

Introduction
Atrial fibrillation (AF) is to date the most common sustained cardiac arrhythmia in humans and is predicted to dramatically increase its prevalence in the near future.1 Antiarrhythmic drugs are only partially effective in treating AF, and have the potential for serious side effects, including life-threatening pro-arrhythmia. On the other hand, it has recently been demonstrated that paroxysmal AF in patients is initiated by focal triggers localized usually to one of the pulmonary veins (PVs)2 and can be cured by a catheter-based ablation procedure.3 However, in persistent AF, the prevailing theory is that multiple random wavelets of activation coexist to create a chaotic cardiac rhythm,4 and therapy is more challenging.5
The dismal therapeutic outcome of AF could be in part due to poor understanding of its underlying mechanisms. A commonly accepted mechanism, the multiple wavelet hypothesis,6,7 characterizes cardiac fibrillation as randomly propagating waves with intermittent blockades, annihilation and re-generation of discrete waves. A consequence of such postulated dynamics is said to be a self sustained complex spatio-temporal patterns of excitation lacking a hierarchical organization in the activation. However, the old idea of Lewis8 on reentry as the mechanism of fibrillation has re-emerged from theoretical9,10 and experimental11 studies showing that wave propagation during AF is not totally random. This postulate is that rotors are the major organizing centers of fibrillation and that certain specific molecular properties of the cardiac muscle at the ion channel level contribute to the establishment of such rotors and to the overall complexity of the arrhythmia.12 A consequence of such complex dynamics paradigm is a hierarchical distribution of local excitation frequencies. Thus, the early studies on regional differences in activation cycle length during AF by Morillo13 and Harada14 were followed by an increased interest in the analysis of the frequency spectrum as a measure of the activation rate in animal models.15-18 Those mapping studies have detected a clear hierarchy in the dominant frequency (DF) of excitation across the atria during fibrillation and correlated the location of the highest frequencies with that of relative organized activity and rotors.15-18 Furthermore, works demonstrating how measurements of AF cycle length in patients can contribute to its treatment,19,20 motivated translation of experimental knowledge and methods into analysis of human AF in the frequency domain. 21
This article has three major objectives: first, to discuss recent experimental and clinical data on wave propagation dynamics and AF maintenance with gradients of DF across the atria; second, to discuss data that strongly support the hypothesis that the ionic mechanism of AF in the structurally and electrophysiologically normal heart may be explained in part on the basis of chamber specific differences in the level of expression of cardiac potassium channels; and third, to discuss the role of structural fibrosis in the dynamics of AF. As will be demonstrated, spectral analysis with the determination of the DF of excitation provides a systematic approach for linking the dynamics of electrical wave propagation in cell cultures, isolated sheep hearts and humans to underlying ionic and structural determinansts of AF.
Frequency-dependent breakdown of wave propagation
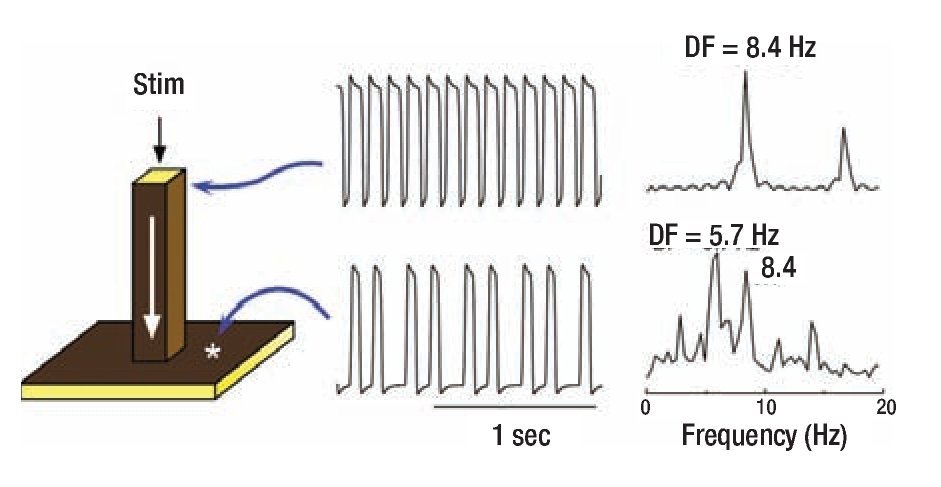
To gain insight into the distinct CLs in Morillo's study,13 we measured the frequencies at varying locations of both atria and found them to be widely different. Most notably, the activation frequencies in certain areas of the left atrium (LA) were always faster than any other region. 15,16 Here we use computer simulation to gain more mechanistic insight into the origin of such data. We start with the premise that the underlying mechanism of AF may be a highly periodic stationary source (e.g., a rotor or an automatic focus) located somewhere in the LA. 17,18 In addition, we assume that, regardless of the nature of the periodic source, the complicated anatomy of the atria plays an integral role in the development of fibrillotory conduction. These assumptions are conveniently considered using a simplified mathematical model of a major pectinate muscle connected to a small area of the atrial wall. 12 The three-dimensional model consisted therefore of a one-dimensional thin bundle attached to a two-dimensional sheet (Figure 1). Periodic stimulation was applied to the top free edge of the thin (25 mm2) bundle and the impulse was allowed to propagate dewnstream to invade the two-dimensional sheet. 12 The traces on the right show the action potentials and corresponding power spectra of sites in the bundle and in the sheet. As shown by the top and bottom time series, stimulation at a constant period of 0.119 sec resulted in a 3:2 propagation pattern across the boundary between the thin bundle and the sheet. This is relected also in the corresponding power spectra: While the source region (i.e., the thin bundle) displayed a DF of 8.4 Hz, the geometrical expansion into the sheet imposed a spectral transformations whereby the DF shifted to 5.7 Hz. The two power spectra display additional peaks originating from the combined effect of the sharp action potential deflections and the inter-beat cycle length variations. The constant cycle length at the thin bundle results in a narrow peak at a DF that is the exact inverse of the cycle length (CL, 1/0.119 = 8.4 Hz) with an additional smaller peak at about 16.8 Hz, which is an integer multiple of the DF (i.e., a harmonic). The CL of the activity in the sheet on the other hand is not constant and can be seen to alternate between short and long values. This in turn gives rise to a more complex profile: Several peaks are seen in the power spectrum, consequent of the combination of various intervals in the time series including not only the long and short CLs, but also their sum and difference. The Fourier algorithm, nevertheless, considers the most stationary combination of the CLs to be the DF at 5.7 Hz, which is the average number of local activations per second,22 corresponding to the 3:2 ratio of the input cycle length (frequency) of 0.119 sec (8.4 Hz) represented by a smaller peak.
