




La presente investigación está relacionada con el análisis de coprolitos de roedores registrados en el sitio Huaca Ventarrón ubicado en Lambayeque, considerado como un asentamiento ceremonial de importancia regional y asociado cronológicamente con el Formativo Inicial en los Andes Centrales.
El problema de investigación está asociado con la determinación de las características paleoambientales y paleoclimáticas presentes en el asentamiento prehispánico y a qué tipo de alimentos consumieron los roedores productores de los coprolitos registrados en el depósito A1, en el sitio Huaca Ventarrón.
Para esta labor se utilizaron las metodologías de rehidratación de coprolitos (Callen y Cameron, 1960) y el aislamiento de granos (Weir y Bonavia, 1985), con la finalidad de observar la presencia de células capilares y granos de polen. Es así que se consiguió clasificar taxonómicamente los restos vegetales, agrupar los restos según el hábito de crecimiento, determinar la utilidad que presenta para el ser humano, identificar la comunidad vegetal a la cual se encuentran relacionados los restos y establecer el tipo de paleoambiente y paleoclima en el cual estuvo inmerso el sitio Ventarrón.
This research is related to the analysis of rodent coprolites registered at the Huaca Ventarrón site in Lambayeque, considered a ceremonial settlement of regional importance and chronologically associated with the early Formative Period in the Central Andes.
The research problem is associated with the determination of paleoenvironmental and paleoclimatic characteristics present in the pre-Hispanic settlement and types of food consumed by the rodents who produced the coprolites registered in the A1 deposit at the Huaca Ventarrón site.
Coprolite rehydration (Callen y Cameron, 1960) and isolation of grains methods (Weir y Bonavia, 1985) were used in order to observe the presence of hair cells and pollen grains. This enabled plant debris to be taxonomically classified, remains to be grouped in accordance with growth habit, the utility for humans to be determined, the plant community associated with the remains to be identified and the type of paleoenvironment and paleoclimate at the Ventarrón site to be established.
La investigación se desarrolló como parte de la tesis para la obtención del grado de magister dentro de la maestría en Arqueología Sudamericana, que se desarrolló en la Universidad Nacional de Trujillo (Perú) y la Université de Rennes 1 (Francia). El material proviene del “Proyecto de Investigación Arqueológica Ventarrón-Collud”, bajo la dirección del arqueólogo Ignacio Alva Meneses.
Para estudiar el medioambiente del pasado es posible agenciarse de los macrorrestos botánicos (semillas, carbones vegetales, corteza y fibra) como indicadores de las condiciones medioambientales presentes durante su desarrollo. Además de esta clase de material también es viable aprovechar los microrrestos, como son las células epidérmicas y los granos de polen.
El primero está asociado con la capa superficial de células que reviste todas las partes del cuerpo primario de la planta y tiene como principales características ser de múltiples formas y tener resistencia a la maceración y descomposición (Alonso, 2011), de tal manera que al establecer las particularidades de cada tejido epidérmico vegetal es posible lograr reconocer los grupos taxonómicos al cual corresponden y conseguir determinar cuáles fueron las condiciones medioambientales óptimas para la proliferación de este tipo de flora (Belmonte, Rosello y Rojas, 1988).
Con respecto a los granos de polen, antes se deben conocer algunas de sus importantes particularidades: este tipo de material presenta una gran resistencia a la degradación química y microbiana (Llergo y Ubera 2008); tienen una gran amplitud espacial porque muchas clases de polen se encuentran dispersos en la atmósfera y más aun considerando “la lluvia de polen”, lo cual corresponde a la cantidad de polen mezclado de diversas especies dentro de una comunidad vegetal y que al ser transportado en otras áreas geográficas adyacentes pueden ser o no similares a la vegetación del entorno (Tejero-Diez, Reyes-Salas y Martínez-Hernández, 1988), asimismo, presenta características diagnósticas que son útiles para su identificación taxonómica (Ibarra-Morales y Fernández-Galán 2012). Al complementarse estos dos datos es posible lograr una inferencia acerca de las condiciones medioambientales presentes en la zona.
Estos tipos de microrrestos son posibles de registrarse dentro de los coprolitos (Martínez, Burry y Civalero, 2012), que son heces conservadas en estado orgánico por desecación o mineralización (Araujo y Ferreira, 1997) y sirven como material de referencia para múltiples temáticas, como las relacionadas con la paleodieta, identificación de especies paleobotánicas (Belmonte et al., 1988) y reconstrucción de medios ambientes y/o relación de estacionalidad (Betancourt y Saavedra, 2002, Velásquez, Burry, Mancini y Fugassa, 2010, Martínez y Yagueddú, 2012).
En Huaca Ventarrón, asociado al Formativo Inicial, se registró en la esquina del templo un contexto de basural (depósito A1), el cual estaba compuesto por diversos restos de material vegetal y animal, en donde también se lograron recuperar coprolitos de ratones. Conociendo el potencial del análisis de los restos de coprolitos es que surgieron interrogantes relacionadas con dos temáticas: dieta y medioambiente.
El primero está vinculado con la composición de la dieta de los roedores productores de estos coprolitos, la cual considerando el tipo de contexto donde se registraron se vinculan con dos entornos naturales: vegetales silvestres (presente en el entorno inmediato del sitio) y cultivados (consumidos en el templo de Ventarrón y presentes en el basural), de tal forma que tomando en consideración que estos tipos de animales presentan una digestión menor que los demás mamíferos aumenta la posibilidad de identificar microrrestos de los alimentos consumidos dentro de sus heces.
El segundo guarda relación con la determinación de las características paleoambientales y paleoclimáticas presentes en la zona donde se estableció el asentamiento prehispánico de Huaca Ventarrón, siendo posible gracias a las particularidades geográficas y climáticas que necesitan para desarrollarse cada uno de los organismos vegetales registrados dentro de los coprolitos. Esto permite inferir diversas peculiaridades medioambientales, tales como condiciones climáticas asociadas a la humedad y temperatura, pero todas estas inferencias se verán dirigidas por dos principios fundamentales: el equilibrio ecológico, las condiciones ambientales en las que se desarrollan las plantas, y el actualismo, el comportamiento contemporáneo de la flora actual (Oxman, 2011).
Ante las problemáticas planteadas en la presente investigación es que se fijan como objetivos la identificación de las especies botánicas consumidas por los animales productores de los coprolitos, la determinación del tipo de hábito de crecimiento de los restos vegetales, la utilidad que presentó para el ser humano y la comunidad vegetal a la cual pertenece, y de esta forma lograr establecer las condiciones paleoambientales donde se estableció este asentamiento prehispánico.
