algunos RNs. C. Vía se señalización genómica de los RNs. Los RNs son activados por su ligando específico en el citoplasma. La activación de los RNs conduce a su cambio conformacional, su disociación con proteínas chaperonas (HSP90), dimerización y traslocación al núcleo. En este compartimento, los RNs se unen a sus elementos de respuesta a la hormona (HRE) e interactúan con correguladores (CoR) (coactivadores o correpresores) para modular la expresión de sus genes blanco.
Clasificación de los RNs
| Grupo | Receptor nuclear | Algunos ligandos |
|---|---|---|
| Grupo 1Receptores de Hormona Tiroidea | Receptor de hormona tiroidea (TR)Receptor de ácido retinoico (RAR)Receptor de la vitamina D (VDR)Receptor activado por proliferador de peroxisoma (PPAR)Receptor pregnano X (PXR)CAR/MB67Receptor X del hígado (LXR)Receptor X farnesoide (FXR)REveRBRZR/ROR | Hormona tiroideaÁcido retinoico1-25 (OH)2 Vitamina D3Ácidos grasos, Benzotrienos, Eicosanoides, Prostaglandinas J2, Tiazolidinedionas.XenobióticosAndrostanosOxisterolesÁcidos biliaresDesconocidoDesconocido |
| Grupo 2Receptores de Retinoide X | Receptor Retinoide X (RXR)COUP-TFHNF-4TLXPNRTR2 | 9-cis ácido retinoicoDesconocidoTioéster acil- CoADesconocidoDesconocidoDesconocido |
| Grupo 3Receptores esteroideos | Receptor de estrógeno (ER)Receptor de progesterona (PR)Receptor de andrógeno (AR)Receptor de glucocorticoide (GR)Receptor de mineralocorticoide | 17β- EstradiolProgesteronaTestosteronaHidrocortisonaAldosterona |
| Grupo 4Receptores Huérfanos | NGFI- B (NR4A-1)NURR1 (NR4A-2)NOR1 (NR4A-3) | DesconocidoDesconocidoDesconocido |
| Grupo 5Receptores Huérfanos | SF-1 /NR5A-1Receptores Relacionados a DrosophilaFTZ-F1 | DesconocidoDesconocido |
| Grupo 6Receptores Huérfanos | GNF1 (NR4A6) | Desconocido |
La estructura de los RNs está conformada por dominios conservados denominados A, B, C, D, E y F, partiendo del extremo N–terminal al C–terminal. La región A/B comprende el primer dominio de activación transcripcional (AF–1), posteriormente se encuentra la región C que presenta el dominio de unión al ADN (DBD). La región D además de contener una secuencia de localización nuclear (NLS), actúa como una estructura “bisagra” entre la región C y E involucrada en los cambios conformacionales de los RNs. La región E contiene el dominio de unión al ligando (LBD) y un segundo dominio de función de activación transcripcional (AF–2)1. Algunos RNs presentan una región F relacionada con la modulación de su actividad (Figura 1B)6.
VÍA DE SEÑALIZACIÓN GENÓMICA DE LOS RNSLa vía de señalización genómica o clásica de los RNs, comienza cuando el ligando se une al dominio LBD de su RN específico. Como resultado de esta interacción ocurre un cambio conformacional en los RNs, que permite la disociación de proteínas que los mantienen en un estado inactivo en el citoplasma, como es el caso de la chaperona HSP90 y favorece su acumulación en el núcleo celular7. En el interior nuclear, los RNs como monómeros, homodímeros o heterodímeros se pueden unir a secuencias específicas en el ADN y reclutar proteínas correguladoras para dirigir la transcripción génica (Figura 1C)8. Los correguladores transcripcionales, no se unen directamente al ADN, sino que son reclutados por los RNs a través de sus dominios de transactivación (AF–1 y AF–2) y se dividen en coactivadores y correpresores3,8. Los coactivadores están asociados a la relajación de la estructura de la cromatina por poseer o reclutar proteínas con actividad de acetilación de histonas, mientras que los correpresores están asociados a la compactación de la cromatina por poseer actividad de desacetilación de histonas. De esta forma, los correguladores de los RNs modulan la compactación o descompactación de la cromatina para reprimir o inducir, respectivamente, la expresión génica9.
A pesar de que los RN son conocidos por sus funciones genómicas o clásicas como FT, algunos de ellos como los receptores para hormonas esteroideas, también han sido localizados en la membrana celular y pueden desencadenar cascadas de señalización intracelulares, activando diversos segundos mensajeros que conllevan a la activación de cinasas o vías dependientes de calcio para generar diversas respuestas biológicas. Estas vías rápidas de transducción de los RNs no relacionadas con sus funciones transcripcionales son conocidas como vías no genómicas o no clásicas10,11.
La actividad de los RNs es regulada por mecanismos como la degradación12, estabilidad13 y bloqueo de la dimerización14, así como por exportación nuclear15,16. Este último mecanismo podría ser crucial en la actividad de los RNs considerando que regulan la expresión génica1,17. A continuación se describen los aspectos que hasta ahora se han estudiado sobre la exportación nuclear de los RNs.