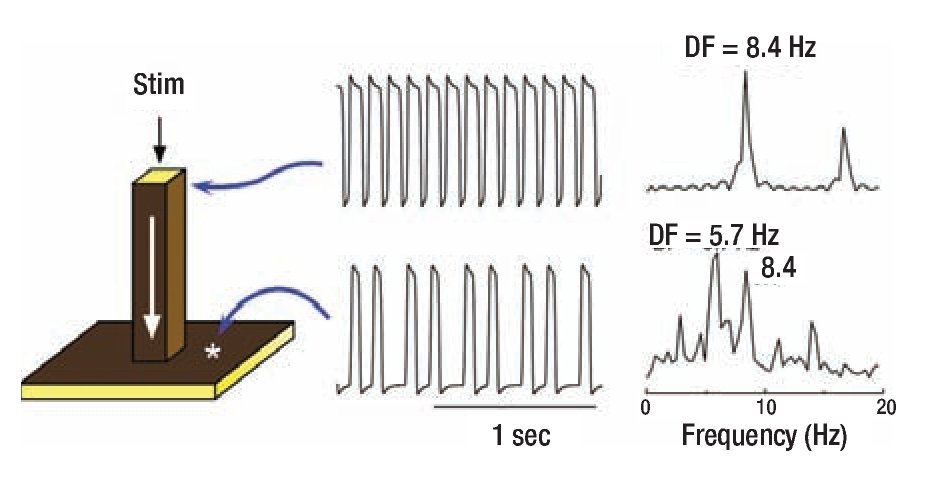
Figure 1. Computer model of action potential propagation from a pactinate muscle to the atrial wall. A 3-dimensional (60x60x60 elements) model includes a 1-dimensional bundle attached to a 2-dimensional sheet (left panel). Periodic stimulation (Stim) was applied at the top edge of the bundle and the impulse was allewed to propagate downward with conduction velocity of ~0.29 m/sec and to invade the two dimensional sheet. The voltage time series and corresponding power spectra are shown for a site near the stimulation point and a site at the sheet. Comparison between the points indicates a 3:2 pattern of propagation into the sheet with a concomitant spectral transformation and a DF shift fron 8.4 to 5.7 Hz. Reproduced from Jalife et al, J Cardiovasc Electrophysiol. 1998;9:S2-S12.12
Keeping in mind all the mathematical and clinical factors discussed above, in the following sections I discuss recent studies that used power spectral analysis of electrical activity during AF. As the reader will be able to tell, in those studies, investigators were able to successfully assign the dominant frequency an electrophysio-logical meaning to determine AF mechanisms and to ghide the AF ablation procedure. It is well known that during atrial flutter both atria are excited in a 1:1 manner at frequencies as high as 300 beats/min (5 Hz). On the other hand, data obtained from experimentally induced AF in the sheep heart do show that stationary reentrant sources in the LA can rotate al frequencies >7 Hz and generate impulses that propagate toward the right atrium (RA). 17,18
Yet the RA in induced AF in sheep experiments is activated at much lower frequencies. This suggests that there must be a critical frequency at which the 1:1 input-output relation between the LA and RA breaks down.
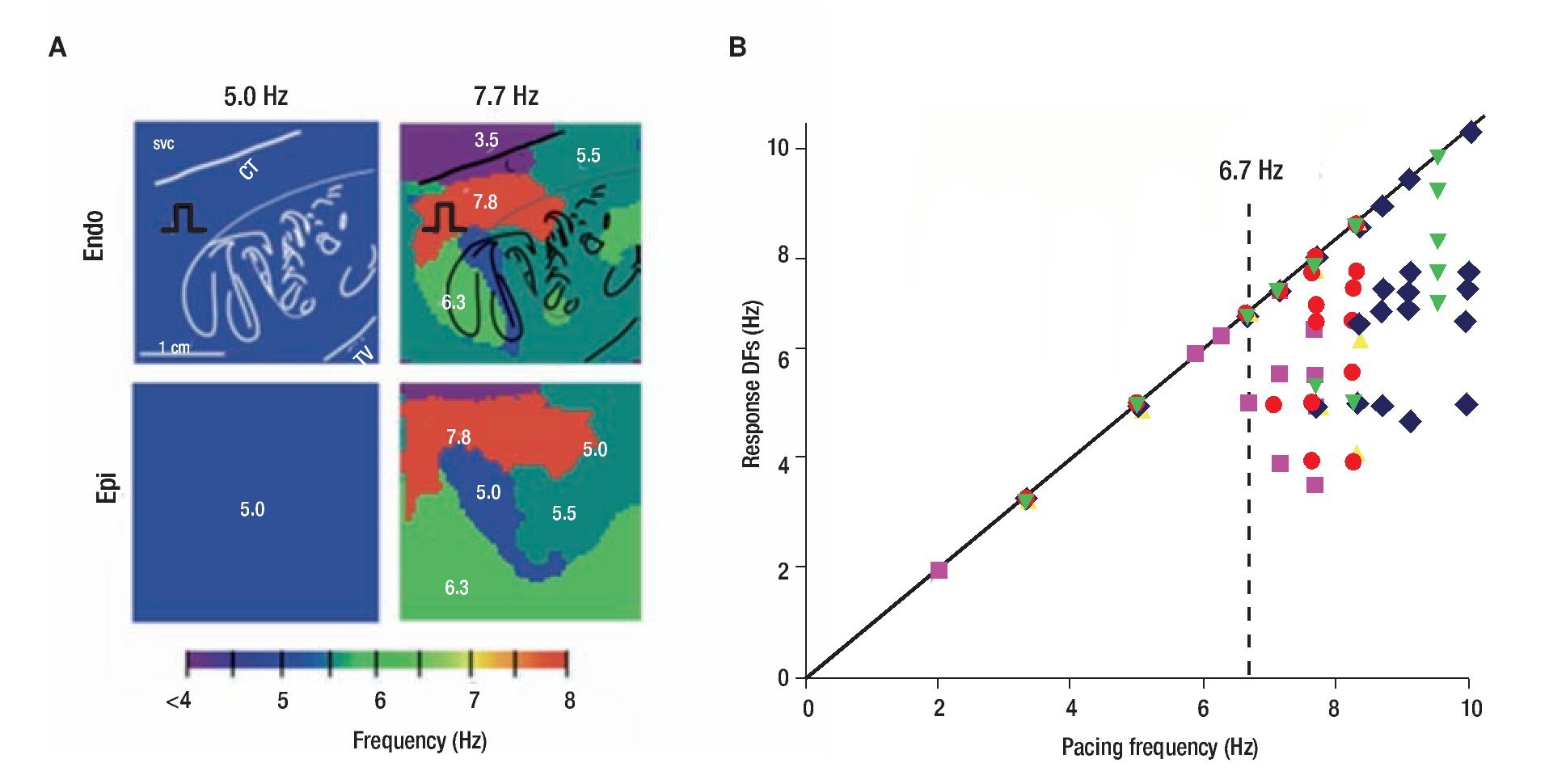
Figure 2 shows results of optical mapping experiments and DF analysis in the isolated RA of a sheep heart adreessing the mechanism underlying of such lack of 1:1 input-output relationship. The preparation was subjected to periodic pacing through a bipolar electrode on Bachmann's Bundle (BB) to simulate activity arriving from a periodic LA source across BB into the RA.23 As shown in panel A, stimulation at 5.0 Hz resulted in 1:1 activation of the entire right atrium and thus the output DF was also 5.0 HZ everywhere on the endocardial and epicardial surfaces. However, at 7.7 Hz, propagation into the RA was no longer 1:1. Instead, a heterogeneous distribution of DF domains was established both in the endocardium and the epicardium, with frequencies ranging between 3.5 and 7.7 Hz. Composite data from five experiments are presented in panel B. The DFs measured