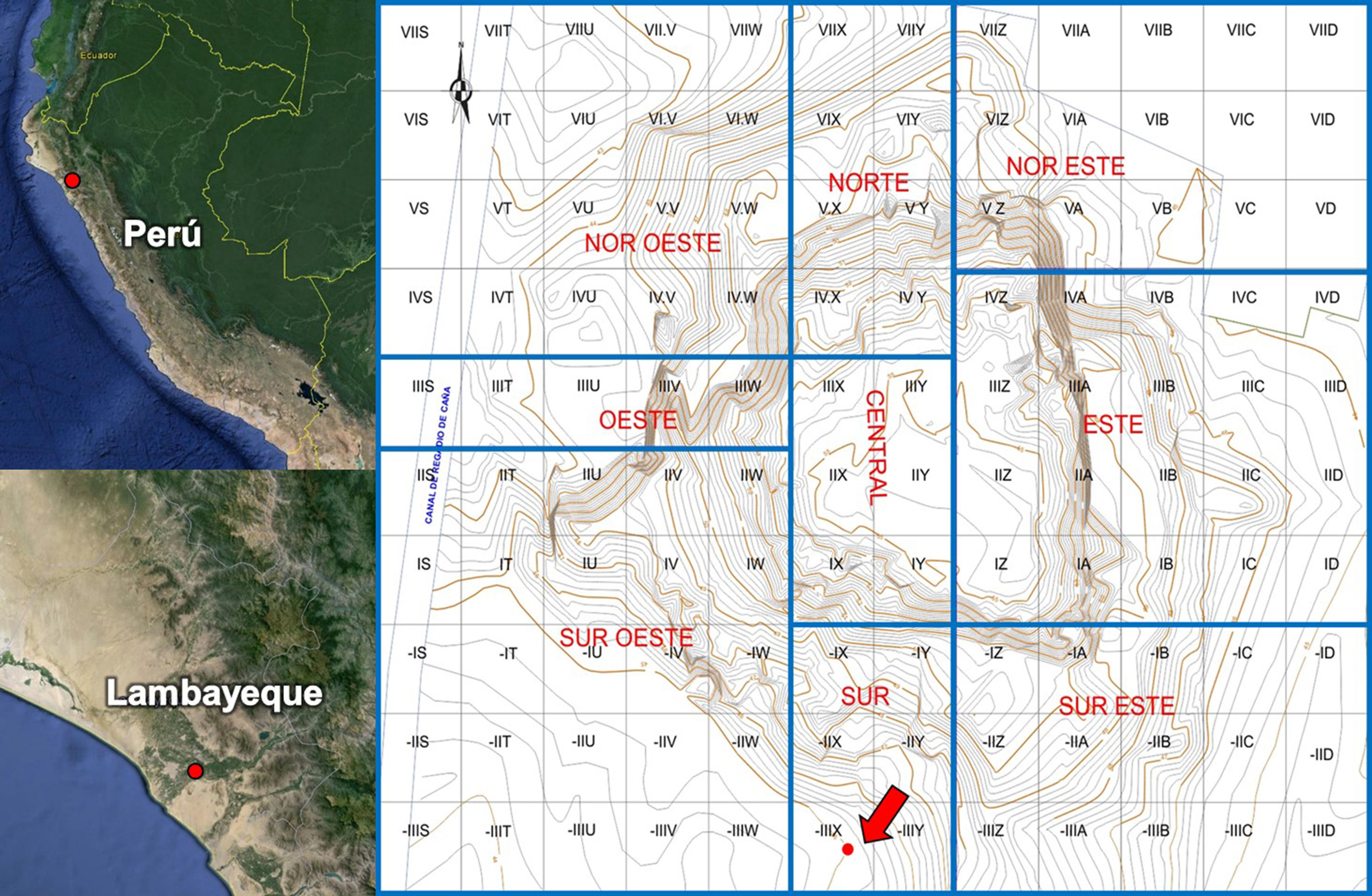
Material y métodosLos materiales provienen del complejo Ventarrón, ubicado en el distrito de Pomalca, provincia de Chiclayo, departamento de Lambayeque (fig. 1). El complejo se encuentra conformado por los sitios denominados Huaca Ventarrón y el conjunto Arenal. Huaca Ventarrón presenta las siguientes características (Alva, 2008, 2012, 2013): El momento de ocupación corresponde al periodo Formativo Inicial (3000 al 1700 a.C.), estando dividido en 9 sectores con un total de 10 fases constructivas. El sitio presenta altorrelieve de seres zoomorfos (peces y zarigüeya) y murales policromos con diversas representaciones, siendo la “cacería de venado”, la pintura mural más antigua de América. Asimismo, se registró un fogón ceremonial, cuyas cenizas al ser datadas arrojaron un fechado de 4000 años antes del presente.
.")
Ubicación política y sectorización del sitio Huaca Ventarrón con la ubicación de la muestra (Alva, 2012, p. 36).
Durante la temporada 2010-2011 (Alva, 2012) se realizó la excavación de la Unidad–III X, localizada en el sector sur y se lograron registrar diversos ambientes (Corredor 1, Recintos 5 y 6) y estructuras (fachada 5, muretes 1 y 2). Al extremo este del Corredor 1 (esquina sureste del templo) entre el murete 1 y el espolón 2 (-III Y) con asociación al segundo piso de la fase siete del templo, se registró el depósito 1A (fig. 2), donde se logró registrar diversos materiales como restos de carbones, material lítico, restos óseos de animales (peces y mamíferos), vegetales (semillas), textiles y coprolitos.
 y los coprolitos de roedores analizados.")
Área de acumulación de basura denominado como depósito 1A en la unidad–III XX, Huaca Ventarrón (Alva, 2013, p. 145) y los coprolitos de roedores analizados.
Este contexto está vinculado con un basural compuesto por los desechos provenientes de las actividades ceremoniales realizadas en la Terraza 3 o el Recinto 5 y presenta una extensión de 1.80m de ancho por 2.70m de largo y 0.20m de espesor. La limpieza de los diferentes ambientes y los desechos producto de las distintas actividades en el templo fueron arrojados durante un tiempo determinado sobre el piso del corredor hasta formar un basural (área de evacuación de desperdicio), lugar de donde los roedores silvestres obtuvieron su alimento. Posteriormente se rellenó el corredor con tierra hasta sellarlo para ser utilizada como base para la construcción del piso 1.
La investigación se encuentra constituida por una población de 332 coprolitos con un peso total de 22.38g. Además, se determinó que según la morfología particular de las heces (pequeño tamaño y cilíndrico), estas se encuentran asociadas con roedores pequeños (ratones) pero debido a la carencia de material de referencia no se logró determinar la especie productora.
Este material se recolectó en campo mediante el método tradicional de excavación en cuadrícula y en estratos naturales. Para el análisis, la totalidad de los coprolitos fueron esparcidos homogéneamente sobre la superficie en tres recipientes de plástico de 12×8cm. Se seleccionaron coprolitos en grupos de cinco de cada uno de los vértices y de la parte central de los recipientes, obteniendo una muestra de 75 coprolitos en total. Los coprolitos restantes se preservaron con la finalidad de tener material para otros tipos de análisis a futuro. El análisis de palinología y arqueobotánica se desarrollarán en el Laboratorio de morfología y fisiología Vegetal de la Facultad de Ciencias Biológicas, en la Universidad Nacional de Trujillo.
MetodologíaEl análisis del material coprolital recuperado durante las excavaciones del sitio Huaca Ventarrón tuvo como objetivo la separación e identificación de los restos vegetales presentes en los coprolitos analizados. Para estos análisis se aplicaron dos metodologías de análisis: la rehidratación de coprolitos y el aislamiento de granos de polen. Antes de hacer el análisis se le asignó un número correlativo a las muestras y se colocaron en frascos de vidrio estériles de 10mL de capacidad. Luego del registro se limpió cuidadosamente la superficie externa de los coprolitos con aire comprimido estéril y chorro de agua destilada, con la finalidad de remover cualquier contaminante extraño. Posteriormente se continuó con los análisis mediantes reactivos químicos.
Se empleó como base el método estándar de rehidratación de coprolitos (Callen y Cameron, 1960) utilizando solución de fosfato trisódico (Na3PO4) al 0.5%, agregando 5mL de esta solución en frascos de vidrio estériles de 10mL hasta cubrirlos completamente. Los recipientes fueron cerrados y las muestras se dejaron remojando por un tiempo mínimo de 72h. Para que los restos se disgreguen fueron agitados periódicamente para que toda la superficie de las muestras sea expuesta al agente químico.