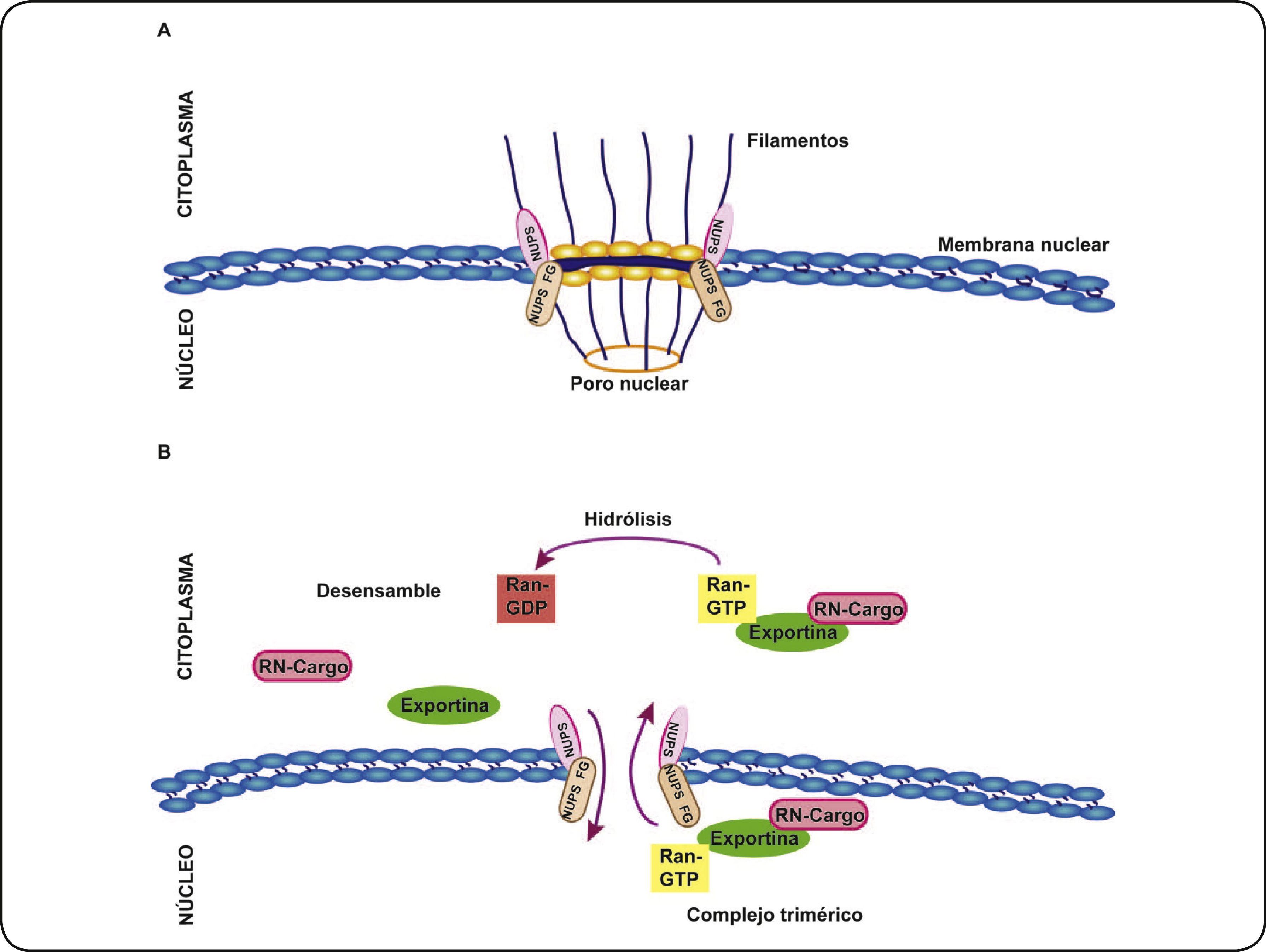
ENVOLTURA NUCLEAR Y PORO NUCLEARLa envoltura nuclear es una doble membrana que separa el compartimento citoplasmático del nuclear y permite el transporte bidireccional a través de los complejos proteicos del poro nuclear. A través de estos canales que miden de 100–150nm de diámetro18, se da el intercambio pasivo de iones, y de pequeñas moléculas y proteínas menores a 20kDa19. El complejo del poro nuclear está constituido por aproximadamente 100 proteínas llamadas nucleoporinas (NUPS), diez de ellas con residuos repetidos FG (Fenilalanina–Glicina) que participan en el transporte de proteínas como los RNs entre núcleo y citoplasma20 (Figura 2A).
 y filamentos proteicos nucleares y citoplasmáticos. B A través de los poros nucleares se lleva a cabo el proceso de exportación nuclear. Este proceso es dinámico y requiere la formación de un complejo trimérico entre la exportina, el RN cargo y RanGTP. El complejo trimérico interactúa con las NUPS F y G y atraviesa el poro nuclear, una vez que en el citoplasma ocurre la hidrólisis de RanGTP a RanGDP y en consecuencia la disociación del complejo, liberando al RN cargo y facilitando el regreso de la exportina al núcleo celular.")
Envoltura nuclear y la exportación nuclear. A La envoltura nuclear separa el citoplasma del núcleo celular y está constituida por poros nucleares conformados por nucleoporinas que contienen repeticiones ricas en Fenilalanina y Glicina (NUPS FG) y filamentos proteicos nucleares y citoplasmáticos. B A través de los poros nucleares se lleva a cabo el proceso de exportación nuclear. Este proceso es dinámico y requiere la formación de un complejo trimérico entre la exportina, el RN cargo y RanGTP. El complejo trimérico interactúa con las NUPS F y G y atraviesa el poro nuclear, una vez que en el citoplasma ocurre la hidrólisis de RanGTP a RanGDP y en consecuencia la disociación del complejo, liberando al RN cargo y facilitando el regreso de la exportina al núcleo celular.
Tanto la localización subcelular de los RNs como la de otros FT, es altamente dinámica entre los compartimentos del citoplasma y el núcleo. La entrada de los RNs al interior del núcleo es un proceso conocido como importación nuclear19. Para que se lleve a cabo este proceso se requiere de transportadores identificados como “importinas α y β” las cuales se unen con los RNs que se denominan proteínas “cargo”21. El transporte subcelular requiere de la interacción de las importinas con secuencias específicas de los RNs denominadas secuencia de localización nuclear (NLS, Nuclear Localization Signal)21. Diversos estudios demuestran que los RNs activados por su ligando, presentan cambios conformacionales y su translocación hacia el núcleo celular1,3,17. Es por ello que el estímulo con su ligando específico conduce a la acumulación nuclear de los RNs y favorece sus funciones como FT regulando la expresión génica.
Los RNs también requieren de transportadores denominados “exportinas” para salir del núcleo hacia el citoplasma. Este proceso se conoce como exportación nuclear y se lleva a cabo tras la interacción de las exportinas con las secuencias denominadas secuencias de exportación nuclear (NES, Nuclear Export Signal) de sus proteínas cargo. Las NES consideradas típicas o consenso se caracterizan por contener de 3 a 4 residuos hidrofóbicos ricos en leucinas, sin embargo existen modificaciones en esta secuencia denominada NES no consenso20. De forma interesante, en la mayoría de los RNs se han identificado NES no consenso, y algunos autores proponen que el DBD también podría funcionar como NES para que se lleve a cabo la exportación nuclear22, como se discutirá más adelante. Este proceso requiere la asociación del RN cargo, la exportina y Ran GTP para formar un complejo trimérico (RNcargo/exportina/RanGTP), que interactúa con repetidos FG de las NUPS del poro nuclear. Lo anterior, facilita la salida del complejo trimérico del núcleo y una vez en el citoplasma se efectúa la hidrólisis de RanGTP a RanGDP y consecuentemente el desensamble de dicho complejo (Figura 2B)20,23. Bajo estas consideraciones, el proceso de exportación nuclear resulta una importante estrategia para regular la función de los RNs. Sin embargo, son pocos los estudios y la información que se conoce sobre la exportación nuclear de los RNs y las exportinas implicadas en este proceso.