on the endocardium, are plotted as a function of the pacing frequency. Clearly, below 6.7 Hz the response DF showed no dispersion in any of the experiments, which meant that activation was 1:1 everywhere in the RA. Above the "breakdown frequency" of 6.7 Hz, there was a large DF dispersion manifested as multiple domains whose individual frequencies were eigher equal or lower than the pacing frequency. In addtion, we found that intermittent blocks occur primarily at branching sites of the pectinate bundles of the atria. Those structurally related blockades lead to a significant loss of consistency in the beat-to-beat direction of wave propagation and provided a direct explanation for the difficulty in tracing back the origin of the activation during fibrillatory conduction.23 We also found that the spatial distribution of action potential duration at a low pacing rate of 3.3 Hz was different from the distribution of DF domains, which led us to suggest that dispersion of refractoriness at normal frequencies is a poor predictor of the spatial distribution of intermittent block patterns that characterize AF outside the source region.23
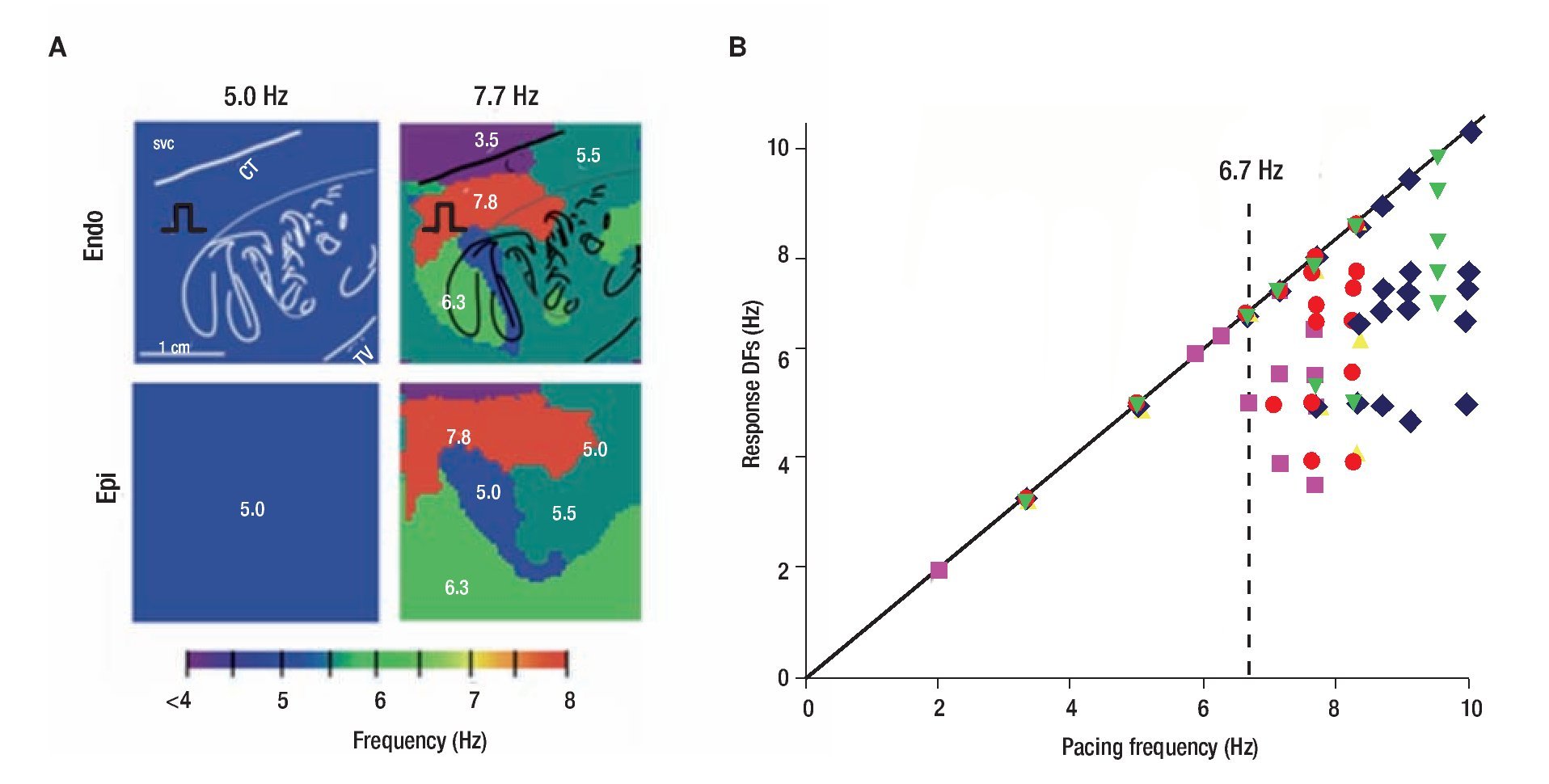
Figure 2. The "breakdown frequency" in a sheep heart. A, Endocardial and epicardial DF maps of same isolated RA preparation paced at 5.0 and 7.7 Hz. Note apeearance of heterogeneous DF domains at 7.7 Hz. B, Response DFs versus the pacing rate (n=5). Each symbol represents one experiment. Pacing BB at rates below ~6.7 Hz, results in 1:1 activation. At higher rates, the number of domains increases but the DFs' value decrease. SVC, superior vena cava; CT, crista terminalis. reproduced from Berenfeld et al, Circ Res 2000;90:1173-1180.23
Reentrant activity during acute AF in the isolated sheep heart
The notion that a localized source of reentrant activity could maintain AF was first postulated by Lewis8 in the early part of the twentieth century and subsequently Scherf.24 Many years later, Morillo et al targeted ablation to sites of short cycle length activity in the posterior LA and observed the termination of arrhythmia in a canine model of AF-. 13 Using a sterile pericarditis model, Kumagai et al identified in the septum dominant unstable reentrant cicuits of very short CL that maintained AF and could be successfully terminated by focal ablation.25,26 Others have reported that in some patients sustained focal activity at the PVs, coronary sins (CS) or superior vena cava initiated and maintained AF, and could be eliminated by discrete ablation.27 However, to this date, whether such sites are either automatic, triggered or reentrant and whether changes in the driver activity would alter spatial frequency gradients, remains unresolved.