Posteriormente se realizó el homogenizado y el tamizado a través de un cedazo de bronce de 200 Mesh (0.074mm) con la ayuda de chorros de agua destilada, para poder separar los materiales sólidos de la parte líquida que fue recogida por el cedazo, donde se contiene el polen y otros restos microscópicos. Los fragmentos de material más grandes presentes entre los cedazos fueron suavemente removidos para separarlos y liberar cualquier grano de polen que pudiera haber quedado atrapado en la matriz y luego el cedazo fue lavado con chorros de agua destilada.
Luego del tamizado, se logró determinar dos tipos de componentes: los fragmentos grandes que se quedaban en el cedazo y la parte líquida que pasaba el cedazo. Los residuos sólidos más grandes fueron secados en papel de filtro, como paso preparatorio para el análisis. Se continuó el análisis según la metodología empleada por Weir y Bonavia (1985). Los residuos secos fueron separados en las diferentes partes componentes con la ayuda de un microscopio estereoscópico binocular de disección y con pinceles finos. Luego los componentes fueron examinados e identificados y los restos de plantas (células epidérmicas) fueron tabulados para cada muestra de coprolito.
Para el análisis polínico (polen y esporas) de la parte líquida guardada en tubos de ensayo de vidrio de 8cm se continuó con el procedimiento empleado por Weir y Bonavia (1985), el cual consiste en una serie de reactivos químicos para poder aislar los granos de restos del material. Este proceso consistió en un tratamiento con ácido clorhídrico (HCl) al 35% para remover los carbonatos y ácido fluorhídrico (HF) para remover los silicatos. Luego los tubos fueron sellados nuevamente hasta el análisis microscópico. Las muestras fueron montadas en fresco con glicerina utilizando láminas portaobjetos y cubreobjetos. Las variedades de granos conservados fueron identificados y clasificados con un microscopio compuesto Olympus modelo CX31 compuesto con un aumento entre 10x, 40x y 100x. Para la identificación de los microrrestos se utilizaron las colecciones de referencia de polen y tejido vegetal del Laboratorio de Morfología y Fisiología Vegetal y del Herbarium Truxillense, ambos de la Universidad Nacional de Trujillo.
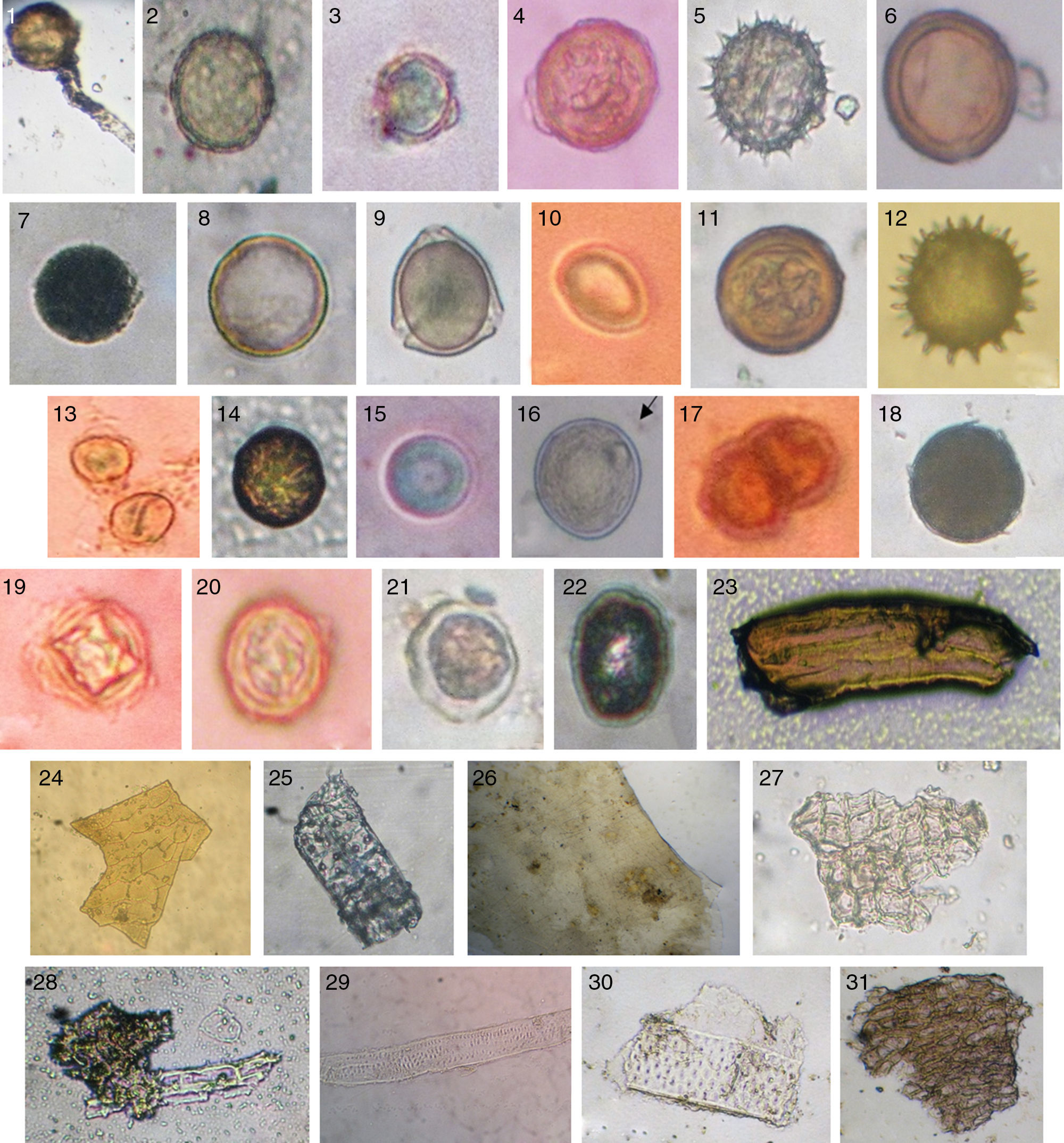
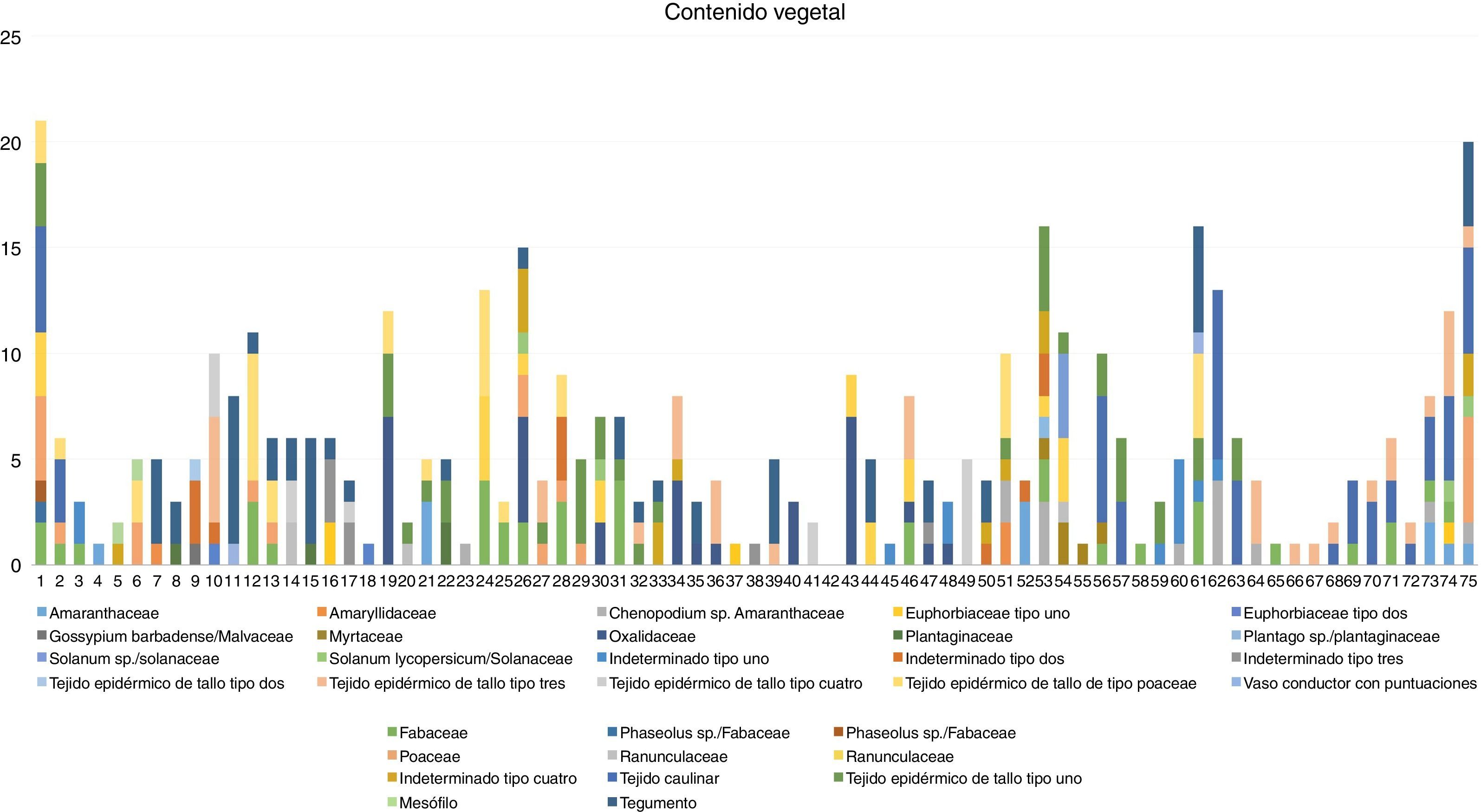
ResultadosEn los 75 coprolitos analizados se logró identificar veintidós tipos granos de polen (diecinueve determinados y tres indeterminados) y trece restos de tejidos vegetales (un resto caulinar, cinco epidérmico de tallo, un vaso conductor, un mesófilo, un tegumento) (fig. 3), formando un total de 212 granos y 221 restos vegetales (Cuadro 1), esto permitió obtener información acerca de especies que posiblemente formaban parte de la dieta y actividades económicas de la población local, de igual manera de los factores ambientales del entorno.
 Amaranthaceae con formación de tubo polínico, 2) Chenopodium sp./Amaranthaceae, 3) Amaryllidaceae, 4) Euphorbiaceae tipo 1, 5) Gossypium barbadense/Malvaceae, 6) Solanum lycopersicum/Solanaceae, 7) Euphorbiaceae tipo 2, 8) Fabaceae, 9) Phaseolus sp./Fabaceae, 10) Myrtaceae, 11) Plantaginaceae, 12) Malvaceae, 13) Oxalidaceae, 14) Plantago sp./Plantaginaceae, 15) Poaceae, 16) Ranunculaceae, 17) Sapotaceae, 18) Solanum sp./Solanaceae, 19) Indeterminado tipo 1, 20) Indeterminado tipo 2, 21) Indeterminado tipo 3, 22) Indeterminado tipo 4, 23) Tejido caulinar, 24) Tejido epidérmico de tallo tipo 1, 25) Tejido epidérmico de tallo tipo 2, 26) Tejido epidérmico de tallo tipo 3, 27) Tejido epidérmico de tallo tipo 4, 28) Tejido epidérmico de tallo de Poaceae, 29) Vaso conductor con puntuaciones, 30) Mesófilo y 31) Tegumento.")