NES Y EXPORTINAS DE LOS RNsMientras que ya se ha identificado y caracterizado ampliamente la NLS de los RNs necesaria para su acumulación en el núcleo celular, no está del todo dilucidado la presencia de NES en los RNs22,24. De toda la superfamilia de los RNs, sólo en algunos de ellos se han identificado NES canónicos16, y en la mayoría de ellos NES no canónicos22,24, e incluso, algunos autores proponen la ausencia de NES en los RNs22. Además, para algunos RNs, como por ejemplo el receptor de estrógenos (ERα), existe discrepancia entre los NES no canónicos funcionales identificados por diversos grupos de investigación22,24,25. Otro aspecto interesante es que algunos autores sugieren que los dominios DBD altamente conservados entre los RNs podrían ser los responsables de su exportación nuclear22. También existen algunos grupos de investigación que consideran que otras proteínas podrían actuar como mediadoras junto a las exportinas para la salida de los RNs del núcleo26. Dentro de la complejidad que existe en la exportación nuclear de los RNs, se incluyen las exportinas. Aunque existen varias exportinas, para los RNs se han reportado principalmente dos: La exportina CRM–1 (Chromosome Region Maintenance 1)24,27 y la Calreticulina (CRT)28. Estas proteínas representan las dos principales vías de exportación nuclear documentadas para RNs. El conocer las características estructurales y funcionales de la CRM–1 y la CRT contribuye a entender la importancia de la exportación nuclear en la regulación de la actividad de los RNs.
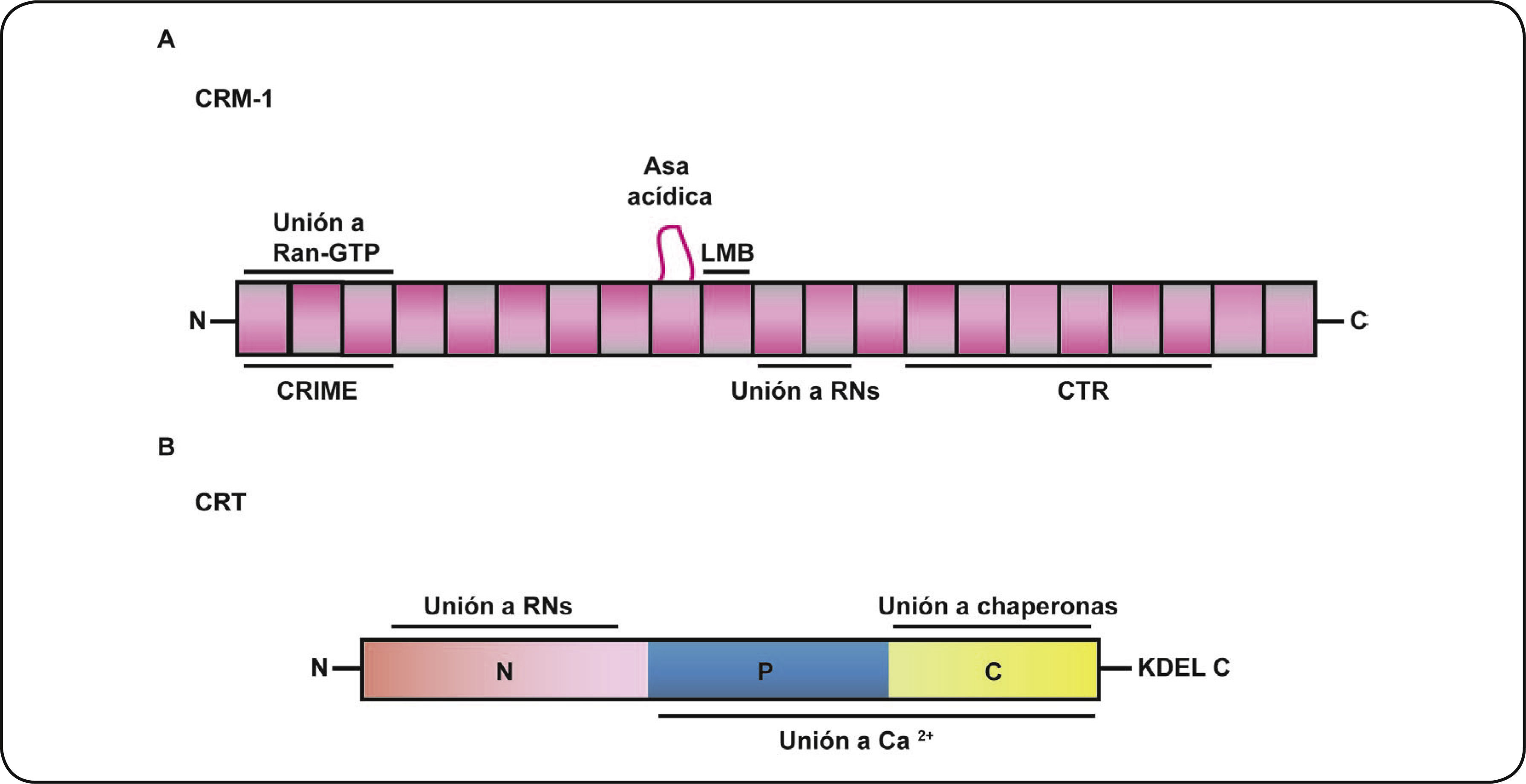
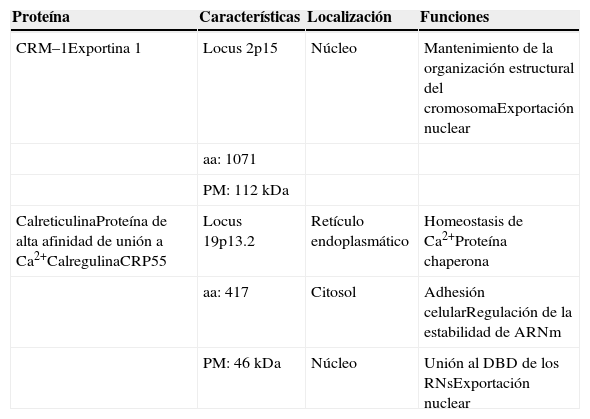
CRM–1 Y CRTLa proteína CRM–1 llamada también exportina 1, es un miembro de la superfamilia de las carioferinas. La CRM–1 es una proteína codificada por el gen localizado en el cromosoma humano 2p15, la cual está conservada entre diferentes especies como Xenopus laevis, Caenorhabditis elegans, Drosophila melanogaster, Rattus norvegicus y Homo sapiens20. La exportina CRM–1 tiene un peso molecular de 112 kDa (Tabla II) y estructuralmente tiene forma de anillo constituido de 21 repetidos HEAT (Huntingtin, elongation factor 3, protein phosphatase 2A and TOR1), formando dos α–hélices anti paralelas conectadas con asas de varios tamaños29. En el fragmento C–Terminal se localiza una región denominada CTR donde sugieren que se une con un asa de unión a Ran–GTP 30. El dominio CRIME (CRM–1, Importina β, Etc) de la región N–terminal junto con el asa acídica ubicada en la región intermedia interactúan con Ran–GTP. Entre los repetidos 11 y 12 se forma un surco hidrofóbico capaz de reconocer las NES de las proteínas cargo (Figura 3A).20
Principales características y funciones de la CRM1/CRT
| Proteína | Características | Localización | Funciones |
|---|---|---|---|
| CRM–1Exportina 1 | Locus 2p15 | Núcleo | Mantenimiento de la organización estructural del cromosomaExportación nuclear |
| aa: 1071 | |||
| PM: 112 kDa | |||
| CalreticulinaProteína de alta afinidad de unión a Ca2+CalregulinaCRP55 | Locus 19p13.2 | Retículo endoplasmático | Homeostasis de Ca2+Proteína chaperona |
| aa: 417 | Citosol | Adhesión celularRegulación de la estabilidad de ARNm | |
| PM: 46 kDa | Núcleo | Unión al DBD de los RNsExportación nuclear |
. El dominio N tiene importantes implicaciones en la unión a los RNs. Los dominios P y C contienen sitios de unión a Ca2+ con afinidad diferencial. En el dominio C además se localiza la señal peptídica KDEL de localización en retículo endoplasmático.")
