




La neurobiología de la agresión y la violencia es de interés para la psicología jurídica porque buena parte de la conducta delictiva tiene componentes violentos. En esta revisión se definen en primer lugar ambos conceptos, para diferenciar a continuación los tipos de agresión (impulsiva vs. instrumental) que aparecen en la literatura científica y finalmente analizar las estructuras nerviosas que según los estudios sobre lesiones cerebrales o de neuroimagen están asociadas con la agresión. Esta revisión destaca: a) las estructuras subcorticales como el hipotálamo/tronco del encéfalo, donde se genera la conducta agresiva y la amígdala, implicada en procesar estímulos emocionalmente destacados; b) las estructuras corticales como la corteza prefrontal (que comprende la corteza orbitofrontal, la corteza prefrontal ventromedial y la corteza cingulada anterior), que parecen ser hipofuncionales en los sujetos violentos. Por último, se revisan estudios sobre el papel del neurotransmisor serotonina en la manifestación del comportamiento agresivo.
Legal psychology has become interested in the neurobiology of agression and violence because in many cases criminal behaviour has violent components. The objective of this review is triple: to give a definition of both concepts; to show the different kinds of aggression (impulsive vs. instrumental) described in the scientific literature; and to analyse the neural structures that have been associated with aggression through lesion or neuroimaging studies. This review highlights mainly: a) subcortical structures such as hypothalamus and brain stem, where aggression is generated, and the amygdala, involved in processing emotionally salient stimuli; b) cortical structures such as the prefrontal cortex (which includes the orbitofrontal cortex, ventromedial prefrontal cortex, and anterior cingulate cortex) which seem to be hypofunctional in the aggressive subjects. Finally, studies about the role of the neurotransmitter serotonin and its role in aggressive behavior are reviewed.
Una definición adaptativa de agresividad sería la expuesta por Valzelli (1983), que la considera como un componente de la conducta normal que se expresa para satisfacer necesidades vitales y para eliminar o superar cualquier amenaza contra la integridad física y/o psicológica. Estaría orientada a la conservación del individuo y de la especie y solamente en el caso de la actividad depredadora conduciría a la destrucción del oponente, llegando hasta provocar su muerte. Siguiendo esta línea, se ha propuesto una distinción entre agresión y violencia basada en criterios de utilidad biológica. La primera sería una conducta normal, fisiológica que ayuda a la supervivencia del individuo y su especie (Archer, 2009). El término violencia se aplicaría a formas de agresión en las que el valor adaptativo se ha perdido, que pueden reflejar una disfunción de los mecanismos neurales relacionados con la expresión y control de la conducta agresiva, en tanto que su objetivo es el daño extremo, incluso llegando a la muerte de la víctima (Anderson y Bushman, 2002; Daly y Wilson, 2003; Mas, 1994). En consecuencia, la violencia está influida por factores culturales, ambientales y sociales que modelan la manera concreta de expresar la conducta violenta (Alcázar, 2011; Siegel y Victoroff, 2009). No obstante, esta conceptualización no implica necesariamente que la agresión y la violencia sean dos categorías separadas; al contrario, desde esta perspectiva se puede considerar que tanto la agresión como la violencia son conductas complejas que en dosis moderadas pueden tener una función adaptativa en entornos ambientales exigentes que supongan retos para la supervivencia del individuo. De este modo, la agresión y la violencia podrían considerarse como parte de una misma dimensión continua (Vassos, Collier y Fazel, 2014).
De esta manera, la violencia debería ser considerada como el resultado final de una cadena de eventos vitales durante la cual los riesgos se van acumulando y potencialmente se refuerzan unos a otros, hasta que la conducta violenta se dispara en una situación específica (Gronde, Kempes, van El, Rinne y Pieters, 2014).
Así, los factores psicosociales y biológicos interactúan modelando la conducta violenta. Por consiguiente, las causas psicosociales y biológicas del crimen violento están inseparablemente unidas y en constante interacción (Gronde, Kempes, van El, Rinne y Pieters, 2014; Stahl, 2014).
Tipos de agresiónLa agresión es un constructo complejo y heterogéneo, por lo que interesa identificar subtipos o clases de agresión para su estudio (Stahl, 2014). Es clásica la distinción entre agresión premeditada (predatoria, instrumental) e impulsiva (afectiva, reactiva). Por tanto, se puede considerar que ha habido consenso en la codificación dicotómica de la agresión en dos categorías: impulsiva vs. instrumental (Alcázar, 2011; Cornell et al., 1996; Kockler, Nelson, Meloy y Stanford, 2006; Raine et al., 1998; Stanford et al., 2003; Weinshenker y Siegel, 2002; Woodworth y Porter, 2002). La primera es una reacción abrupta, en “caliente”, como una respuesta a una percepción de provocación o amenaza, mientras que la instrumental es una respuesta premeditada, orientada a un objetivo y a “sangre fría”. Ahora bien, dando por sentada esta clasificación dicotómica, se debe subrayar que es muy frecuente que los actos violentos puedan mostrar características de ambas, impulsiva e instrumental (Bushman y Anderson, 2001; Penado, Andreu y Peña, 2014). Por ejemplo, la conducta agresiva se puede dar de manera repentina como respuesta a una provocación percibida, con enfado y afecto hostil. Pero es que, además, esta misma conducta agresiva podría darse de una manera controlada y con un objetivo bien delimitado (intimidación, elevación de la autoestima, etc.) (Rosell y Siever, 2015).
Una clasificación similar a la impulsiva/instrumental es la que distingue entre agresión reactiva y proactiva. En esta clasificación se asume desde el principio, a diferencia de la anterior que opta por una concepción categórica de base, que ambas coexisten y contribuyen conjuntamente al nivel de agresión total del individuo y cada una es evaluada de manera dimensional (Rosell y Siever, 2015).
El tipo reactivo es el que más se parece a la categoría impulsiva y sería una agresión que sucede como reacción a una frustración o a una provocación percibida (normalmente en un contexto interpersonal). Este tipo de agresión está invariablemente acompañada de hostilidad, ira o rabia. Su objetivo básico sería compensar o mitigar el estado afectivo desagradable que siente el sujeto. Por otra parte, la agresión proactiva está caracterizada por que no tiene que ir necesariamente acompañada de un estado emocional desagradable (ira, rabia, etc.), suele ser iniciada por el agresor más que como reacción a una provocación y está motivada de manera explícita por la expectativa del agresor de obtener alguna recompensa (un objeto, un premio, poder, estatus, dominancia social, etc.) (Penado et al., 2014; Rosell y Siever, 2015). Estas dos maneras de agresión coexisten y están altamente correlacionadas. Sin embargo, la reactiva se ha vinculado con historia de abuso (Kolla et al., 2013), impulsividad (Cima, Raine, Meesters y Popma, 2013; Raine et al., 2006), emociones negativas (como ira y frustración) y dureza emocional (que es un componente de la psicopatía) (Cima et al., 2013). Por su parte, la proactiva se ha relacionado positivamente con la psicopatía (Kolla et al., 2013), agresión física y delitos violentos (Cima et al., 2013; Rosell y Siever, 2015).