Diversos granos de polen identificados en 40x: 1) Amaranthaceae con formación de tubo polínico, 2) Chenopodium sp./Amaranthaceae, 3) Amaryllidaceae, 4) Euphorbiaceae tipo 1, 5) Gossypium barbadense/Malvaceae, 6) Solanum lycopersicum/Solanaceae, 7) Euphorbiaceae tipo 2, 8) Fabaceae, 9) Phaseolus sp./Fabaceae, 10) Myrtaceae, 11) Plantaginaceae, 12) Malvaceae, 13) Oxalidaceae, 14) Plantago sp./Plantaginaceae, 15) Poaceae, 16) Ranunculaceae, 17) Sapotaceae, 18) Solanum sp./Solanaceae, 19) Indeterminado tipo 1, 20) Indeterminado tipo 2, 21) Indeterminado tipo 3, 22) Indeterminado tipo 4, 23) Tejido caulinar, 24) Tejido epidérmico de tallo tipo 1, 25) Tejido epidérmico de tallo tipo 2, 26) Tejido epidérmico de tallo tipo 3, 27) Tejido epidérmico de tallo tipo 4, 28) Tejido epidérmico de tallo de Poaceae, 29) Vaso conductor con puntuaciones, 30) Mesófilo y 31) Tegumento.
Contenido vegetal y totalidad de restos identificados en los coprolitos analizados
| Contenido vegetal | N.o | Total | |
|---|---|---|---|
| Granos de polen | Amaranthaceae | 11 | 212 |
| Amaryllidaceae | 3 | ||
| Chenopodium sp. Amaranthaceae | 14 | ||
| Euphorbiaceae tipo 1 | 4 | ||
| Euphorbiaceae tipo 2 | 2 | ||
| Fabaceae | 38 | ||
| Phaseolus sp./Fabaceae | 1 | ||
| Malvaceae | 1 | ||
| Gossypium barbadense/Malvaceae | 1 | ||
| Myrtaceae | 5 | ||
| Oxalidaceae | 33 | ||
| Plantaginaceae | 4 | ||
| Plantago sp./Plantaginaceae | 1 | ||
| Poaceae | 19 | ||
| Ranunculaceae | 4 | ||
| Sapotaceae | 20 | ||
| Solanum sp./Solanaceae | 4 | ||
| Solanum lycopersicum/Solanaceae | 4 | ||
| Indeterminado tipo 1 | 12 | ||
| Indeterminado tipo 2 | 11 | ||
| Indeterminado tipo 3 | 7 | ||
| Indeterminado tipo 4 | 13 | ||
| Restos de tejido vegetal | Tejido caulinar | 51 | 221 |
| Tejido epidérmico de tallo tipo 1 | 37 | ||
| Tejido epidérmico de tallo tipo 2 | 1 | ||
| Tejido epidérmico de tallo tipo 3 | 34 | ||
| Tejido epidérmico de tallo tipo 4 | 13 | ||
| Tejido epidérmico de tallo de Poaceae | 28 | ||
| Vaso conductor con puntuaciones | 2 | ||
| Mesófilo | 2 | ||
| Tegumento | 53 | ||
| Total | 433 | ||
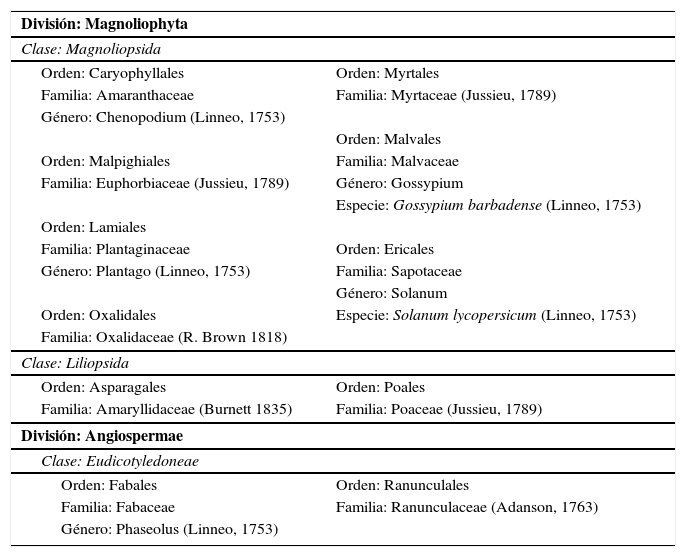
En el análisis se logró registrar granos de polen y restos de tejido vegetal, y debido a las características de conservación de los restos se alcanzó clasificar considerando un orden sistemático y taxonómico (APGII, 2003) en dos divisiones, tres clases, once órdenes, once familias, cinco géneros y dos especies (Cuadro 2). Algunos restos no lograron ser determinados debido al estado de conservación de estos; se ordenaron de la siguiente manera: indeterminado tipo 1, indeterminado tipo 2, indeterminado tipo 3, indeterminado tipo 4, tejido caulinar, tejido epidérmico de tallo tipo 1, tejido epidérmico de tallo tipo 2, tejido epidérmico de tallo tipo 3, tejido epidérmico de tallo tipo 4, tejido epidérmico de tallo de Poaceae, vaso conductor con puntuaciones, mesófilo y tegumento.