Dominios estructurales de la CRM–1 y la CRT. La CRM1 es una proteína que está estructuralmente constituida por 21 repetidos HEAT. El dominio CRIME corresponde a la región N-terminal, mientras que la región CRT corresponde al dominio C-terminal. Contiene una asa acídica que interactúa con RanGTP y entre los repetidos HEAT 11 y 12 ocurre el reconocimiento con los RNs. B. Calreticulina o la CRT está estructuralmente dividida en tres dominios (N, P y C). El dominio N tiene importantes implicaciones en la unión a los RNs. Los dominios P y C contienen sitios de unión a Ca2+ con afinidad diferencial. En el dominio C además se localiza la señal peptídica KDEL de localización en retículo endoplasmático.
En lo que respecta a la CRT o Calreticulina (Calcium binding protein localized to the endoplasmic/sarcoplasmic Reticulum membranes), es una proteína de 46 kDa conformada por tres dominios estructurales y funcionales (Tabla II). La región N–terminal (dominio N) altamente conservada31,32, el dominio P rico en prolina que contiene una secuencia NLS putativa33 y la Región C–terminal (dominio C) de interacción con otras proteínas chaperonas que contienen la secuencia de retención en el retículo endoplasmático KDEL (Lys, Asp, Glut, Leu)31,32,34,35. Tanto el dominio P y C tienen afinidad de unión y la capacidad de unirse diferencialmente al calcio, mientras que el dominio N tiene importantes implicaciones en la unión con integrinas y con los RNs (Figura 3B).
LOCALIZACIÓN Y FUNCIÓN DE LA CRM–1 Y LA CRTDiversos estudios han mostrado que la CRM–1 es nuclear, localizándose principalmente en la membrana nuclear20,30,36. Ésto se identificó por primera vez en Schizosaccharomyces pombe, donde participa en mantener organizada la estructura del cromosoma36,37. Katrin Statde et al., en 1997, identificaron a la CRM–1 como una exportina en Saccharomyces cerevisiae y la renombraron como exportina 136. En ese mismo año se identificó que la CRM–1 de humano podía interactuar con componentes del poro nuclear como las NUPS38. La proteína CRM–1 puede llevar a cabo la exportación nuclear de diferentes proteínas, entre ellas los RNs pero también de ARN como es el caso de la subunidad pequeña y grande del ribosoma, sin embargo, la exportación nuclear de ARN por la CRM–1 requiere de proteínas adaptadoras39.
Mientras que la CRM–1 es una proteína nuclear, la CRT es una proteína que se localiza en diversos compartimentos celulares. Se identificó por primera vez en el retículo sarcoplasmático del músculo esquelético del conejo40, como una proteína de unión a Ca2+41. Las funciones de la CRT son diversas, modula las concentraciones intracelulares de Ca2+ y actúa como una chaperona al facilitar el plegamiento de las proteínas32. Aunque en un inicio se consideró una proteína exclusiva del retículo endoplasmático estudios posteriores documentaron su localización en el núcleo y el citoplasma de células no musculares28,42–48 (Tabla II). Al respecto, la CRT tiene una importante y novedosa función en el núcleo como exportina28. No obstante, esta nueva función de la CRT se ha reportado sólo para la proteína inhibidora de proteínas cinasas (PKI) y para algunos RNs27,28,49. El mecanismo de la CRT como exportina parece ser similar al de la CRM–1, debido a que se asocia para formar un complejo trimérico con una NES presente en PKI y RanGTP (PKI/CRT/RanGTP)28. Adicional a esta función, también se ha reportado como una exportina específica para el Receptor de glucocorticoides (GR) y como una exportina que podría tener un efecto sinérgico a la función de la exportina CRM–1 para los otros RNs27.
Los estudios que han establecido que la CRM–1 y la CRT actúan como exportinas de diferentes proteínas han requerido el uso de inhibidores que bloqueen su actividad28,50,51. En el caso de la CRT no se tienen inhibidores farmacológicos para controlar sus funciones y la estrategia con la que se cuenta es la reducción de su expresión mediante un sistema de ARN interferente51. En cuanto a la CRM–1, se ha usado un inhibidor que actúa a través de su unión covalente a residuos de cisteína de la CRM–1 para bloquear su unión a las NES de las proteínas cargo, conocido como Leptomicina B (LMB)50. Es relevante identificar nuevos inhibidores de estas exportinas, las cuales tienen importantes funciones en condiciones normales y también han sido asociadas con la deslocalización de diversas proteínas en algunas enfermedades (Tabla III).