Neurobiología de la agresión impulsiva (centros y circuitos)La agresión impulsiva es generalmente una respuesta inmediata a un estímulo del medio ambiente. Para Stahl (2014) este tipo de violencia puede reflejar “una hipersensibilidad emocional y una percepción exagerada de las amenazas, lo que puede ir ligado a un desequilibrio entre los controles inhibidores corticales de arriba-abajo y los impulsos límbicos de abajo-arriba” (p. 360). El paradigma clásico, que liga la corteza prefrontal y áreas límbicas como la amígdala, es que la actividad en estructuras límbicas subcorticales como la amígdala es modulada por una influencia inhibidora desde estructuras corticales como la corteza prefrontal orbitofrontal (COF). De tal manera que un individuo que no restrinja su agresión impulsiva tendrá una gran actividad en la zona amigdalar y poca actividad inhibidora en la zona COF, un individuo que sea capaz de controlar su agresión impulsiva tendrá una gran actividad en la COF y un individuo con una lesión en la COF tendrá un aumento de agresión impulsiva.
¿Pero dónde se origina el comportamiento de agresión en el sistema nervioso central?
Hipotálamo y sustancia gris periacueductalDiversos estudios de lesión y estimulación realizados en la primera mitad del siglo XX en gatos mostraron que existía una región en el hipotálamo posterior que al ser destruida y separada de sus conexiones con centros situados en el tronco del encéfalo o la médula espinal hacia desaparecer un comportamiento agresivo de rabia (sham rage o falsa rabia) que no parecía estar asociado con la ira real y que no siempre iba dirigido hacia el estímulo que la había desencadenado, y al ser estimulada dicha región provocaba la aparición de dicho comportamiento (Finger, 1994; Siegel, 2005).
Los estudios sobre las bases neurobiológicas de la agresión en el gato han conducido a la descripción de un “ataque afectivo”, caracterizado por respuestas emocionales típicas del sistema nervioso simpático, y un “ataque predador”, sin aquellas. (McEllistrem, 2004; Siegel, 2005). Este tipo de conductas se pueden considerar análogas a la agresión afectiva y la agresión instrumental en humanos.
El “ataque afectivo” puede ser controlado desde una gran extensión del hipotálamo medial, extendiéndose hacia el tronco del encéfalo donde se encuentran centros nerviosos que controlan la expresión del ataque como es bufar y gruñir (McEllistrem, 2004). Además del hipotálamo medial también están implicados la amígdala medial, de la que recibe información excitadora, y la sustancia gris periacueductal dorsal del tronco del encéfalo, a la que envía información excitadora. Desde esta última hay conexiones excitadoras con el locus coeruleus y el núcleo solitario que median las respuestas autónomas durante el “ataque afectivo”; también hay conexiones excitadoras con los centros de los nervios trigémino y facial para el control de la apertura de la boca y las vocalizaciones; por último hay conexiones excitadoras indirectas con la médula espinal cervical para el movimiento de golpeo con la pata anterior (Haller, 2014; Siegel, 2005).
El “ataque predador” es controlado desde el hipotálamo lateral y desde regiones del tronco del encéfalo, como es la sustancia gris periacueductal ventrolateral (obsérvese que esta región es distinta de la implicada en el “ataque afectivo”), entre otras. Además, el hipotálamo lateral recibe información excitadora desde la amígdala central y lateral e inhibidora desde la amígdala medial; la conexión entre el hipotálamo lateral y la sustancia gris periacueductal ventrolateral es también excitadora. Desde diversas regiones del tronco del encéfalo se controlan los movimientos de acecho y golpeo así como de mordedura (Haller, 2014; Siegel, 2005).
Ambos circuitos se inhiben entre sí, es decir mientras el gato está realizando un “ataque predador”, que debe ser silencioso para no alertar a la presa, no puede mostrar “ataque afectivo” con manifestaciones como arquearse, piloerección o bufidos que le harían fácilmente detectable por la presa.
¿Qué hay de la influencia de la corteza prefrontal sobre las estructuras límbicas (amígdala e hipotálamo), indicada por otros autores, en los circuitos del comportamiento agresivo en el gato? Esa influencia existe en el gato a través de una conexión multisináptica a través del tálamo (Haller, 2014).
En relación con las estructuras cerebrales implicadas en el control de la conducta agresiva en el ser humano, los estudios se han basado en el comportamiento de seres humanos con daños cerebrales debido a enfermedades (tumores, quistes, rabia), heridas o enfermedades mentales. Por ejemplo, un estudio de Sano, Mayanagi, Sekino, Ogashiwa y Ishijima (1970) en el que se lesionó el llamado hipotálamo posteromedial tuvo éxito en reducir o abolir la agresividad en pacientes violentos. Otros estudios replicaron este hallazgo (Haller, 2014).
Un ejemplo interesante de control hipotálamico de la agresión es el hamartoma hipotalámico (con el nombre de hamartoma se conoce a un grupo de neuronas, glia o manojos de fibras nerviosas que perdieron su camino en el momento de la migración celular embrionaria, ubicándose en estructuras donde normalmente no deberían estar). Un subgrupo de pacientes que desarrollan esta malformación muestra un incremento de la agresión y cuando se retira el hamartoma se abole la agresividad (Ng et al., 2011). Los hallazgos observados con esta malformación muestran que las funciones hipotalámicas están estrechamente ligadas con la agresión en humanos.
Haller (2014) afirma que los mecanismos descritos para el gato en el hipotálamo, sustancia gris periacueductal y otros centros como la amígdala también pueden funcionar de forma similar en humanos con el añadido de la corteza prefrontal como sustrato de los factores psicológicos.
Las estructuras límbicas (amígdala, formación hipocampal, área septal, corteza prefrontal y circunvolución del cíngulo) modulan fuertemente la agresión a través de sus conexiones con el hipotálamo medial y el lateral (Haller, 2014).