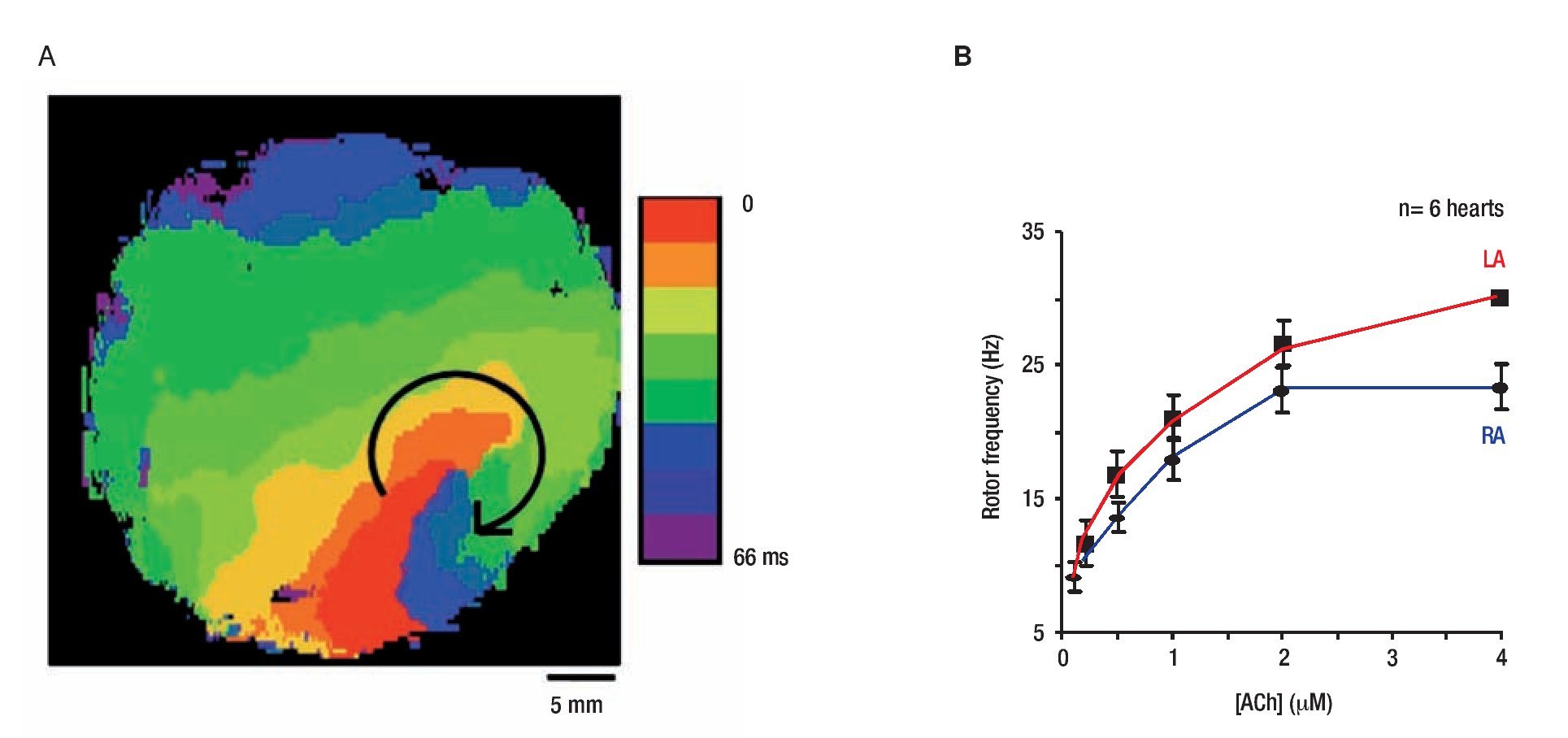
The general working hypothesis that acute AF results from activity of a small number of high frequency reentrant sources localized in one atrium, with fibrillatory conduction to the other atrium is based primarily on results obtained in the isolated, Langendorff-perfused, sheep heart in the presence of acetylcholine (ACh).17 We localized the sources that maintain AF by a combined use of optical mapping and frequency analysis. Figure 3 shows data from on the rotation frequency of rotors during AF episodes in which the site of the high-frequency periodic activity was localized.17 In Figure 3A, an isochrone map from the LA shows one cycle of a vortex that rotated clockwise at a period of 68.6 ± 8.9 ms (~14.7 Hz) and persisted for the entire length of the episode (25 min). The fact that the frequency of this reentrant source (rotor) was equal to the highest and narrowest DF peak (most regular signal) recorded from all other atrial sites (both optical and electrical recordings; see Mandapati et al17 for details) provides direct evidence that it was the mechanism underlying AF in this episode.12
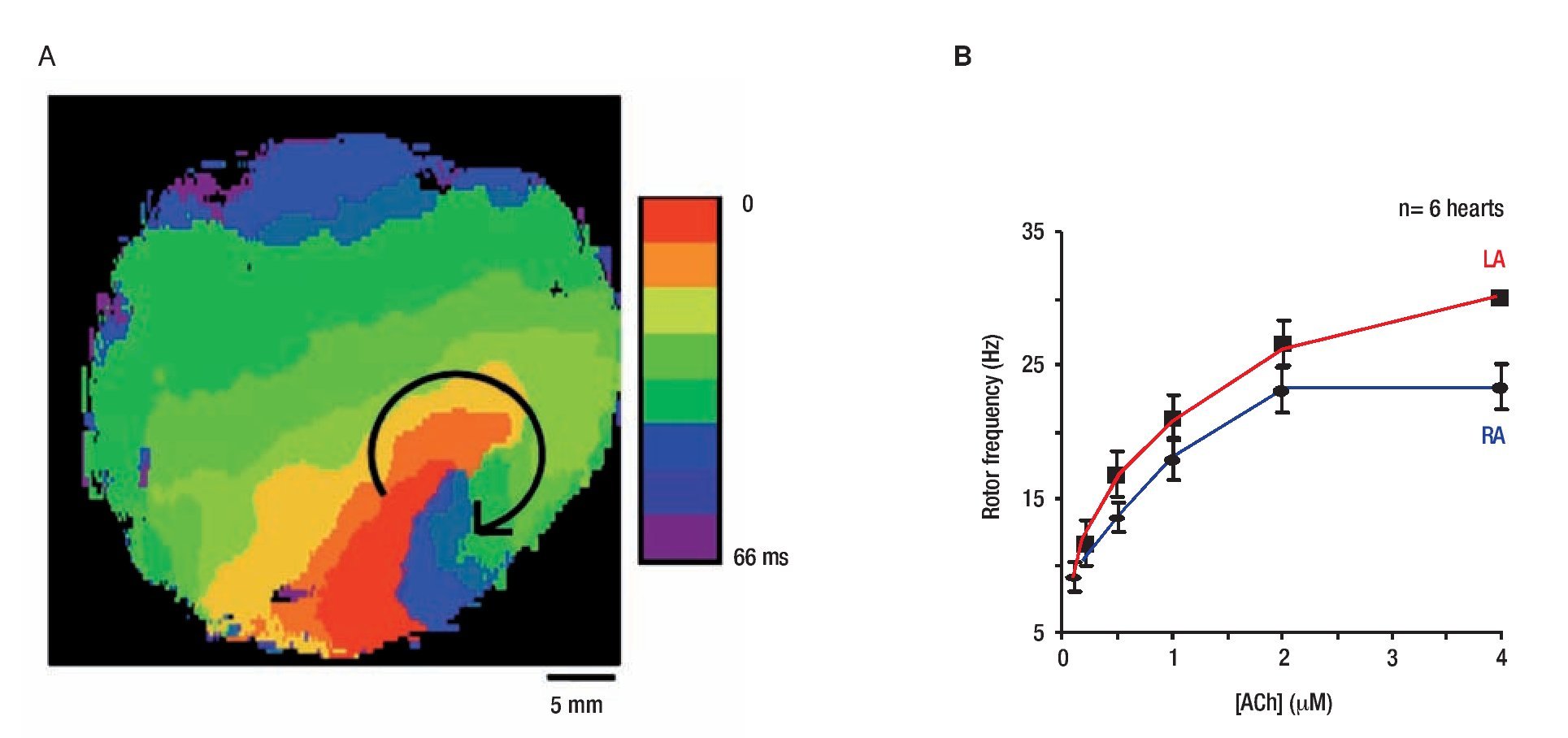
Figure 3. Rotor activity in AF and ACh concentration. A, Isochrone map of optical activity shows a microreentrant AF driver localized to the LA appendage in the form of a rotor rotating clockwise in the presence of ACh. Reproduced from Mandapati et al, Circulation 2000;101:194-199.17 B, RElationship between ACh concentration and rotor frequency created using frequencies from the 5 longest living rotors for each of 6 experiments, at each ACh concentration. There is an increase in rotor frequency with ACh, as well as a LA prodominance over the RA. Reproduced from Sarmast et al, Cardiovasc Res 2003;59:863-873.29
Mapping an isolated canine atrial preparation, Schuessler et al28 found that with increasing concentrations of ACh, activation patterns characterized by multiple reentrant circuits converted to a single, relatively stable, high-frequency re-entrant circuit that resulted in fibrillatory conduction. More recently a systematic evaluation of the effect of ACh on the frequency of rotation in isolated sheep hearts was performed.29 Figure 3B demonstrates the direct ACh concentration-dependence of rotor frequency in both atria of 6 hearts, with frequency being significantly higher in the LA than RA at all ACh concentrations. Moreover, the LA-to-RA rotor frequency gradient also increased with ACh concentration from 1.0 Hz at 0.1μM ACh to 6.2 Hz at 4.0 μM ACh.