Clasificación sistemática y taxonómica de los restos vegetales registrados en los coprolitos
| División: Magnoliophyta | |
| Clase: Magnoliopsida | |
| Orden: Caryophyllales | Orden: Myrtales |
| Familia: Amaranthaceae | Familia: Myrtaceae (Jussieu, 1789) |
| Género: Chenopodium (Linneo, 1753) | |
| Orden: Malvales | |
| Orden: Malpighiales | Familia: Malvaceae |
| Familia: Euphorbiaceae (Jussieu, 1789) | Género: Gossypium |
| Especie: Gossypium barbadense (Linneo, 1753) | |
| Orden: Lamiales | |
| Familia: Plantaginaceae | Orden: Ericales |
| Género: Plantago (Linneo, 1753) | Familia: Sapotaceae |
| Género: Solanum | |
| Orden: Oxalidales | Especie: Solanum lycopersicum (Linneo, 1753) |
| Familia: Oxalidaceae (R. Brown 1818) | |
| Clase: Liliopsida | |
| Orden: Asparagales | Orden: Poales |
| Familia: Amaryllidaceae (Burnett 1835) | Familia: Poaceae (Jussieu, 1789) |
| División: Angiospermae | |
| Clase: Eudicotyledoneae | |
| Orden: Fabales | Orden: Ranunculales |
| Familia: Fabaceae | Familia: Ranunculaceae (Adanson, 1763) |
| Género: Phaseolus (Linneo, 1753) | |
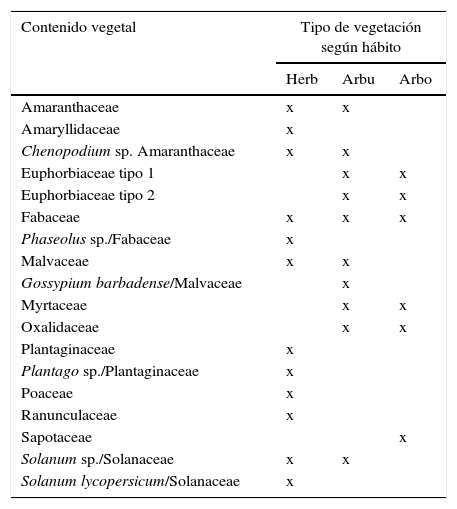
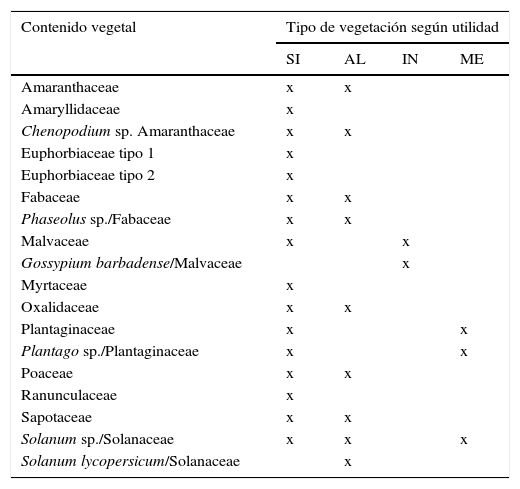
La presencia de restos vegetales en los 75 coprolitos analizados fue relativamente alta, por lo cual es posible determinar que los coprolitos contenían menor cantidad de granos de polen en comparación con los que presentaron restos de tejido vegetal (gráfico 1); sin embargo, los granos de polen mostraron mejor estado de conservación. Debido al bajo estado de conservación de los restos de tejido vegetal, fue difícil identificar el grupo taxonómico al cual estos pertenecían. Los coprolitos 53 (16), 61 (16) y 75 (20) fueron aquellos que presentaron mayor número de restos vegetales. Asimismo, los coprolitos 18, 23, 37, 38, 45, 58, 65, 66 y 67 fueron los que menor número de restos presentaron con tan solo uno por coprolito. Los restos que se encontraron en mayor cantidad considerando todos los coprolitos fueron: resto de tejido caulinar, resto de tegumento, granos de polen de Fabaceae, resto de tejido epidérmico de tallo tipo 1 y granos de polen de Oxalidaceae con un total de 53, 51, 38, 37 y 33 respectivamente. Según el grupo dentro del cual fueron identificados los restos vegetales, estos fueron clasificados, de acuerdo al hábito de crecimiento: doce herbáceas, diez arbustivas y seis arbóreas (Mostacero, Mejía y Gamarra, 2009) (Cuadro 3). Igualmente se clasificaron según la posible utilidad de los restos identificados (Mostacero et al., 2009), logrando determinar dieciséis vegetales silvestres, nueve alimenticias, dos industriales y tres medicinales (Cuadro 4).

Tipo de vegetación según el hábito y cantidad del contenido vegetal encontrado en los 75 coprolitos analizados: Herb, Arbu, Arbo
| Contenido vegetal | Tipo de vegetación según hábito | ||
|---|---|---|---|
| Herb | Arbu | Arbo | |
| Amaranthaceae | x | x | |
| Amaryllidaceae | x | ||
| Chenopodium sp. Amaranthaceae | x | x | |
| Euphorbiaceae tipo 1 | x | x | |
| Euphorbiaceae tipo 2 | x | x | |
| Fabaceae | x | x | x |
| Phaseolus sp./Fabaceae | x | ||
| Malvaceae | x | x | |
| Gossypium barbadense/Malvaceae | x | ||
| Myrtaceae | x | x | |
| Oxalidaceae | x | x | |
| Plantaginaceae | x | ||
| Plantago sp./Plantaginaceae | x | ||
| Poaceae | x | ||
| Ranunculaceae | x | ||
| Sapotaceae | x | ||
| Solanum sp./Solanaceae | x | x | |
| Solanum lycopersicum/Solanaceae | x | ||
Arbo: arbórea; Arbu: arbustiva; Herb: herbácea.
Tipo de vegetación según la utilidad (SI, AL, IN, ME) y cantidad del contenido vegetal encontrado en los 75 coprolitos analizados
| Contenido vegetal | Tipo de vegetación según utilidad | |||
|---|---|---|---|---|
| SI | AL | IN | ME | |
| Amaranthaceae | x | x | ||
| Amaryllidaceae | x | |||
| Chenopodium sp. Amaranthaceae | x | x | ||
| Euphorbiaceae tipo 1 | x | |||
| Euphorbiaceae tipo 2 | x | |||
| Fabaceae | x | x | ||
| Phaseolus sp./Fabaceae | x | x | ||
| Malvaceae | x | x | ||
| Gossypium barbadense/Malvaceae | x | |||
| Myrtaceae | x | |||
| Oxalidaceae | x | x | ||
| Plantaginaceae | x | x | ||
| Plantago sp./Plantaginaceae | x | x | ||
| Poaceae | x | x | ||
| Ranunculaceae | x | |||
| Sapotaceae | x | x | ||
| Solanum sp./Solanaceae | x | x | x | |
| Solanum lycopersicum/Solanaceae | x | |||
AL: alimenticio; IN: industrial; ME: medicinal; SI: silvestre.