La amígdalaLa amígdala se relaciona actualmente con un conjunto de procesos nerviosos como son la cognición social, la regulación de la emoción, el procesamiento de la recompensa y la memoria emocional; también con la detección de las amenazas procedentes del medio ambiente visual o auditivo así como la excitación de respuestas de lucha o huida a través de sus conexiones con estructuras del tronco del encéfalo. Personas con lesión en la amígdala muestran dificultades en reconocer las señales faciales de malestar y tienen dificultades para generar respuestas de miedo condicionadas (Adolphs, 2013; Klumpers, Morgan, Terburg, Stein y van Honk, 2015). Esto es parecido a lo que ocurre en individuos con alta tendencia a la violencia (jóvenes con callosidad emocional; Blair, 2013a; Lozier, Cardinale, VanMeter y Marsh, 2014; psicópatas adultos; Blair, 2013b; Blair et al., 2004; Marsh y Blair, 2008).
A raíz de las observaciones de Kluver y Bucy (1939) sobre lesiones bilaterales de los lóbulos temporales anteriores en macacos se determinó que estas conducían, entre otras modificaciones de la conducta, a un apaciguamiento del animal haciéndolo menos agresivo. En orden a determinar si la pérdida de la agresividad de los monos era debida a lesión cortical o subcortical, Weiskrantz (1956) produjo ablaciones en su grupo experimental en ambas amígdalas y en el polo temporal medial, mientras que el grupo control había recibido lesión en la convexidad temporal inferior o una operación bilateral sin llegar a dañar el tejido cortical o subcortical. Sólo obtuvo modificaciones relacionadas con disminución de la agresión cuando la lesión afectaba al complejo amigdaloide.
En humanos, el complejo amigdaloide se ha subdividido, de acuerdo con criterios de tipos celulares y densidad de dichos tipos, en 4 conjuntos de núcleos denominados laterobasal o basolateral, centromedial o central, masas intercaladas y superficial o cortical; las masas celulares intercaladas, situadas entre el grupo basolateral y el centro medial, son importantes para el control inhibidor de la actividad de la amígdala (Barbas, Zikopoulos y Timbie, 2011; Rosell y Siever, 2015). Posteriormente esta subdivisión se ha confirmado mediante resonancia magnética nuclear (Bzdok, Laird, Zilles, Fox y Eickhoff, 2013). El grupo de núcleos laterobasal es un receptor de información sensorial del tipo exteroceptivo (visual, auditiva, somatosensorial), el grupo centromedial genera respuestas endocrinas, autónomas y somatomotoras pero también recibe información visceral y gustativa y el grupo superficial está ligado al procesamiento de estímulos olfativos. El grupo laterobasal se coactiva con la corteza prefrontal ventromedial (CPFvm), que se piensa que actualiza contingencias de respuesta. La información del grupo laterobasal se envía al grupo centromedial, bien directa o bien indirectamente a través de las masas intercaladas, el cual conecta fundamentalmente con estructuras corticales y subcorticales motoras.
Dado que hemos visto que en el hipotálamo existen dos regiones implicadas en el control de la agresión reactiva y la agresión predadora, ¿existen conexiones entre la amígdala y aquellas regiones? Sí, existen conexiones desde la amígdala a través de dos vías llamadas estría terminal y vía amigdalófuga ventral (Nieuwenhuys, Voogd y van Huijzen, 2009), que conectan a la amígdala con el hipotálamo anterior y con el hipotálamo lateral.
Estudios de neuroimagen estructural de la amígdala y agresión. Algunos estudios han estudiado sujetos sanos, no clínicos, en este caso mujeres, cuyos niveles de agresión no estaban fuera de lo normal (Matthies et al., 2012). La agresión se midió mediante la prueba “Life History of Aggression” y se subdividió el grupo en dos grupos, uno con puntuaciones superiores a la mediana de la cohorte y otro con puntuaciones inferiores a la mediana de la cohorte. El subgrupo más agresivo tenía un volumen de la amígdala, corregido para el volumen total del cerebro, significativamente menor (16-18%) que el volumen del subgrupo menos agresivo. La limitación de este estudio es que se basa sólo en mujeres y se mide, mediante entrevista, la agresión pasada.
En sujetos violentos psicópatas se ha encontrado reducción significativa del volumen de sustancia gris de la amígdala en varios estudios (Ermer, Cope, Nyalakanti, Calhoun y Kiehl, 2012; Tiihonen et al., 2000). En cambio un estudio del mismo grupo de investigación en adolescentes encarcelados con caracteres psicopáticos (Ermer, Cope, Nyalakanti, Calhoun y Kiehl, 2013) no encontró una disminución en el volumen de la amígdala en función de la puntuación en el test correspondiente de medida de la psicopatía. Estos autores no dan una explicación para la diferencia en el volumen amigdalar entre adolescentes y adultos. Boccardi et al. (2011) estudiaron la morfología cortical y amigdalar en sujetos violentos que se podían caracterizar como psicópatas. El volumen global de la amígdala era significativamente mayor en los violentos que en los controles, pero lo más interesante es que la morfología de los distintos núcleos era diferente de la de los controles, observándose tanto aumento como disminución de tejido en diferentes núcleos amigdalinos considerándose este aspecto como más importante que el volumen mayor de la estructura (Boccardi, comunicación personal). En cambio, un estudio previo (Yang, Raine, Narr, Colletti y Toga, 2009) había encontrado disminución en el volumen de la amígdala de sujetos violentos psicópatas. Para Boccardi et al. (2011) la diferencia entre ambos estudios podría encontrarse en la inclusión en el estudio de Yang et al. (2009) de sujetos con trastorno del espectro de la esquizofrenia.
Vemos, por lo tanto, que existe una cierta relación inconsistente entre volumen de la amígdala y la agresión. Para algunos autores esto puede ser debido a que no se haya distinguido entre agresión impulsiva y agresión predadora o instrumental (Pardini, Raine, Erickson y Loeber, 2014; Rosell y Siever, 2015) o a que tal vez sea necesario tener en cuenta la subdivisión en núcleos de la amígdala como han hecho ya algunos estudios (Boccardi et al., 2011; Gopal et al., 2013; Yang et al., 2009; Yoder, Porges y Decety, 2015). Por ejemplo Gopal et al. (2013) estudiaron una población de pacientes psiquiátricos cuya agresión y violencia midieron mediante escalas psicométricas y subdividieron la amígdala en una región dorsal (que incluye el núcleo central) y una región ventral (que incluye el complejo basolateral) midiendo el volumen mediante resonancia magnética nuclear y correlacionando el volumen total y el volumen de las regiones dorsal y ventral con los índices psicométricos de agresión/impulsividad; no obtuvieron una correlación entre el volumen total y los índices de agresión/impulsividad, pero sí una relación positiva entre las regiones ventrales de la amígdala y la agresión/impulsividad. Los autores consideraron que la amígdala dorsal tendría un papel de control y la amígdala ventral uno de activación, dada su conexión con la corteza orbito-frontal que tiene que evaluar e integrar los estímulos que recibe de las áreas corticales sensoriales de asociación y de la propia amígdala. Dicha activación se pondría de manifiesto en los sujetos agresivos por falta de evaluación e integración de la información procedente desde la corteza prefrontal.