An ionic mechanism underlying the DF distribution
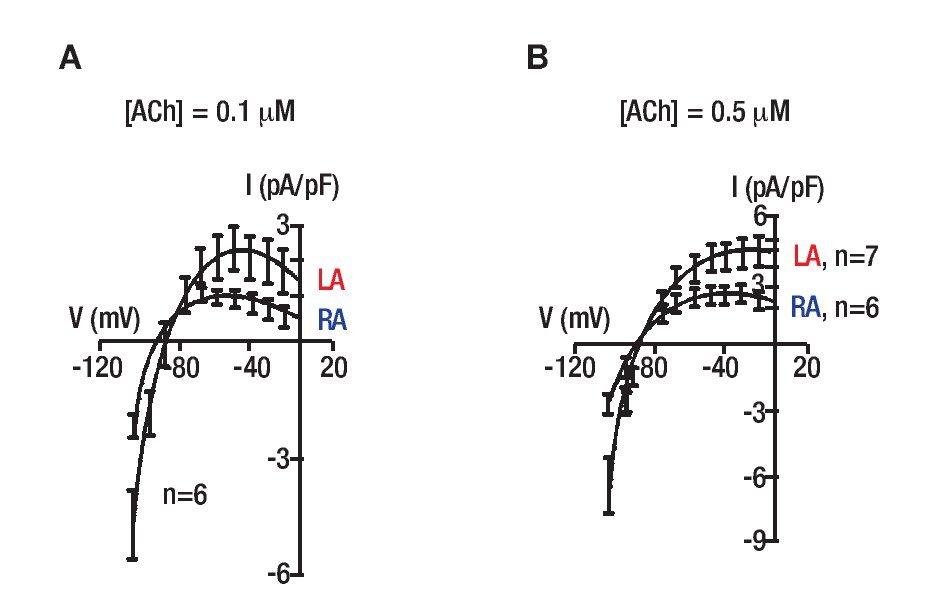
The data presented in Figure 3 suggest that different responses of the rotors frequency in the LA and RA to ACh during AF are somehow related to the ACh-modulated potassium current, IK,ACh. A numerical simulation study using canine atrial cells with realistic ionic and coupling properties, including vagal actions that were formulated based on patch-clamp studies of ACh effects30 VKRZHG WKDW IK,ACh is a determinant of frequency and stability of rotors during AF. Nevertheless, that study did not account for the differential levels of that current in the two atria. To provide a more definite evidence for such contention we studied the effects of ACh (0.05, 0.1 and 0.5 μM) on IK,ACh density of sheep LA and RA myocytes.29 We did no find any significant differences in IK,ACh density at 0.05 +M (shown), however IK,ACh was significantly higher in the LA than in the RA at 0.1 and 0.5 μM. Figure 4 shows ramp-generated current density-voltage relationships in the LA and RA. In Figure 4A, the mean current density-voltage relationship at 0.1 μMACh shows that inward (at-100 mV) and peak outward (at -40 mV) current densities were significantly higher in LA than RA (p<0.05). Figure 4B shows mean IK,ACh-voltage relationships at 0.5 mM ACh. Clearly, the LA has a higher density than the RA also at this concentration of ACh in both inward and outward currentes (p<0.05). Also, note that at the higher ACh concentration,the was an apparent reduction in current rectification. As the strong inward rectifier potassium current, IK1 was found to underlie DF gradients during ventricular fibrillation in the guinea pigs,31 we also investigated its heterogeneity in across the sheep atria. However, additional set of voltage-clamp ramp measurements in the sheep atria found IK1 density to be similar in LA and RA cells.