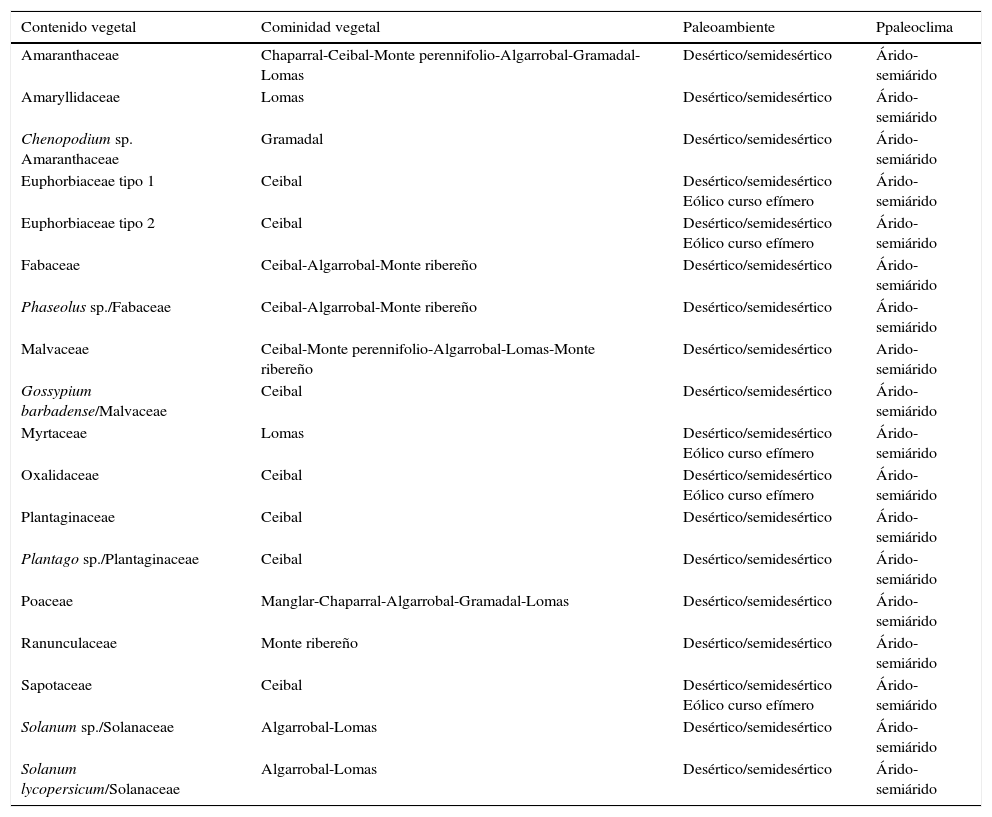
Considerando las regiones ecológicas donde se desarrollan los grupos taxonómicos identificados y la zona de origen de los coprolitos evaluados, se infirió la comunidad vegetal de costa peruana (Ferreyra, 1983), el paleoambiente y paleoclima (Grill, 1997) (Cuadro 5). Se tiene siete comunidades vegetales, tales como chaparral, ceibal, monte perennifolio y ribereño, algarrobal, gramadal y lomas. Se logró establecer dos paleoambientes como desértico/semidesértico, eólico curso efímero; y un paleoclima como el árido-semiárido.
Comunidad vegetal, paleoambiente y paleoclima de los restos vegetales hallados en los 75 coprolitos analizados
| Contenido vegetal | Cominidad vegetal | Paleoambiente | Ppaleoclima |
|---|---|---|---|
| Amaranthaceae | Chaparral-Ceibal-Monte perennifolio-Algarrobal-Gramadal-Lomas | Desértico/semidesértico | Árido-semiárido |
| Amaryllidaceae | Lomas | Desértico/semidesértico | Árido-semiárido |
| Chenopodium sp. Amaranthaceae | Gramadal | Desértico/semidesértico | Árido-semiárido |
| Euphorbiaceae tipo 1 | Ceibal | Desértico/semidesértico Eólico curso efímero | Árido-semiárido |
| Euphorbiaceae tipo 2 | Ceibal | Desértico/semidesértico Eólico curso efímero | Árido-semiárido |
| Fabaceae | Ceibal-Algarrobal-Monte ribereño | Desértico/semidesértico | Árido-semiárido |
| Phaseolus sp./Fabaceae | Ceibal-Algarrobal-Monte ribereño | Desértico/semidesértico | Árido-semiárido |
| Malvaceae | Ceibal-Monte perennifolio-Algarrobal-Lomas-Monte ribereño | Desértico/semidesértico | Arido-semiárido |
| Gossypium barbadense/Malvaceae | Ceibal | Desértico/semidesértico | Árido-semiárido |
| Myrtaceae | Lomas | Desértico/semidesértico Eólico curso efímero | Árido-semiárido |
| Oxalidaceae | Ceibal | Desértico/semidesértico Eólico curso efímero | Árido-semiárido |
| Plantaginaceae | Ceibal | Desértico/semidesértico | Árido-semiárido |
| Plantago sp./Plantaginaceae | Ceibal | Desértico/semidesértico | Árido-semiárido |
| Poaceae | Manglar-Chaparral-Algarrobal-Gramadal-Lomas | Desértico/semidesértico | Árido-semiárido |
| Ranunculaceae | Monte ribereño | Desértico/semidesértico | Árido-semiárido |
| Sapotaceae | Ceibal | Desértico/semidesértico Eólico curso efímero | Árido-semiárido |
| Solanum sp./Solanaceae | Algarrobal-Lomas | Desértico/semidesértico | Árido-semiárido |
| Solanum lycopersicum/Solanaceae | Algarrobal-Lomas | Desértico/semidesértico | Árido-semiárido |
Mediante el análisis de polen y células capilares presentes en los coprolitos en Ventarrón se logró determinar la dieta de los roedores, la utilidad de este tipo de material como indicador medioambiental y las condiciones tanto paleoclimáticas como paleoambientales.
En los coprolitos se logró determinar dos especies vegetales en particular (Solanum lycopersicum y Gossypium barbadense) y dos géneros vegetales (Chenopodium y Phaseolus) vinculados con plantas consumidas por el ser humano y utilizadas para la industria. Producto de las condiciones de conservación de algunos granos de polen y células capilares solamente fue posible identificarlas hasta el nivel de familia, y su asociación con vegetación silvestre y medicinal se basó en la existencia de especies registradas que son empleadas para este fin y que pertenecen a las familias identificadas.
La combinación alimenticia entre plantas silvestres y cultivadas es coherente con el tipo de contexto en donde se registró, así como el comportamiento natural del roedor pues al estar en contacto directo con el ser humano se genera un estado de comensalismo, en el cual los ratones sacan beneficio del hombre mediante el consumo de sus alimentos o desperdicios, asimismo, ocupando las estructuras con la finalidad de lograr obtener un refugio y calor. En estos contextos de abundancia, los ratones se inclinan a seleccionar una dieta nutricionalmente balanceada cuando se le da a elegir entre un amplio rango de alimentos diferentes (Benavides y Guénet, 2003). Este comportamiento se encuentra completamente evidenciado porque el tipo de contexto donde se registraron los coprolitos es de un depósito de basura que contenía restos orgánicos provenientes de las áreas internas del templo (ambientes) y de las actividades dentro de la misma (ceremonias y mantenimiento) (Alva, 2012). Los desperdicios que alimentaban a estos roedores se encuentran formados por especies domésticas como Persea americana (palta), Capparis sp. (sapote), Cucurbita maxima (zapallo), Zea mays (maíz), entre otras especies (Vásquez y Rosales, 2012).