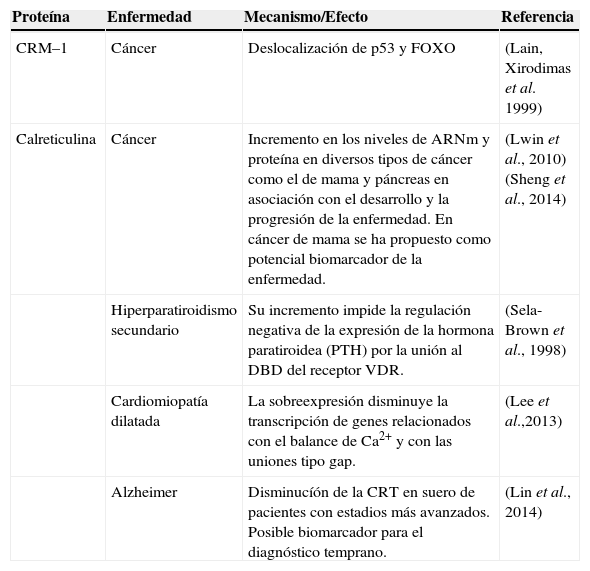
La CRM–1 y la Calreticulina en procesos fisiopatológicos
| Proteína | Enfermedad | Mecanismo/Efecto | Referencia |
|---|---|---|---|
| CRM–1 | Cáncer | Deslocalización de p53 y FOXO | (Lain, Xirodimas et al. 1999) |
| Calreticulina | Cáncer | Incremento en los niveles de ARNm y proteína en diversos tipos de cáncer como el de mama y páncreas en asociación con el desarrollo y la progresión de la enfermedad. En cáncer de mama se ha propuesto como potencial biomarcador de la enfermedad. | (Lwin et al., 2010)(Sheng et al., 2014) |
| Hiperparatiroidismo secundario | Su incremento impide la regulación negativa de la expresión de la hormona paratiroidea (PTH) por la unión al DBD del receptor VDR. | (Sela-Brown et al., 1998) | |
| Cardiomiopatía dilatada | La sobreexpresión disminuye la transcripción de genes relacionados con el balance de Ca2+ y con las uniones tipo gap. | (Lee et al.,2013) | |
| Alzheimer | Disminucíón de la CRT en suero de pacientes con estadios más avanzados. Posible biomarcador para el diagnóstico temprano. | (Lin et al., 2014) |
La desregulación de la CRM–1 se ha reportado en diferentes tipos de cáncer, por ejemplo, altos niveles de la CRM–1 conducen a la deslocalización de supresores tumorales p53 y FOXO para afectar sus funciones en células de distintos tipos de cáncer52–54. Por esta razón, algunos estudios se están enfocando en el diseño de inhibidores específicos y no tóxicos de la actividad de la CRM–153. La CRT, en cambio, es una proteína multifuncional, importante para la homeostasis. La expresión de la CRT es crucial para el desarrollo embrionario y en el adulto regula las concentraciones de calcio, actúa como chaperona y como exportina de la proteína PKI y los RNs y posiblemente de otras proteínas aún no descubiertas28. En algunas enfermedades se ha visto una desregulación de la expresión de la CRT, una de ellas en cáncer de distintos tejidos (Tabla III)55,56. La inhibición de la actividad de la CRM–1 y la CRT resulta por lo tanto importante para el estudio de sus proteínas cargo y de los factores y señales implicados en la exportación nuclear. Además, es interesante considerar que los inhibidores de la actividad de exportinas de la CRM–1 y la CRT en contextos determinados pueden tener implicaciones dentro de las estrategias terapéuticas asociadas con la deslocalización de FT en distintos padecimientos.
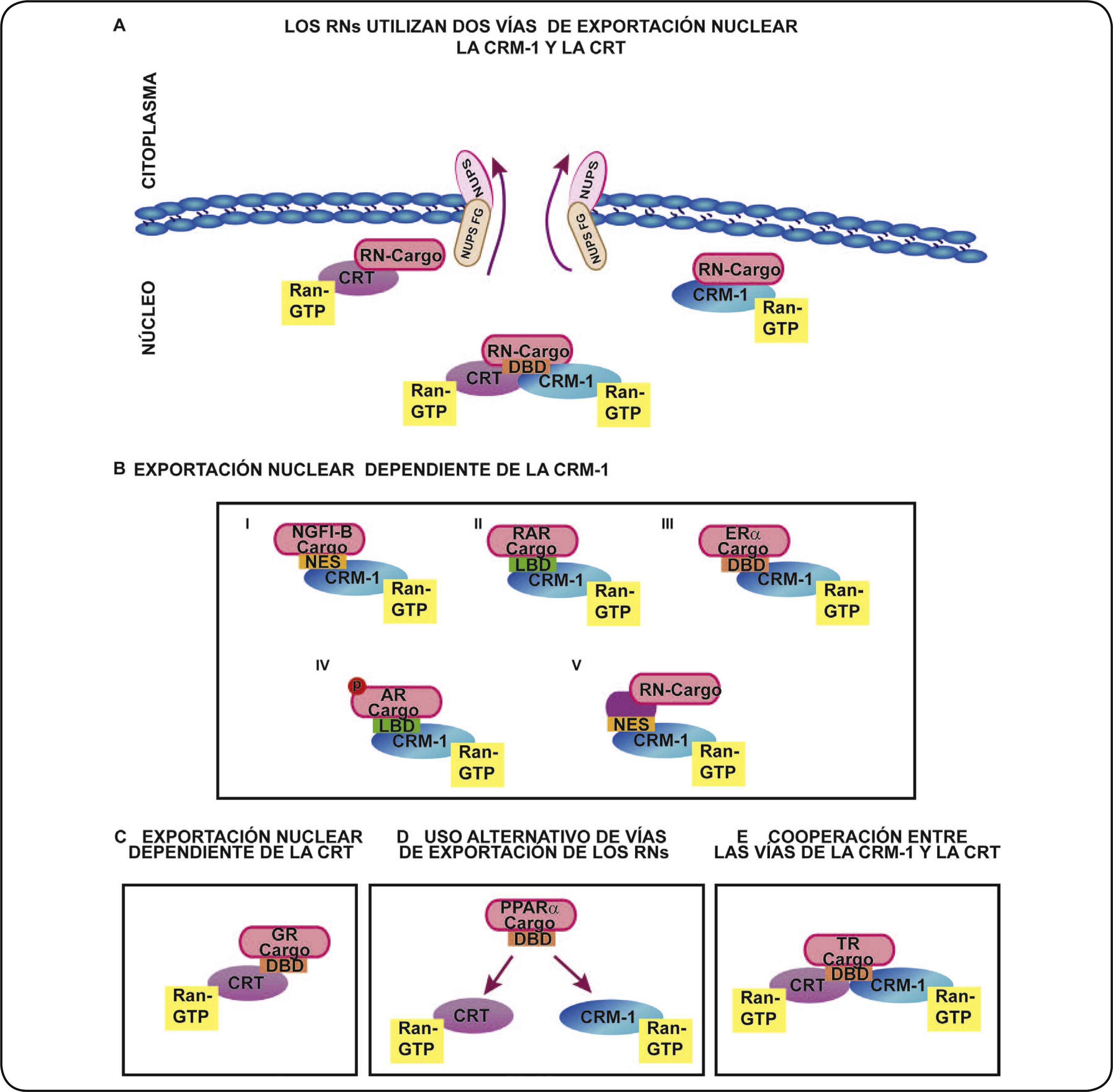
CRM–1 Y CRT COMO LAS PRINCIPALES EXPORTINAS DE LOS RNsComo se ha mencionado, la CRM–1 es la exportina más estudiada que está implicada en la salida de diversas proteínas del núcleo. La exportación nuclear de la mayoría de los RNs evaluados es también dependiente de la actividad de la CRM–1 (Figura 4A y B). A pesar de las discrepancias en la identificación de las NES de los RNs, el uso del inhibidor LMB, establece la participación de la CRM–1 como la exportina requerida para el transporte del núcleo al citoplasma de un gran número de RNs, entre ellos el ERα24, el Receptor de andrógenos (AR)57 y el NR4A126, entre otros.