Estudios de neuroimagen funcional de la amígdala y agresión. Los estudios de neuroimagen funcional se han basado en diversas técnicas (tomografía de emisión de positrones [TEP] y resonancia magnética funcional [RMf]].
En un estudio con una muestra muy diversa de sujetos asesinos que habían sido declarados inocentes por razones de locura, entre los que había esquizofrénicos, epilépticos y otros, Raine, Buchsbaum y Lacasse (1997), utilizando la técnica TEP, encontraron una asimetría anormal en la actividad de la amígdala, mostrando la amígdala izquierda una actividad reducida y la derecha una actividad aumentada.
Para diferenciar las regiones corticales y subcorticales implicadas en la agresión reactiva frente a la agresión instrumental, Raine et al. (1998) dividieron a los sujetos del estudio previo (Raine et al., 1997), mediante una escala de 4 puntos de violencia predadora-afectiva, en un subgrupo de 15 sujetos que mostraban violencia predadora y 9 sujetos que mostraban violencia afectiva. Se debe tener en cuenta que en este estudio el análisis subcortical incluía el hipocampo, la amígdala, el tálamo y el tronco del encéfalo de forma indiferenciada. Ambos grupos mostraban niveles de glucosa subcortical superiores al grupo control sólo en el hemisferio derecho. Los autores interpretan los hallazgos en el sentido de que la activación del hemisferio derecho interviene en la generación de afecto negativo, lo que daría lugar a una predisposición a la conducta violenta.
Los estudios recientes más extensos de neuroimagen funcional se han realizado generalmente en poblaciones de psicópatas. Estos han demostrado niveles más bajos de actividad en la amígdala cuando ven imágenes que muestran violaciones morales e imágenes con caras de miedo, también durante el condicionamiento aversivo y mientras ven imágenes de estímulos aversivos, así como en una tarea de recuerdo de palabras con contenido emocional frente a otras neutras (Anderson y Kiehl, 2012, 2013). Hay que tener en cuenta que los psicópatas no suelen reaccionar a este tipo de imágenes y no muestran condicionamiento aversivo, por lo que los niveles bajos de actividad de la amígdala estarían en la base de la insensibilidad emocional de estos sujetos.
Los adolescentes con problemas de conducta y rasgos de callosidad emocional muestran menos sensibilidad de la amígdala a caras de miedo, aunque no a otras expresiones faciales, que los sujetos control y que los niños con problemas de conducta pero rasgos de callosidad emocional menos intensos (véanse revisiones en Herpers, Scheepers, Bons, Buitelaar y Rommelse, 2014 y Umbach, Berryessa y Raine, 2015).
Como veremos más adelante, partes de la corteza prefrontal están también asociadas con el comportamiento agresivo, en particular la corteza orbitofrontal y la corteza prefrontal ventromedial. La amígdala está conectada intensamente con estas regiones corticales a través del fascículo uncinado (Catani y Thiebaut de Schotten, 2008; Fortin et al., 2012; Heide, Skipper, Klobusicky y Olson, 2013) que conecta bilateralmente la corteza orbitofrontal y la amígdala mientras que la conexión entre la corteza prefrontal ventromedial y la amígdala es más bien unidireccional en el sentido de aquella a esta.
La integridad estructural del fascículo uncinado (FU) ha sido estudiada en sujetos violentos adultos (Craig et al., 2009; Motzkin, Newman, Kiehl y Koenigs, 2011; Sundram et al., 2012; Wolf et al., 2015) y en adolescentes con trastorno de conducta (Pape et al., 2015; Passamonti et al., 2012; Sarkar et al., 2013 y véase revisión en Olson, Von Der Heide, Alm y Vyas, 2015). En los sujetos adultos se ha encontrado una reducción microestructural del FU derecho–que, por otro lado, es el FU de mayor tamaño de los dos hemisferios y, por ello, tal vez más fácil de cuantificar en la resonancia magnética–comparado con sujetos control de la misma edad y de igual nivel de inteligencia (Craig et al., 2009; Motzkin et al., 2011), aunque esta alteración no está restringida al FU (Sundram et al., 2012) y además está reducción está relacionada con las características interpersonales (encanto superficial, sentido grandioso de lo que uno vale, mentira patológica y manipulación de los otros) de los psicópatas.
En cuanto a los adolescentes violentos, tal conducta se ha observado asociada con una conectividad estructural más elevada que en los sujetos adultos violentos (Pape et al., 2015; Passamonti et al., 2012; Sarkar et al., 2013) y que en jóvenes control de la misma edad y nivel de inteligencia. ¿Qué explicación podría darse a esta diferencia entre sujetos adultos y jóvenes violentos en la microestructura del FU? Se ha planteado que podría deberse a una maduración anormal acelerada y que sería en parte de origen genético y de desarrollo (Passamonti et al., 2012) y cuando estos sujetos adolescentes llegaran a adultos la maduración anormal acelerada explicaría su FU reducido.
La corteza prefrontalLa corteza prefrontal es aquella parte del lóbulo frontal situada delante de la denominada corteza premotora, incluyendo tanto regiones de la parte medial del hemisferio como de la parte lateral. En lo que nos interesa a nosotros para el estudio de la agresión, la corteza prefrontal contiene 3 regiones importantes: la corteza orbitofrontal (COF), la corteza cingulada anterior (CCA) y la corteza prefrontal ventromedial (CPFvm). La COF se encuentra en la parte basal de los hemisferios mientras que la CPFvm se encuentra en la cara medial de los hemisferios y en su parte ventral y la CCA se encuentra en la cara medial de los hemisferios (fig. 1).

A. Vista sagital medial del hemisferio derecho. La parte frontal está a la izquierda y la parte occipital a la derecha; en negro, el cuerpo calloso. B. Visión sagital medial del hemisferio derecho en la que se han numerado las áreas de Brodmann que se citan en el texto. Las líneas corresponden a los límites de las áreas. En rayado discontinuo se encuentran las BA 11,12 y 25 que se consideran parte de la CPFvm. La corteza cingulada anterior forma parte del BA 24. C. Vista basal del hemisferio derecho: OLF, surco olfativo, MOS, surco orbital medio, TOS, surco orbital transversal, LOS, surco orbital lateral. D. Vista basal del lóbulo frontal del hemisferio derecho en la que se muestra parte de la CPFvm en rayado discontinuo. El resto de la base del lóbulo frontal está constituido por las circunvoluciones orbitales u orbitarias: OLF, surco olfativo, SOM, surco orbital medio, SOT, surco orbital transversal, SOL, surco orbital lateral.