Gracias a los resultados obtenidos de los análisis de granos de polen y tejido vegetal es posible agregar a la variada dieta del poblador en Ventarrón el consumo del tomate (Solanum lycopersicum) que por tener altos niveles de agua en su composición es rápidamente degradable, siendo este el principal problema para su conservación en el registro arqueológico. La presencia de esta fruta está relacionada con dos posibles escenarios: por el consumo de flores y tallos de este vegetal por los roedores o por la ingesta de otros vegetales con polen de tomate impregnado; similar situación se encuentra en el algodón (Gossypium barbadense), el cual corresponde al tipo pardo (nativo). Se encuentran registrados diversos textiles elaborados con algodón en el sitio Huaca Ventarrón y al ser una especie característica de la costa del Perú su presencia en el contexto arqueológico es abundante y desde épocas tempranas, tal como los textiles registrados en Huaca Prieta (Bird, Hyslop y Skinner, 1985), hasta los periodos posteriores (Roque, Cano y Cook, 2003, Millaire y Surette, 2011).
En cuanto a la identificación del tomate, se tiene muy poca presencia de su registro a nivel de especie; esto se debe a lo difícil de su identificación y al estado de conservación del polen. El tomate forma parte de una familia muy amplia como son las solanáceas y cuyos miembros son muy diversificados, como la maleza, papa, tomate, plantas silvestres, entre otros. La presencia de esta familia se encuentra evidenciada en el registro paleopalinológico prehispánico proveniente del análisis de coprolitos de diversos animales, como en el sitio precerámico en Huarmey (Weir y Bonavia, 1985). Los datos del Solanum lycopersicum se remontan a las evidencias en Paloma, donde fueron registradas con un fechado aproximado de 5316 y 3630 a.C (Weir y Dering, 1986), de igual manera se determinó que en el sitio Casa Vieja, asociado con ocupación Nazca, parte de la dieta estaba conformada por el tomate (Cook y Parrish, 2005). Parece ser que este fruto no fue un cultivo muy divulgado durante la época prehispánica pero sí muy bien conocido por el poblador prehispánico ya que según se registra en 1578 después de ser azotados por un evento ENSO, se consumió el tomate como alimento de emergencia (León, 2013).
Aunque dos importantes géneros fueron identificados dentro de los coprolitos, como son Chenopodium sp. y Phaseolus sp., no se lograron determinar qué nivel de especie es posible asociar a las especies que la conforman. El primero está relacionado con la cañihua (Chenopodium pallidicaule) y quinua (Chenopodium quinoa), que presentan un alto nivel alimenticio y fueron alimentos difundidos en toda el área andina, siendo la cañihua una especie cultivada en zonas alto andinas y su registro en la costa sería relacionado con la redes de intercambios prehispánicas como lo evidencia su presencia en el sitio de Galindo (Lockard, 2005). La quinua es un cultivo más propagado ya que puede crecer desde el nivel del mar hasta 3800msnm (FAO, 2011), siendo fácilmente sembrada en áreas cercanas del sitio o en la región. Su presencia en el registro arqueológico es muy variada y se tiene desde el sitio lítico de Quebrada de las Pircas 1, costa norperuana, hasta en el asentamiento inca de Machu Picchu (León, 2013). Otro cultivo de importancia sería el género Phaseolus, que abarca el pallar (Phaseolus lunatus) y fréjol (Phaseolus vulgaris); son legumbres con un elevado valor nutricional y de una importancia transcendental en las diversas sociedades en los andes centrales. Asociado con el contexto donde se recuperaron los coprolitos, se logró identificar restos de fréjol (Vásquez y Rosales 2012), quedando plenamente confirmada su presencia en el sitio de Huaca Ventarrón. La presencia de esta planta no solamente se evidencia en esta región sino que presenta un amplio consumo que se remonta desde épocas muy tempranas, como en la ocupación lítica en Guitarrero (8200-7800 a.C.), pasando por múltiples periodos culturales (Sociedades formativas, Nasca, Moche, Wari, Chimú, Inca) hasta la invasión hispánica (León, 2013).
Los restos orgánicos presentes en el basural fueron complementados con vegetación silvestre del entorno como parte de su dieta y a pesar de no lograr determinar la especie específica de cada resto fue posible establecer asociaciones con las familias identificadas, como árboles (Sapotaceae), arbustos (Oxalidaceae) y hierbas (Amaryllidaceae). Lo antes mencionado corresponde a un escenario típico de poblaciones que cuentan con basurales cerca de las áreas de ocupación, en el cual los desperdicios de sus actividades son recolectados y colocados en un área específica en las inmediaciones de las zonas de donde procede el material. En Ventarrón el área de evacuación estuvo en la parte posterior del templo, lugar donde se depositaban los desperdicios producidos en la estructura principal y que era constantemente circulado por los encargados de brindar el mantenimiento a estos recintos sagrados. Se produjo una relación de comensalismo con los roedores silvestres del entorno, quedando plasmado en la presencia de diferentes tipos de vegetales que consumieron.
Con respecto al clima presente durante la ocupación del sitio Huaca Ventarrón, se logró establecer que, según el tipo de flora identificada, correspondería a un paleoclima árido-semiárido. En la actualidad la región de Lambayeque presenta este tipo de clima con precipitación media anual de los 100mm, lluvias esporádicas en verano y con una alta frecuencia de sequías, pero esto varia como en la mayoría de las áreas costeras, durante la presencia de los eventos “ENSO”, que puede generar abundantes daños a las poblaciones establecidas pero son altamente beneficiosos en la regeneración del bosque seco (López, 1986). De tal manera se puede establecer que los pobladores del sitio Huaca Ventarrón afrontaron similares condiciones climáticas que las actuales, de este modo el tipo de clima tiene una permanencia mayor a los tres mil años, esto considerando la datación de 2300-2035 a.C. que tiene la primera fase constructiva registrada en el sitio. La variación climática en el área andina se encuentra ampliamente investigada, prueba de ello es que se identificó en el registro polínico en múltiples áreas de Sudamérica que cerca de los cuatro mil años comenzó la estabilidad climática y aproximadamente hace tres mil años la vegetación es similar a la actual (Mc Glone, Kersiiaw y Markgraf, 1992, Villagrán, 1993).
Con relación al paleoambiente en donde se asentó la población de Huaca Ventarrón, se consideró que, a pesar de la cantidad de granos de polen, fue menor en comparación con los que presentaron restos de tejido vegetal; sin embargo, todos los coprolitos presentaron restos vegetales, lo cual indicaría la presencia de un nivel de riqueza vegetal relativamente alto en la zona adyacente. Asimismo, la totalidad de los grupos taxonómicos identificados se desarrollan en ambientes xéricos (desértico/semidesértico), lo cual concuerda con las condiciones ambientales actuales en la zona y corrobora la información paleoclimática establecida. De igual manera se observó presencia de polen de familias que incluyen especies arbóreas, y ello indicaría corrientes eólicas en la zona, siendo los fuertes vientos característicos del sitio.