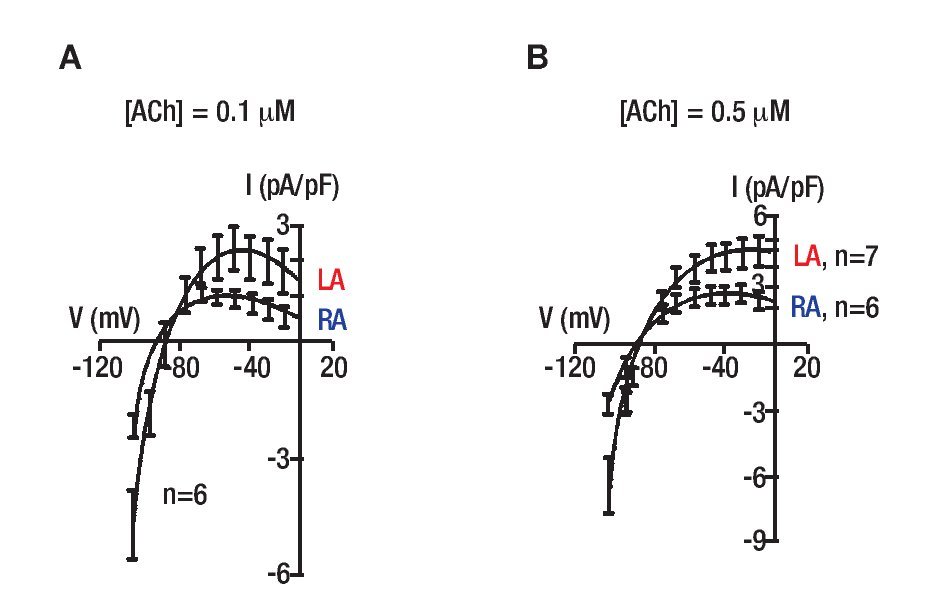
Figure 4. IK,ACh density in sheep atrial cells. A, V-clamp ramp generated currents in the presence of 0.1 µM ACh. Shown are current density in left (LA, n=6) and right (RA, n=6) atrial cells. B, Current density in left (LA, n=7) and right (RA, n=6) atrial cells in the presence of 0.5 µM ACh. LA IK,ACh density is larger than the RA at both concentrations (p<0.05). Note the apparent reduction in current rectification at the higher ACh concentration. Reproduced from Sarmast et al., Cardiovasc Res 2003;59:863-873.29
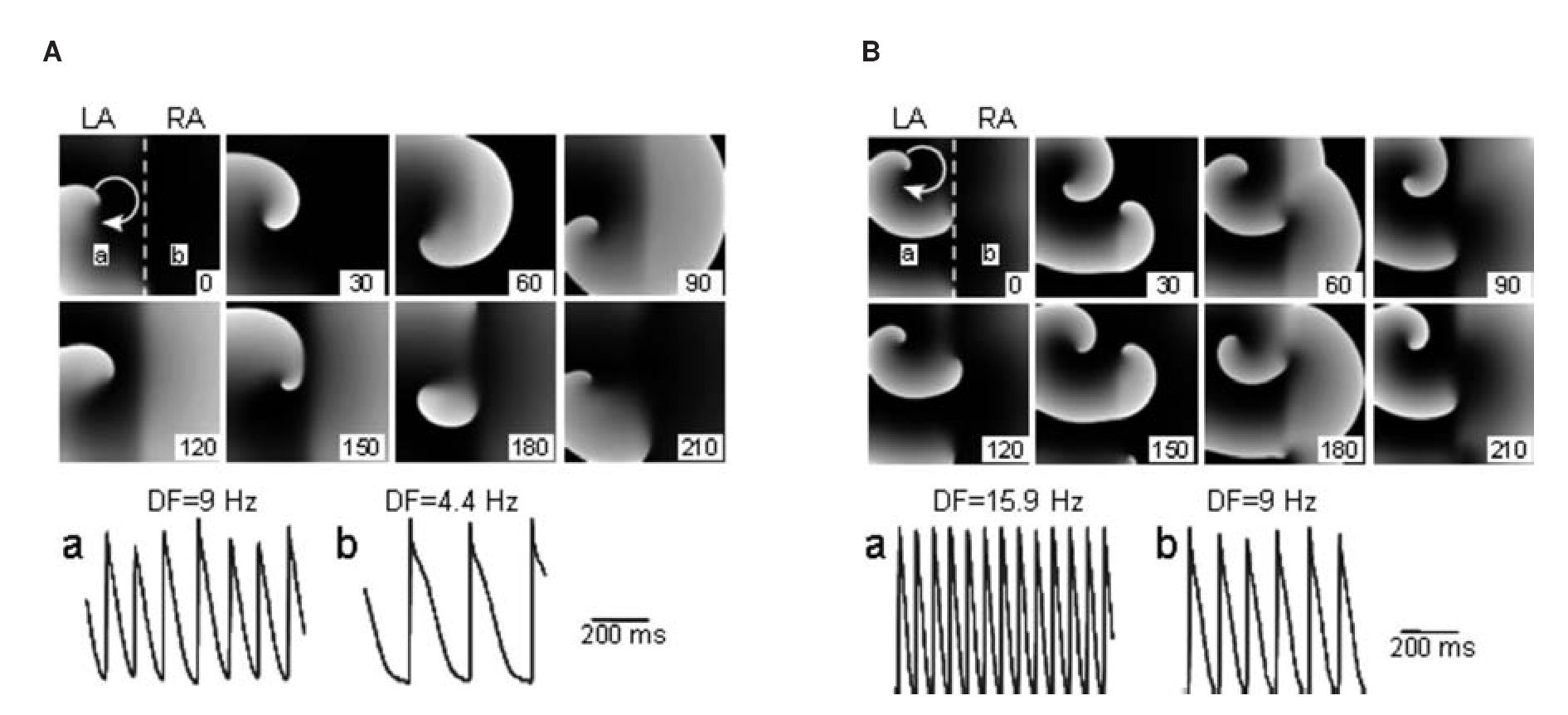
Taken together, the patch-clamp results support the hypothesis that in the sheep heart, LA myocytes can adapt to higher excitation frequencies than RA myocytes. We further studied those findings using a simplified in-silico 2D model of the atria that was used to provide mechanistic insight into the effects of ACh on the maximal DF and LA-to-RA DF gradient by the activation of IK,Ach.32 We used a 5x5 cm2 model with realistic human atrial kinetics33 with heterogeneous IK,ACh density30 and an increased ACh concentration from 0.003 to 0.1 μM, simulating the transition from baseline to increased vagal tone. The model was divided in two whereby one half, representing the LA, was assigned 2 and 1.6 fold the maximal conductance for IK,ACh and IKr of the other half, respectively.29,34 Figure 5 was consecutive snapshots during a simulation of paroxysmal AF at baseline (Figure 5A) and at increased ACh concentration (Figure 5B). In A, stable reentry in the left half of the sheet acted as the high frequency source (a; DF = 9 Hz) of waves that propagated toward the right (b; DF = 4.4 Hz). In Panel B, IK,ACh activation increased the rotation frequency and, consequently, the DFs in the left (15.9 Hz) and right atria (9 Hz). When conditions of persistent AF where established and a reentrant source was simulated in the LA, IK,ACh activation increased the DF in the left (11 to 14.6 Hz) and unified the DFs in the RA from a range of 8.8-11 Hz during baseline to 9.8 Hz during IK,ACh activation. Overall, while the AF frequencies are somewhat different in the simulations than in our sheep experiments and higher than the patients described below,32 qualitatively, the numerical results demonstrate that IK,ACh activation increased the source region DF and consequently the left-to-right DF gradient in both paroxysmal (4.6 to 6.9 Hz) and persistent AF (maximum 2.2 to 4.8 Hz).
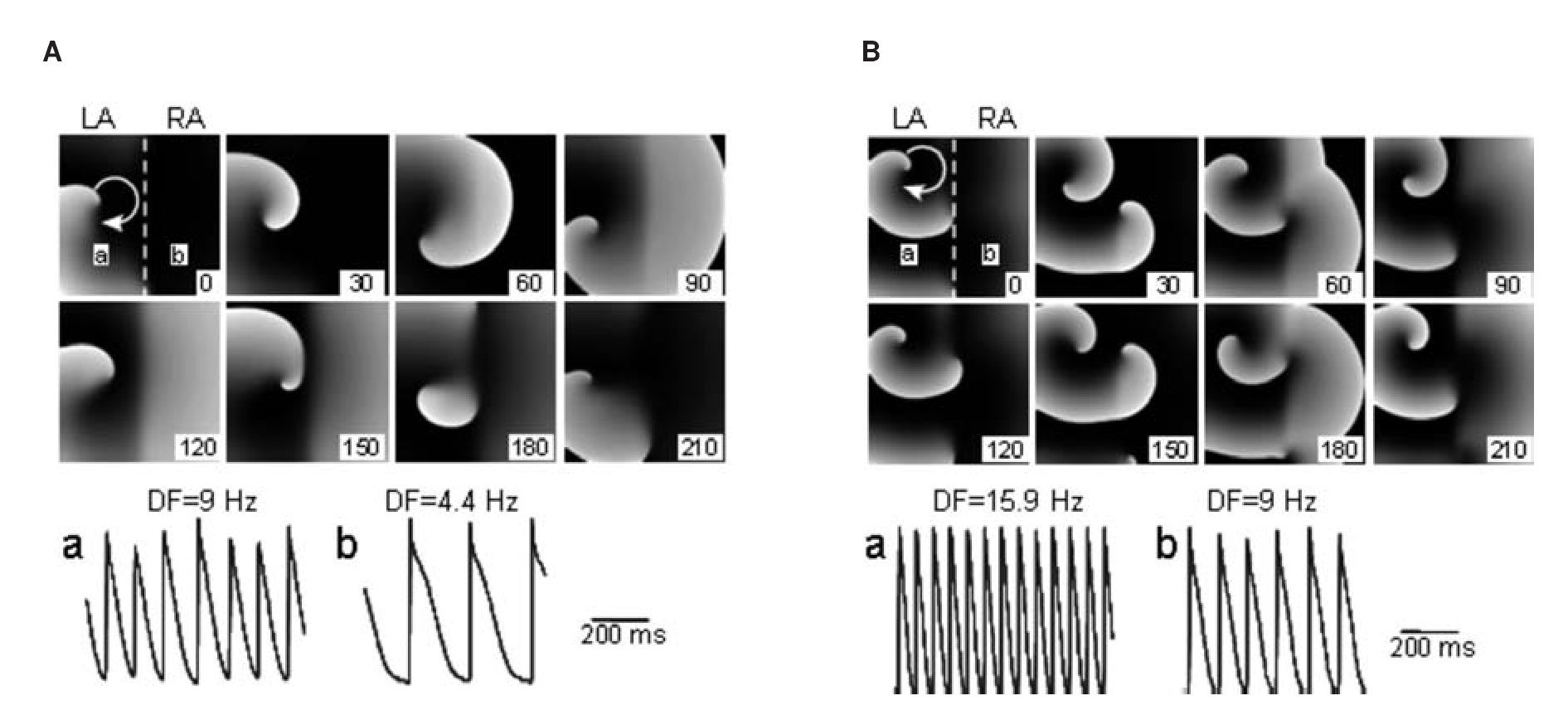
Figure 5. Computer simulations in a 2D sheet of paroxysmal AF at baseline (A) and increased IK,ACh (B). For each snapshot, stable reentry in the left half of the sheet acted as the high-frequency source of fibrillatory waves in the LA (a) that propagated toward the RA (b) in the right part of the sheet. At baseline ([ACh]=0.003 μM) (A), LA DF (a; 9 Hz) is higher than RA DF (b; 4.4 Hz); left-to right gradient is 4.6 Hz. At increased ACh (0.1 μM) (B), DFs in the LA (a) is 15.9 Hz and in RA (b) 9 Hz; the left-to-right gradient is 6.9 Hz. Numbers in frames indicate relative time in milliseconds. Reproduced from Atienza et al, Circulation 2006;114:2434-2442.32