La COF se ha subdividido en varias áreas de Brodmann (BA en adelante) y no es uniforme anatómicamente y por lo tanto no lo será desde un punto de vista funcional (Price, 2006, 2007). La COF forma parte de la corteza frontal límbica junto con la CCA. Ambas cortezas, en el caso de la COF sobre todo su zona posterior, tienen las conexiones más fuertes con la amígdala (Ghashghaei, Hilgetag y Barbas, 2007). La vía amígdala-COF posterior puede tener un papel destacado en el enfoque de la atención sobre estímulos motivacionalmente relevantes, consistente con el papel de la amígdala en la alerta y vigilancia emocional. Como la amígdala recibe información sensorial de todas las modalidades sensoriales (que terminan en las mismas partes de la amígdala que se proyectan a la COF posterior), sus proyecciones pueden enviar el significado afectivo de los estímulos sensoriales externos. A su vez, la información que la CCA y la COF enviarían a la amígdala sería acerca del medio interno incluyendo emociones internalizadas como celos, vergüenza y culpabilidad, que evocan una excitación emocional (Ghashghaei et al., 2007).
Dado su patrón de conexiones con la amígdala, existiría una diferencia en las funciones de la COF y la CCA: como la COF recibe más proyecciones de la amígdala y la CCA envía más proyecciones a la amígdala (macacos, Ghashghaei et al., 2007; humanos, Roy et al., 2009), la primera tendría un valor más sensorial o perceptual evaluando el valor motivacional y afectivo de los estímulos mientras que la segunda tendría una relación más importante con las acciones o respuestas (hay que tener en cuenta que la CCA tiene sus principales conexiones de salida con centros autónomos del tronco del encéfalo y de la médula espinal implicados en la expresión de emociones como vocalizaciones) (Rosell y Siever, 2015; Timbie y Barbas, 2014). Es importante destacar que la COF contiene una conexión especializada con la amígdala, a través de las masas intermedias de esta, que activaría centros del tronco del encéfalo y de la médula espinal implicados en la activación emocional y en el retorno a una situación emocional previa disminuyendo dicha activación. Este circuito permitiría explicar la excitación emocional (incremento de frecuencia respiratoria y cardiaca, etc.) en situaciones de comportamiento agresivo.
Estudios de daño cerebral en la CPF y agresión. En la literatura neuropsicológica se han encontrado ejemplos de agresión impulsiva tras daño a la COF ya sea durante la infancia/adolescencia (Anderson, Bechara, Damasio, Tranel y Damasio, 1999; Boes et al., 2011; Eslinger, Flaherty-Craig y Benton 2004; Mitchell, Avny y Blair, 2006) o durante la edad adulta (Bechara, Damasio y Damasio, 2000; Benton, 1991; Berlin, Rolls y Kischka, 2004; Grafman et al., 1996; Hornak et al., 2003; Szczepanski y Knight, 2014) (tabla 1).
Resumen de las regiones de la CPF afectadas en los estudios citados anteriormente (no se citan las revisiones; Benton, 1991; Eslinger, Flaherty-Craig y Benton, 2004; Szczepanski y Knight, 2014)
| Región lesionada | Tipo de neuroimagen | Datos conductuales | |
|---|---|---|---|
| Anderson et al. (1999) | Sujeto A: COFm (BA 11) bilateralmente; polo frontal (BA 10) bilateralmente; CPFdl (BA 9 y 46); COFl (BA 47) izquierda. Sujeto B: polo frontal (BA 10) derecho; BA 8, 9 y 46 medial y lateralmente; COFl (BA 47/12); CCA (BA 24, 32). | Resonancia magnética | Sujeto A: múltiples detenciones por robos; abusaba verbal y físicamente de otros; comportamiento sexual de riesgo; no mostraba culpabilidad ni remordimiento por sus errores; inteligente y académicamente capaz. Sujeto B: generalmente mostraba un afecto neutro; estallidos de ira breves y explosivos; mentía frecuentemente y se implicaba en asaltos físicos. |
| Boes et al. (2011) | CPFvm: (BA 11, 12) CCA (BA 25, 32) | Resonancia magnética | A la edad de 6 años en la escuela: robo, mentira, agresión, rabia y desobediencia. Inteligente y académicamente capaz. Manipulador y controlador. Ataque planeado a su padre (“a sangre fría, sin emoción”). |
| Mitchell et al. (2006) | Polo frontal (BA 10) COF (BA 11) CPFdl (BA 9, 46) | Tomografía axial computarizada (TAC). | Estudio de laboratorio mediante test neuropsicológicos. Déficits en la toma de decisiones. Agresión reactiva |
| Bechara et al. (2000) | Polo frontal (BA 10) COF (BA 11) CPFvm (BA 11, 12) Insula (BA 13) CCA (BA 25, 32) | Resonancia magnética estructural. | La mayoría de los sujetos estudiados eran incapaces de reexperimentar una situación previa de miedo, aunque sí podían hacerlo con una de ira. |
| Berlin et al. (2004) | Polo frontal (BA 10) COF (BA 11, 12) CPFdm (BA 8, 9) CPFdl (BA 9, 46) | Resonancia magnética estructural. Tomografía axial computarizada (TAC). | Agresivos/violentos, airados o irritables. |
| Grafman et al. (1996) | Grupo lesión sólo en COF (COF). Grupo lesión sólo en CPFvm (CPFvm). | Tomografía axial computarizada (TAC). | Test neuropsicológicos: sujetos con lesiones que se restringían o incluían COF eran más violentos (agresión impulsiva). |
| Hornak et al. (2003) | Pacientes orbitales (BA 10, 11, 12, 25). Pacientes mediales (BA 8, 9, 10). Pacientes dorsolaterales (BA 9, 46). | Test neuropsicológicos: lesiones bilaterales de la COF dan lugar a cambios significativos en la conducta social (indicados por alguna persona próxima) y en el estado emocional subjetivo (comparado con antes de la lesión). |
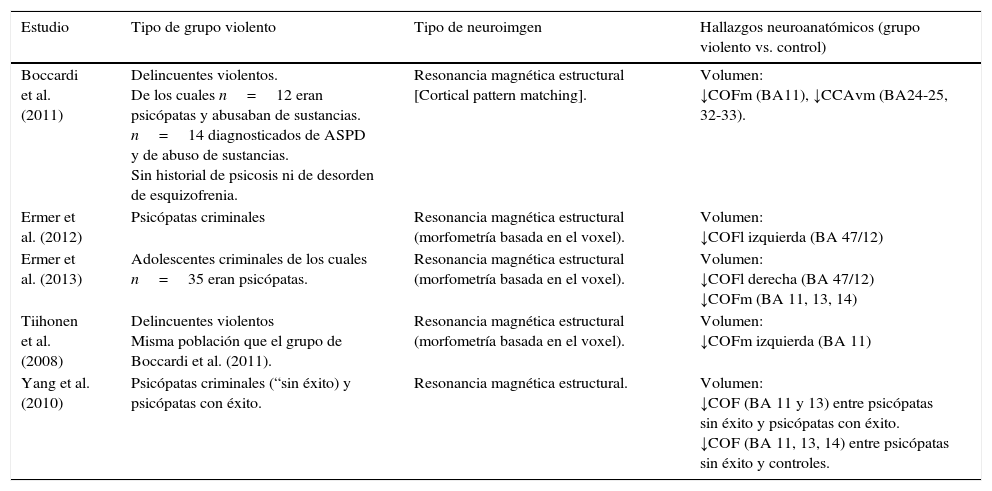
Estudios de neuroimagen estructural de la CPF y agresión. Se han encontrado disminuciones del volumen de sustancia gris en la COF en sujetos adultos de una muestra criminal que mostraba psicopatía (Boccardi et al., 2011; Ermer et al., 2012), en adolescentes varones encarcelados con caracteres psicopáticos (Ermer et al., 2013), en delincuentes violentos (Tiihonen et al., 2008), y en psicópatas sin éxito (que no han podido evitar condenas criminales) (Yang, Raine, Colletti, Toga y Narr, 2010) (tabla 2) (véase también revisión de diversos estudios en Yang, Glenn y Raine, 2008).