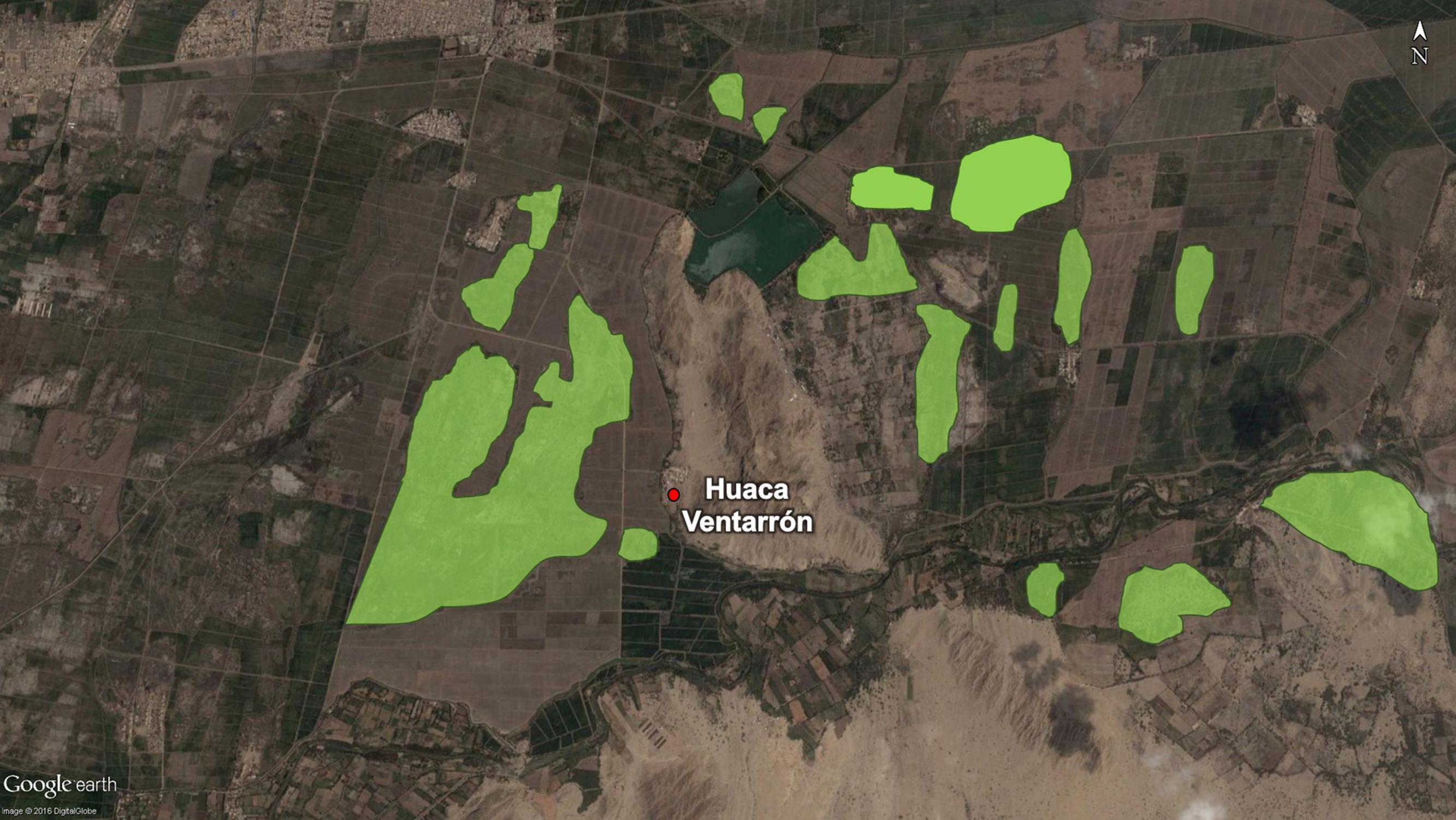
Lamentablemente alrededor del sitio se conserva muy poca flora natural en la actualidad, siendo los campos de cultivos los que predominan en el paisaje (fig. 4). Según el tipo de vegetación identificada y las características tanto paleoambientales como paleoclimáticas, se puede establecer que durante el desarrollo pleno de sus actividades, el entorno estuvo dominado por hierbas, arbustos y vegetación leñosa natural, todos asociados con flora nativa como el algarrobo (Prosqpis paluda), sapote (Capparis angulata), espino (Acacia macracantha), bichayo (Capparis oval/folia), palo santo (Bursera graveolens), entre otros. Un entorno similar a las condiciones identificadas en Huaca Ventarrón sería el Santuario Histórico Bosque de Pómac, en donde es posible reconocer los diversos paleoambientes determinados en la presente investigación (Angulo, 2013, SENARP, 2011), quedando claramente corroborada la relación cercana que tuvieron con su ambiente natural las diversas poblaciones asentadas en la región desde épocas inmemoriales, ya que no solamente fue utilizada para explotar diversos tipos de recursos sino también como una fuente inspiradora para construir un imaginario sólido que luego materializarían en la elaboración de su variada cultura material.

La evidencia directa e indirecta de presencia de ratones es una útil herramienta para el estudio paleoambiental, como es el caso de las paleomadrigueras o “packrat middens” (Anderson y van Devender, 1991), estos contextos son acumulaciones de diversos tipos de materiales (plantas comestibles, microrrestos vegetales, espinas de cactus, restos de insectos, pequeños guijarros y las heces) que el roedor emplea en la construcción y/o utilización de la madriguera. Al pasar miles de años la acumulación se compacta convirtiéndose en una muy buena fuente de información paleoecológica (Fisher, Cole y Anderson, 2009; Maldonado, Betancourt, Latorre y Villagran, 2005), esto es aprovechado en la actualidad para realizar estudios paleoambientales basados en diversos tipos de indicadores (Elias, Mead y Agenbroad, 1992; Richardson y Meyer, 2012). A pesar de no registrarse en la costa norte del Perú estos contextos, es posible establecer la validación del uso de los coprolitos de ratones como indicadores medioambientales; si bien los coprolitos no fueron registrados dentro de una madriguera sino asociados a un área de acumulación de basura se logró determinar que los roedores se alimentaban de estos desechos y a su vez de vegetación silvestre. De igual forma queda completamente descartado asociar la presencia de estos coprolitos con factores tafonómicos o intrusivos ya que durante el proceso de excavación no se registró ningún indicador que se asocie con este comportamiento y todo este material se asocia estratigráficamente con la formación del basural.
Considerando los estados de conservación registrados en el polen y los tejidos vegetales, lo cual dificultó la identificación de los grupos taxonómicos, es posible establecer que el contexto donde se ubicaron los coprolitos ha estado expuesto a condiciones desfavorables, como baja humedad relativa y altas temperaturas, apoyando a su degradación de los microrrestos vegetales.
ConclusionesLos resultados muestran la gran utilidad de los coprolitos como registros de la vegetación del pasado y como indicadores paleoambientales, ya que se logró obtener información acerca de la vegetación y el tipo de clima presente en la zona, así también, de especies, géneros y familias que muy probablemente formaban parte de la dieta de los roedores (también del poblador de Ventarrón), las actividades económicas de la población local y de las comunidades vegetales aledañas.
Asimismo, se logró determinar que durante la ocupación de este asentamiento el paisaje era muy diferente al actual, con un entorno de bosque seco y de similares condiciones medioambientales a las actuales, demostrando claramente una estabilidad climática desde hace alrededor tres mil años.
Al director del Proyecto de Investigación Arqueológica Ventarrón–Collud, Ignacio Alva Meneses, quien me brindó todas las facilidades para realizar el análisis de los coprolitos de roedores. A los doctores Santiago Uceda Castillo y Ramiro J. March por sus esfuerzos para desarrollar la maestría y abrir los horizontes de muchos jóvenes investigadores. A mis colegas Grace Ramírez Zavaleta y Susy Esquivel Valverde por su paciencia y apoyo al revisar el presente artículo.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.