The computer simulations suggest that the activation at extremely fast rates by stationary rotors may be the result of the strong repolarizing influence exerted by their core, which abbreviates the action potential duration (APD) in its proximity.35 This is explained by the fact that during reentry there is a sink-to-source mismatch at the highly curved tip of the pivoting wave creating a central core with excitable but unexcited cells that remain near the resting potential. The slightly depolarized resting potential in the core results in a sustained electrotonic currents between cells in the core and cells in the immediate vicinity of the core. This current in turn drives neighboring cells towards repolarized potentials, and hence, shortens APD. As the inward rectifier current IK,ACh is time-independent it may be able to exert its repolarizing influence and abbreviate APD even at high rates of activity and as such to allow the acceleration of the rotors. However, with increasing distance from the core, this influence weakens and the APD progressively increases.35 Consequently, the tissue close to the core achieves very fast CL's, whereas, far from the core the myocardium cannot conduct at the rate of the rotor and non-uniform (i.e. other than 1:1) conduction develops. In theory, this effect may contribute to the gradient in DFs observed in during fibrillation. As illustrated Figure 3 and in the study by Mandapati et al on AF in isolated sheep hearts17 (as well as in the study by Samie et al on ventricular fibrillation in isolated guinea pig hearts,31) propagation near the rotor is usually 1:1, however, at a certain distance from the rotor, intermittent conduction block and wavebreaks develop and slower DF domains are formed. Nevertheless, while this mechanism may account for the shortening of the APD and the formation of DF domains, it does not explain the consistent localization of the fastest DF domain to the LA which may be further based on dispersion of ionic properties as illustrated in Figure 4.29
Fibrosis in experimental models of CHF sheep and cell cultures
AF is often associated with congestive heart failure (CHF), which induces extracellular matrix remodeling involving atrial fibrosis and dilatation.36-38 In addition to CHF, cardiac fibroblast proliferation and concomitant collagenous matrix accumulation (fibrosis) develop during various insults to the myocardium in ischemic, hypertensive, hyper-trophic, and dilated cardiomyopathies39 and arrhythmogenic right ventricular cardiomyopathy.40,41 Fibrosis per-se has been implicated in arrhythmia initiation and maintenance42 affecting electrical propagation through slow, discontinuous conduction with ''zigzag'' propagation,43,44 reduced regional coupling,45 abrupt changes in fibrotic bundle size,46 and micro-anatomical reentry.47 Most clinical, experimental, and numerical studies have regarded fibroblasts and fibrosis as electrically insulating obstacles,48,49 although some evidence of fibroblast-to-myocyte electrical coupling was shown in the rabbit sino-atrial node.50 However, heart injury promotes fibroblast differentiation into myofibroblasts,51 which have been shown to be coupled electrotonically to myocytes in vitro.52-56 Despite