Resumen de las regiones de la CPF afectadas en los estudios citados anteriormente
| Estudio | Tipo de grupo violento | Tipo de neuroimgen | Hallazgos neuroanatómicos (grupo violento vs. control) |
|---|---|---|---|
| Boccardi et al. (2011) | Delincuentes violentos. De los cuales n=12 eran psicópatas y abusaban de sustancias. n=14 diagnosticados de ASPD y de abuso de sustancias. Sin historial de psicosis ni de desorden de esquizofrenia. | Resonancia magnética estructural [Cortical pattern matching]. | Volumen: ↓COFm (BA11), ↓CCAvm (BA24-25, 32-33). |
| Ermer et al. (2012) | Psicópatas criminales | Resonancia magnética estructural (morfometría basada en el voxel). | Volumen: ↓COFl izquierda (BA 47/12) |
| Ermer et al. (2013) | Adolescentes criminales de los cuales n=35 eran psicópatas. | Resonancia magnética estructural (morfometría basada en el voxel). | Volumen: ↓COFl derecha (BA 47/12) ↓COFm (BA 11, 13, 14) |
| Tiihonen et al. (2008) | Delincuentes violentos Misma población que el grupo de Boccardi et al. (2011). | Resonancia magnética estructural (morfometría basada en el voxel). | Volumen: ↓COFm izquierda (BA 11) |
| Yang et al. (2010) | Psicópatas criminales (“sin éxito) y psicópatas con éxito. | Resonancia magnética estructural. | Volumen: ↓COF (BA 11 y 13) entre psicópatas sin éxito y psicópatas con éxito. ↓COF (BA 11, 13, 14) entre psicópatas sin éxito y controles. |
Estudios de neuroimagen funcional de la CPF y agresión. Diversos estudios han mostrado, en sujetos normales, una potenciación de la actividad de la COF según se incrementaba la intensidad de expresiones faciales de ira que estaban observando (Blair, Morris, Frith, Perrett y Dolan, 1999). Utilizando un escenario imaginario de agresión en el que se le pedía a sujetos normales que expresaran una agresión no restringida hacia un asaltante o que intentaran inhibir la respuesta agresiva, se observó que en el primer caso la desactivación funcional de la COF era mayor que en el segundo (Pietrini, Guazzelli, Basso, Jaffe y Grafman, 2000). Se observa, por lo tanto, en los sujetos normales que un control de impulsos se traduce en una mayor actividad de la COF. Por lo tanto, sujetos con daño en la COF tenderán a exhibir un bajo control de impulsos y estallidos agresivos entre otras conductas, como se ha indicado en el apartado “Hipotálamo y sustancia gris periacueductal”. Hoptman (2003) cita también estudios realizados en sujetos sanos en los que se ha inducido un estado de ira al que se compara con un estado neutro y se les miden sus cambios en el flujo sanguíneo cerebral regional; en este caso, se observaron incrementos en la COF, CCA y polos temporales anteriores, que los autores interpretaron “como representando la inhibición de la agresión en el contexto de la ira generada” (Hoptman, 2003, p. 269). La cuestión es que en el estudio anterior al que hace referencia Hoptman (2003) no se habían estudiado otras emociones.
¿Qué ocurre funcionalmente en la CPF de los sujetos violentos? Un estudio mediante TEP llevado a cabo en asesinos predadores (psicópatas) e impulsivos y en sujetos normales neurológica y conductualmente (Raine et al., 1998) aportó los siguientes resultados en relación con el tipo de agresión y la actividad de la corteza prefrontal: los asesinos impulsivos tenían menor actividad prefrontal y mayor actividad subcortical en el lóbulo temporal (que contiene la amígdala) que los sujetos control y los asesinos predadores tenían una actividad prefrontal similar a la de los sujetos control pero tenían una actividad subcortical excesiva. En este estudio no se realizaba aún una subdivisión más fina de la corteza prefrontal.
También la revisión llevada a cabo por Hoptmann (2003) mostraba que “la activación en el lóbulo frontal y parte medial del lóbulo temporal está asociada con la agresión imaginada, la inducción de estados de ira o la actividad violenta y criminal, especialmente en relación con crímenes que tenían una base afectiva/impulsiva” (p. 269).