 o los NES no canónicos localizados en el LBD (II) o DBD (III) de los RNs. También se ha propuesto que modificaciones postraduccionales de los RNs podrían afectar su exportación nuclear (IV). Además, proteínas adaptadoras pueden ser requeridas para la interacción de la CRM–1 con los RNs (V). C. La exportación nuclear dependiente de la CRT. El DBD de los RNs se asocia a la CRT para su salida del núcleo. D. Uso alternativo de vías de exportación de los RNs. Bajo ciertas condiciones los RNs pueden ser exportados por una u otra exportina. E. Cooperación entre las vías de la CRM–1 y la CRT. Para incrementar la eficiencia de la salida de los RNs del núcleo, ambas vías pueden acoplarse de forma cooperativa. Receptor de hormona tiroidea (TR); Receptor de estrógenos (ERα); receptor de glucocorticoides (GR); receptor de andrógenos (AR). Gen B inducible por el factor de crecimiento neuronal (NGFI-B) Receptor de ácido retinoico (RAR), Receptor Activado por Proliferador de Peroxisoma (PPAR).")
Vías de exportación nuclear de los RNs. A. Los RNs utilizan dos vías de exportación nuclear, la CRM–1 y la CRT. Algunos RNs son dependientes de la CRM1, otros lo son de la CRT y algunos otros RNs pueden usar alternativamente una u otra exportina, o bien usar de forma cooperativa ambas exportinas para salir del núcleo celular. B. La exportación nuclear dependiente de la CRM–1. Esta vía de exportación nuclear puede efectuarse a través de la asociación de la CRM–1 con los NES canónicos (I) o los NES no canónicos localizados en el LBD (II) o DBD (III) de los RNs. También se ha propuesto que modificaciones postraduccionales de los RNs podrían afectar su exportación nuclear (IV). Además, proteínas adaptadoras pueden ser requeridas para la interacción de la CRM–1 con los RNs (V). C. La exportación nuclear dependiente de la CRT. El DBD de los RNs se asocia a la CRT para su salida del núcleo. D. Uso alternativo de vías de exportación de los RNs. Bajo ciertas condiciones los RNs pueden ser exportados por una u otra exportina. E. Cooperación entre las vías de la CRM–1 y la CRT. Para incrementar la eficiencia de la salida de los RNs del núcleo, ambas vías pueden acoplarse de forma cooperativa.
Receptor de hormona tiroidea (TR); Receptor de estrógenos (ERα); receptor de glucocorticoides (GR); receptor de andrógenos (AR). Gen B inducible por el factor de crecimiento neuronal (NGFI-B) Receptor de ácido retinoico (RAR), Receptor Activado por Proliferador de Peroxisoma (PPAR).
En otros RNs las NES se han identificado y probado experimentalmente comprobando su funcionalidad (Figura 4B– I)16, mientras que otros sólo han sido identificados mediante estudios bioinformáticos siendo consideradas como las NES putativas58. En algunos otros casos existe divergencia sobre la ubicación de las NES funcionales entre el LBD o DBD de los RNs por diversos grupos de investigación (Figura 4B II, III). En el caso del ERα, un estudio identificó una NES en la región del DBD22, mientras que otros dos estudios diferentes reportaron una NES localizada en el LDB24,25. No obstante, todos estos estudios establecen que para la exportación nuclear de los RNs se requiere de la exportina CRM–1.
El proceso de exportación nuclear dependiente de la CRM–1 de algunos RNs puede estar acoplado a modificaciones postraduccionales de los mismos (Figura 4B IV), un ejemplo importante es el AR. En un estudio se determinó que la inhibición de la cinasa de serina treonina GSK–3, bloquea la fosforilación del AR para facilitar su exportación nuclear57. Otros RNs, parecen requerir una proteína para la formación de un complejo con CRM–1 y favorecer el proceso de su exportación nuclear. Ejemplo de ello es el RN huérfano NR4A1, el cual requiere a una pequeña proteína denominada ISG12 para favorecer la salida de NR4A1 del núcleo al citoplasma26. También se ha propuesto que los RNs que carecen de NES podrían usar proteínas adaptadoras con secuencias NES para su exportación nuclear por CRM–1 (Figura 4B V)26.
La CRT es la otra exportina utilizada por los RNs para llevar a cabo su salida del núcleo (Figura 4A). La función de la CRT como una exportina de los RNs resulta relevante debido a que únicamente se ha mostrado su participación en la exportación nuclear de la proteína PKI y de algunos RNs. De forma interesante, la proteína PKI contiene una NES a través de la cual se asocia con la CRT para ser transportada al exterior del núcleo, en contraste, no todos los RNs a los que se asocia la CRT, presentan NES (Figura 4C)22. Burns et al. en 1994 reportaron que interactúa, mediante su dominio N, con una secuencia de aminoácidos KXFFKR situada en el DBD de los RNs, como GR, AR y el receptor de la vitamina D (VDR)59. La interacción de la CRT con estos RNs bloquea su reclutamiento a sus elementos de respuesta situados en el ADN de sus genes blanco. Además, en el mismo estudio, la sobreexpresión de la CRT conduce a la disminución de la actividad transcripcional de estos RNs.