En un meta-análisis en el que se intentó fraccionar la corteza prefrontal en relación con la conducta de sujetos violentos, antisociales y psicópatas, Yang y Raine (2009) encuentran que–en los 31 estudios de imagen funcional que abarcaban desde 1992 a 2007 e incluían las técnicas SPECT (tomografía computarizada de emisión de protón único), RMf (resonancia magnética funcional), espectroscopia de resonancia magnética (MRS), tomografía de emisión de positrones (TEP)–las regiones de la CPF donde había una reducción de la actividad cerebral medida mediante alguna de las técnicas anteriores eran la COF derecha, la CPFdl izquierda y la CCA derecha. La lateralización de la disminución de la actividad cerebral en COF y CCA es paralela a la alteración observada tras lesión en estas regiones en la conducta social, procesamiento emocional y toma de decisiones (Eslinger y Damasio, 1985; Hornak et al., 2003; Tranel, Bechara y Denburg, 2002), mientras que la lateralización en la actividad de la CPFdl estaría relacionada más bien con impulsividad y pobre control conductual.
En general, los estudios de neuroimagen funcional llevados a cabo en sujetos violentos y antisociales muestran una actividad funcional atípica en la COF, la CPFvm y la CCA a menudo manifestada como una hipofuncionalidad (Anderson y Kiehl, 2012; Glenn y Raine, 2014; Patrick, 2015).
La neuroquímica de la agresión impulsiva: la serotoninaAunque otros neurotransmisores y hormonas se han asociado con el comportamiento agresivo, el neurotransmisor más intensamente estudiado ha sido la serotonina o 5-hidroxitriptamina (abreviadamente, 5-HT). La 5-HT es fabricada en los núcleos del rafe del tronco del encéfalo. Estos núcleos contactan con la COF y el sistema límbico entre otros.
Se ha encontrado que los niveles del neurotransmisor 5-HT en regiones corticales como la CPF y la CCA son inversamente proporcionales a los niveles de agresión impulsiva (Coccaro, Fanning, Phan y Lee, 2015; Siegel y Douard, 2011; Yanowitch y Coccaro, 2011).
Utilizando tareas en que los sujetos podían demostrar una conducta agresiva o no agresiva hacia otra persona, varios estudios han mostrado, tanto en hombres como en mujeres, que la reducción mediante la dieta de la 5-HT cerebral está asociada con la conducta agresiva impulsiva (Coccaro et al., 2015; Quadros, Takahashi y Miczek, 2010; Siegel y Douard, 2011) que se muestra ante estímulos nocivos, amenazantes o provocadores. Recíprocamente, si se aumenta la dosis de triptófano en la dieta se observa una irritabilidad y agresión disminuida en estudios experimentales en humanos.
También se ha manipulado farmacológicamente los niveles de 5-HT en el cerebro, por ejemplo mediante la administración de fluoxetina (Prozac), un inhibidor selectivo de la recaptura de 5-HT que aumenta la concentración de esta en el espacio sináptico. Comparada con la administración de un placebo, la administración de fluoxetina originaba una reducción en las puntuaciones de una escala de medición de la agresión, la “Overt Aggression Scale” (Coccaro et al., 2015; Siegel y Douard, 2011). Este dato, junto con el hallazgo original de que los niveles en el líquido céfalo-raquídeo del metabolito principal de la 5-HT, el ácido 5-hidroxiindolacético (5-HIAA), explicaba un porcentaje muy elevado, casi el 61% de la varianza en un grupo de militares diagnosticados con trastornos de personalidad (Brown, Goodwin, Ballenger, Goyer y Major, 1979), condujo a la hipótesis de la deficiencia de serotonina en la agresión humana. Los estudios del grupo de Brown fueron criticados por Berman, Tracy y Coccaro (1997) debido a la ausencia de grupo de comparación, problemas estadísticos y otros. Berman et al. (1997) concluyen que “la idea de que la 5-HT regula la conducta agresiva es apoyada sólo modestamente por los estudios de 5-HIAA en el fluido cerebro-espinal” (p. 655).
Recientemente un meta-análisis ha intentado analizar los estudios que han puesto en relación la 5-HT con la agresión (Duke, Bègue, Bell y Eisenlohr-Moul, 2013). En este meta-análisis se encuentra “que la serotonina explica sólo un poco más del 1% de la varianza en agresión, ira y hostilidad” (Duke et al., 2013, p. 1159). Pero los mismos autores afirman que la hipótesis de la deficiencia de serotonina permanece abierta al debate porque puede haber varias interpretaciones posibles de los resultados obtenidos, desde la poca fiabilidad y validez discriminante de la escala conductual para medir la agresión al conocimiento actual más detallado de las vías nerviosas en las que interviene la serotonina como neurotransmisor y sus receptores, que hacen que el papel de la 5-HT en relación con el control de la conducta sea más complejo de lo que se suponía inicialmente (Bortolato et al., 2013; Coccaro et al., 2015; Duke et al., 2013).
Por otro lado, hay que tener en cuenta que la 5-HT es utilizada como neurotransmisor en diversos circuitos además de la CPF, por ejemplo en el hipotálamo (Hornung, 2003), que es un centro importante relacionado con el control de la agresión como hemos visto anteriormente. Además no se puede determinar qué vías serotoninérgicas contribuyen primariamente a los contenidos de 5-HIAA que se miden en el líquido cefaloraquídeo (Quadros et al., 2010).
La 5-HT tiene al menos 14 tipos de receptores, con una complejidad de localización (pre o postsináptica) y de función (favoreciendo o inhibiendo la agresión) elevada (Bortolato et al., 2013; Coccaro et al., 2015). De ellos, los más importantes en relación con la agresión parecen ser los denominados 5-HT1A y 5-HT1B (Olivier y van Oorschot, 2005; Pavlov, Chistiakov y Chekhonin, 2012; Takahashi, Quadros, de Almeida y Miczek, 2011). Por otro lado, es importante conocer en qué lugares del cerebro son más abundantes los receptores ligados con la agresión: un estudio de TEP (Witte et al., 2009) mostró que la distribución más alta del receptor 5-HT1A, que es un receptor inhibidor y por lo tanto hace que disminuya la transmisión sináptica serotoninérgica, en sujetos con altas puntuaciones en agresión se encontraba en la corteza prefrontal (CPF) y la corteza cingulada anterior (CCA). Por lo tanto, en los sujetos del estudio de Witte et al. (2009), la menor activación de las CPF y CCA haría que estas tuvieran una menor inhibición sobre estructuras del sistema límbico como la amígdala o el hipotálamo, que hemos visto implicadas en el origen de la conducta agresiva. En cambio, se ha encontrado que otro de los receptores de 5-HT, el receptor 5-HT2A, que es un receptor excitador, está asociado con conducta agresiva y con conducta de suicidio (en análisis postmortem en este caso y asociada su localización a la CPF) (Takahashi et al., 2011).