Es interesante como otro estudio mostró que la CRT es capaz de regular la exportación nuclear de los RNs mediante la formación de un complejo con la proteína Ran GTP, por un mecanismo similar a la exportina CRM–128. De esta forma, se evidenció que la CRT participa en la salida del núcleo al citoplasma del GR. Por lo tanto, la CRT parece requerir interactuar con secuencias como las NES localizadas dentro del DBD del GR para inducir su salida del núcleo celular28. Dado que el DBD es conservado en la superfamilia de los RNs, muchos otros miembros de esta familia podrían cambiar su localización subcelular de manera dependiente de la CRT. Estos datos sugieren que el DBD podría representar una atípica NES de los RNs reconocida por la exportina CRT28.
Por otra parte, se mostró que el heterodímero VDR/receptor x retinoide (RXR) se une al promotor de la hormona paratiroidea (PTH) para regular negativamente su expresión. En la enfermedad de hiperparatiroidismo, los altos niveles de la proteína CRT bloquean la actividad de VDR60. La CRT se une al motivo KXFF(K/R)R localizado en el DBD del VDR para bloquear su asociación al ADN y así controlar su actividad, teniendo como consecuencia la desrepresión de la PTH60. Este estudio sugiere que la CRT podría unirse al DBD de otros RNs para bloquear su unión a sus elementos de respuesta a la hormona (HRE) y posteriormente promover su salida del núcleo celular60.
La CRT parece ser muy importante en la exportación nuclear de algunos RNs insensibles al efecto del inhibidor LMB, pero además también se han identificado los RNs cuya salida del núcleo es promovida por la colaboración de ambas exportinas, CRT y CRM–115,16,57,61. Dos ejemplos son: el Receptor Activado por Proliferador de Peroxisoma (PPARα) y PPARγ que contienen una NES en el DBD reconocida por la proteína CRT para su exportación nuclear62. En contraste, mutantes de las NES de PPARα y PPARγ bloquean su exportación nuclear por la CRT62. Sin embargo, estos receptores también contienen una NES localizada en la región LBD que es sensible a la LMB. Por lo tanto, la exportación nuclear de PPARα y PPARγ es dependiente de la CRT y la CRM–1, por su interacción con diferentes NES, localizadas en el DBD y LBD, respectivamente. En este estudio se propone que la CRT es la principal exportina de ambos PPARs y que la CRM–1 podría ser una exportina auxiliar que se utiliza bajo condiciones específicas (Figura 4D)62.
Ambas exportinas de los NRs también podrían actuar no sólo como una vía alternativa una de la otra, sino en conjunto para lograr el objetivo en común de la exportación nuclear de ciertos RNs, como ocurre con el Receptor tiroideo (TR)27. Este RN modifica su localización nuclear al citoplasma a través de la acción cooperativa entre la CRT y la CRM–1. Se propone que la cooperación entre ambas vías de exportación nuclear permite una eficiente translocación núcleo–citoplasma de algunos RNs (Figura 4E)27. Cabe señalar que recientemente en un solo reporte se mencionan que otras exportinas participan en la exportación nuclear de TR, sin embargo, más estudios aún son necesarios para dilucidar si éstas podrían también asociarse con otros RNs.
LA IMPORTANCIA DE LA EXPORTACIÓN DE LOS RNsLos RNs en respuesta a su ligando se acumulan en el núcleo para llevar a cabo sus funciones transcripcionales. Cómo y cuándo son transportados al exterior del núcleo luego de la estimulación con su ligando no es conocido. No obstante, la exportación nuclear de estas proteínas resulta un mecanismo crucial en el control de la intensidad y duración de su vía de señalización genómica en contextos celulares específicos.
Mientras que se describe un mecanismo general para la importación nuclear de prácticamente todos los RNs, su exportación nuclear se lleva a cabo principalmente por dos diferentes vías, la CRM–1 y la CRT (Figura 4A). A pesar de que las vías de exportación nuclear para los RNs realizan la misma función, las proteínas CRM–1 y CRT no están relacionadas a nivel de secuencia, ni estructura (Figura 3). El contar con dos vías de exportación para los RNs podría sugerir la importancia de este mecanismo en la regulación de la transcripción de sus genes. También, las vías de la CRM–1 y la CRT indican la trascendencia de promover la salida de RNs del núcleo y sugieren una posible regulación o coordinación entre ambas exportinas para lograr su función en condiciones específicas (Figura 4D, E)27. Así, la exportación nuclear de los RNs podría representar un mecanismo evolutivamente conservado para regular su actividad como FT.
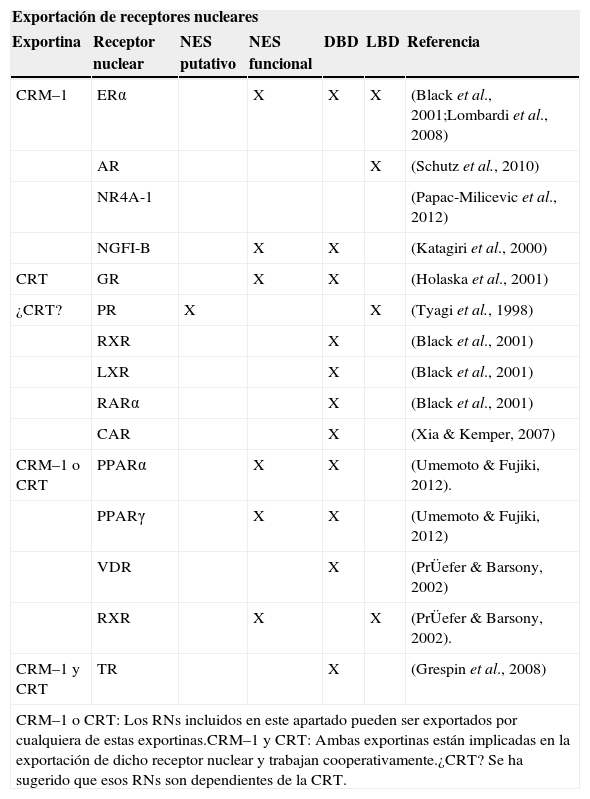
El concepto del DBD como un dominio necesario para la vía de la CRM–1 y la CRT principalmente, indica que el proceso de exportación nuclear podría ser un mecanismo general para controlar la función de todos los RNs22,28. En los estudios que se han realizado, más de 10 RNs requieren el DBD para ser transportados del núcleo al citoplasma (Tabla IV). El DBD es altamente conservado y estructuralmente similar entre los RNs y aunque se asocia con la exportina CRT, también se reportan algunos tipos de NES en el DBD asociados con la CRM–1. De esta manera, el DBD es suficiente para el cambio de localización del núcleo al citoplasma de los RNs y podría definir un nuevo tipo de señal de exportación nuclear. El requerimiento del DBD para la exportación nuclear de los RNs presume que todos éstos podrían competir por las exportinas para ser llevados al exterior del núcleo y que por ende deben existir otras señales que modulen dicha competencia y confieran especificidad.
Vías de exportación nuclear/NES para algunos RNs reportados
| Exportación de receptores nucleares | ||||||
|---|---|---|---|---|---|---|
| Exportina | Receptor nuclear | NES putativo | NES funcional | DBD | LBD | Referencia |
| CRM–1 | ERα | X | X | X | (Black et al., 2001;Lombardi et al., 2008) | |
| AR | X | (Schutz et al., 2010) | ||||
| NR4A-1 | (Papac-Milicevic et al., 2012) | |||||
| NGFI-B | X | X | (Katagiri et al., 2000) | |||
| CRT | GR | X | X | (Holaska et al., 2001) | ||
| ¿CRT? | PR | X | X | (Tyagi et al., 1998) | ||
| RXR | X | (Black et al., 2001) | ||||
| LXR | X | (Black et al., 2001) | ||||
| RARα | X | (Black et al., 2001) | ||||
| CAR | X | (Xia & Kemper, 2007) | ||||
| CRM–1 o CRT | PPARα | X | X | (Umemoto & Fujiki, 2012). | ||
| PPARγ | X | X | (Umemoto & Fujiki, 2012) | |||
| VDR | X | (PrÜefer & Barsony, 2002) | ||||
| RXR | X | X | (PrÜefer & Barsony, 2002). | |||
| CRM–1 y CRT | TR | X | (Grespin et al., 2008) | |||
| CRM–1 o CRT: Los RNs incluidos en este apartado pueden ser exportados por cualquiera de estas exportinas.CRM–1 y CRT: Ambas exportinas están implicadas en la exportación de dicho receptor nuclear y trabajan cooperativamente.¿CRT? Se ha sugerido que esos RNs son dependientes de la CRT. | ||||||
En forma global, se puede entender la exportación nuclear como un mecanismo absoluto y trascendental de apagado del proceso de transcripción génica, considerando que los RNs son FT y que el núcleo celular es su sitio de acción. Sin embargo, las consecuencias de la exportación nuclear no sólo conllevan a la inhibición de la actividad transcripcional de los RNs por relocalizarlos en el citoplasma, sino además podría tener efectos secundarios dentro de la vía de señalización de los RNs. Por ejemplo, al ser llevados los RNs fuera del núcleo se podrían desfavorecer sus funciones genómicas, pero favorecer sus funciones no genómicas, las cuales han sido descritas para algunos RNs en ciertos contextos celulares. También, tras la salida del núcleo de los RNs es probable que muchos de ellos puedan ser degradados para mantener un balance en sus niveles y en sus funciones, como ocurre con el GR49 y el AR63 que tras la inducción de su exportación nuclear hay un incremento en su ubiquitinación y degradación por el sistema de ubiquitina proteosoma49,63.
En diversas enfermedades se sabe que la actividad transcripcional de algunos RNs está desregulada por diferentes mecanismos moleculares, uno de ellos, podría ser la salida desde el núcleo al citoplasma de los RNs53,60. Muchas enfermedades asociadas con los RNs como el cáncer de diversos tejidos implican un incremento de su actividad transcripcional en el núcleo de las células3. Es probable que en estos padecimientos el transporte de los RNs por los compartimentos celulares esté afectado, influyendo en su función. La identificación de las bases moleculares de la dinámica de la exportación nuclear de los RNs en condiciones normales y en alteraciones particulares de tejidos específicos, ayudará a entender mejor la importancia de este proceso y dicho conocimiento pueda ser usado para el descubrimiento de nuevas estrategias terapéuticas. Es por tanto, un campo de investigación abierto que vislumbra importantes hallazgos dentro del estudio de los RNs.
CONCLUSIONES Y PERSPECTIVASDentro del compartimento nuclear de las células, los RNs modulan la expresión de sus genes blanco. La salida de los RNs del núcleo constituye un mecanismo control de sus vías de señalización genómica. El transporte de los RNs del núcleo al citoplasma es un proceso dependiente de las exportinas CRM–1 y CRT. Algunos RNs parecen ser preferenciales para una u otra exportina, o bien para ambas. En algunos casos, la exportación nuclear de los RNs puede verse afectada por modificaciones postraduccionales de los RNs, así como de la asociación con otras proteínas adaptadoras de las exportinas. Tanto la CRM–1 como la CRT pueden reconocer secuencias NES canónicas y/o no canónicas que pueden encontrarse principalmente en el LBD y DBD. El DBD conservado entre los RNs se postula importante en el proceso de exportación nuclear. Aún falta detallar los mecanismos implicados en la exportación nuclear de los RNs e identificar las señales que regulan este proceso, sin embargo, a pesar de su complejidad, es clara su transcendencia en la regulación negativa de la actividad transcripcional de los RNs en los tejidos y contextos específicos donde se expresan estos RNs. En adelante, es importante determinar si las exportinas CRM–1 y CRT son las únicas exportinas implicadas en la regulación de los RNs, además de entender por qué algunos RNs son dependientes solamente de la CRM–1, si existen más RNs únicamente dependientes de la CRT, y bajo qué circunstancias existe una cooperación entre ambas exportinas o cuándo y por qué actúan como exportinas alternativas una de la otra o cooperativamente. Otro aspecto a dilucidar es la implicación de las NES y/o el DBD de los RNs en la exportación nuclear. Además, aún se requiere conocer si existen para todos los RNs proteínas adaptadoras que puedan facilitar su transporte del núcleo al citoplasma. El clarificar los mecanismos moleculares de la exportación nuclear de cómo un proceso de regulación de la localización y actividad de los RNs podría conducir a la identificación de nuevos blancos terapéuticos en las diversas enfermedades asociadas a la desregulación de los RNs.
Al Programa de Investigación de Cáncer de Mama del Instituto de Investigaciones Biomédicas de la Universidad Nacional Autónoma de México. Agradecemos al Dr. Josué Orlando Ramírez Jarquín por su apoyo técnico para la revisión del manuscrito. A Edgar Ortiz Hernández por sus contribuciones en esta revisión. Bibiana Ortega Domínguez es becaria CONACyT del Programa de Doctorado en Ciencias Biomédicas, UNAM.