La 5-HT es retirada del espacio sináptico por una proteína que es el transportador de serotonina (5-HTT). Cuando se bloquea el 5-HTT mediante moléculas conocidas como inhibidores selectivos de la recaptura de la serotonina (ISRSs, por ejemplo la fluoxetina), la concentración de 5-HT en el espacio sináptico aumenta y se observa una reducción en la conducta agresiva (Quadros et al., 2010; Takahashi et al., 2011). La proteína 5-HTT es fabricada por un gen cuya región promotora presenta polimorfismo, el cual afecta a la transcripción de dicho gen. Así, existen dos alelos de dicho gen, uno de longitud corta (alelo s) y otro de longitud larga (alelo l). Los individuos s/s y s/l, con menor síntesis de 5-HT, son más ansiosos, depresivos, hostiles y agresivos que los individuos l/l (Takahashi et al., 2011; Teodorovic y Uzelac, 2015), aunque existen numerosas discrepancias en los estudios genéticos que muestran asociación entre el gen 5-HTT y la agresión, ya sea hacia otros o hacia sí mismo (suicidio) (Bortolato et al., 2013), porque tal vez existan interacciones gen-ambiente de tal manera que, por ejemplo, los portadores del alelo s tienen mayor proclividad a la agresión y las ideas suicidas en respuesta a situaciones de stress vividas durante la infancia, adolescencia y comienzo de la edad adulta (Caspi et al., 2003; Conway et al., 2012).
Otro gen que relaciona la 5-HT con la agresión es el gen para el enzima monoaminooxidasa A (MAOA), del cual existe un polimorfismo para baja expresión de MAOA (MAOA-L) y otro para alta expresión de MAOA (MAOA-H) (Dorfman, Meyer-Lindenberg y Buckholtz, 2014; Pavlov et al., 2012; Quadros et al., 2010). Se ha informado de una interacción genotipo-ambiente (maltrato, sucesos traumáticos en los primeros 15 años de vida), de tal manera que los individuos con el polimorfismo MAOA-L y ambiente del tipo indicado anteriormente mostraban mayor propensión a los arrestos criminales, mayor desorden de conducta durante la adolescencia y mayor disposición agresiva que los individuos con polimorfismo MAOA-H o los individuos MAOA-L que no habían estado expuestos al tipo de ambiente indicado arriba (Caspi et al., 2002). Obsérvese que este resultado va en contra de la hipótesis de la deficiencia de serotonina, puesto que los individuos MAOA-L tienen más 5-HT en sus circuitos y tal vez la relación sea que la 5-HT aumenta la ansiedad y hace que estos individuos respondan de manera agresiva ante un reto. Algunos grupos han encontrado una asociación similar de la agresión con el polimorfismo MAOA-H (Dorfman et al., 2014).
ConclusionesLos estudios neurobiológicos sobre la agresión humana han prestado atención sobre todo a estructuras subcorticales como la amígdala y a estructuras corticales como la corteza orbitofrontal, la corteza prefrontal ventromedial y la corteza cingulada. En cambio, para los centros organizadores de la agresión, sobre todo impulsiva, todavía debemos seguir basándonos en estudios llevados a cabo en animales como el gato. Los datos experimentales procedentes de este modelo apuntan a la existencia de centros organizadores en el hipotálamo, tanto para la agresión reactiva como para la instrumental, así como en la sustancia gris periacueductal del tronco del encéfalo. A través de sus conexiones nerviosas con los centros anteriores, la amígdala y las cortezas anteriormente citadas llevan a cabo un control excitador o inhibidor sobre aquellos.
Los estudios revisados no son concluyentes pero sugieren que la conducta violenta podría ser el resultado de la disfunción entre la actividad cortical y la subcortical. En concreto, con respecto a la actividad cortical, las áreas en las que los estudios son más coincidentes se vincularían con un mal funcionamiento de la corteza orbitofrontal, la corteza prefrontal ventromedial y la corteza cingulada anterior, que son estructuras que se relacionan con los componentes cognitivo y emocional de la toma de decisiones. En relación a las estructuras subcorticales que se relacionan con los componentes emocionales de la conducta, la violencia se vincula tradicionalmente y de forma consensuada por la mayoría de los estudios revisados con alteraciones en el funcionamiento de la amígdala. Por consiguiente, la alteración de estas estructuras y los circuitos que las unen podrían relacionarse con la agresión y la violencia, de manera que un gradiente de alteraciones pudiera vincularse con el espectro de las conductas agresivas y violentas. Nuevas investigaciones se tendrán que ir acumulando para que se puedan realizar correspondencias entre los distintos tipos de agresión y las distintas alteraciones en circuitos nerviosos. Estas investigaciones tendrán que ir considerando que la expresión concreta de una conducta agresiva o violenta va a estar propiciada, inhibida o modulada por la sociedad y la cultura donde se exprese tal conducta, lo cual tendrá lugar durante el desarrollo del individuo en una interacción constante con el cerebro del sujeto para poder desarrollar modelos situacionales que permitan la prevención de la conducta violenta de un sujeto en una situación concreta.
El comportamiento agresivo aparece ya durante la infancia y continúa su desarrollo en la adolescencia. Además de los datos ya conocidos sobre desarrollo retrasado de la conducta prefrontal, es de destacar cómo un fascículo que conecta la amígdala con la corteza prefrontal, más concretamente la corteza orbitofrontal y la corteza prefrontal ventromedial, el llamado fascículo uncinado, presenta alteraciones estructurales que pueden estar en la base de algunos de los déficits conductuales de estos sujetos violentos como pueden ser los déficits en invertir el aprendizaje (dejar de responder a los estímulos que antes señalaban recompensa pero que ahora indican castigo), lo cual puede conducir a una perseverancia en la conducta antisocial: robos, asaltos sexuales, etc.
Como muestra de la complejidad del tema tratado está el papel de la serotonina en la conducta violenta. Aunque su relación con la violencia es incuestionable por la convergencia de resultados de investigación, no podemos conocer su función concreta entre otros motivos porque se están descubriendo nuevos receptores de la serotonina que deberán integrarse en el estudio de la violencia. Sin embargo, el núcleo central del consenso científico sobre su papel en la expresión de la violencia ejemplifica la relación entre el cerebro, el ambiente (desarrollo, situaciones de estres, etc.) y la conducta violenta. De esta forma, parece que una persona con un déficit serotoninérgico en su cerebro tiende a interpretar más amenazadoramente los estímulos de su ambiente incrementándose la probabilidad de una respuesta agresiva o violenta. De manera análoga a los estudios de interacción genes/ambiente, el enfoque integrador de todos estos resultados en modelos explicativos y predictivos de la violencia es el reto de investigación al que nos enfrentamos en este campo de estudio.
Conflicto de interesesLos autores de este artículo declaran que no tienen conflicto de intereses.