accumulating evidence of potential heterocellular electrical coupling in the diseased myocardium and its implications in arrhythmogenesis, the electrophysiological interplay between myofibroblasts and their neighboring myocytes has not been studied in detail. Myofibroblasts are unexcitable cells; their resting membrane potential is less negative than myocytes, and their membrane resistance is higher.55,57,58 These characteristics suggest that myofibroblasts may function as a sink for electrical charge and as short and long-range conductors.59
Our experimental studies on AF included sheep with atria remodeled by ventricular tachypacing to induce CHF conditions.60 Those experiments demonstrated that posterior LA (PLA) sources maintain AF in both normal and CHF hearts.60 In both groups, AF waves were visualized as breakthroughs traveling from inside to outside the field of view corroborating previous experimental findings61,62 confirming that the PLA plays a key role in maintaining AF activity. Nevertheless the origin sites of AF breakthrough waves in CHF hearts tended to cluster peripherally, in closer vicinity to the PV ostia (PVO) compared with breakthrough waves in control. The shift in the breakthrough sites toward the periphery of the PLA in the CHF vs. control hearts coincided with histological observations demonstrating an increased fibrosis particularly near the PVO in the CHF hearts.60 Spectral analysis revealed a significantly higher DF in the posterior LA compared with other atrial areas in both groups, however significant differences between the groups were also observed: As previously described,61,63,64 regional maximal DF (DFmax) values at other locations (PLA, Bachmann's Bundle and RA appendage) were significantly lower in the CHF group compared with the control group. The reduced local frequency of excitation in our CHF may be related to reduced conduction velocity during AF and pacing, as observed recently in mice pretreated with angiotensin II to increase fibrosis.65 Correlation with anatomical markers of the PLA in the sheep hearts revealed that in control the DFmax domain extended over the left PVO in 2/6 experiments and over both left PVO and center of the PLA in 4/6 experiments. In contrast, in CHF, the DFmax domain localized mainly to the margin of the PLA, enclosing one or several PVO (5/7 experiments), or extended over the entire PLA (2/7 experiments). Interestingly, in all CHF experiments, the signals recorded in the DFmax area were more fractionated than those recorded in other areas. Conversely, the signals from the DFmax area in control hearts were the most regular, in agreement with another report on fractionation in the periphery of DFmax domains during AF.66
Overall, the isolated sheep heart experiments demonstrated that in the PLA major changes in AF dynamics are caused by an increased amount and a different architecture of fibrosis caused by CHF.60 Structural remodeling has been observed in both clinical and experimental AF and is an important feature of the AF substrate producing fibrosis that alters atrial tissue composition and function, however the precise mechanisms underlying AF in the setting of atrial fibrosis are not fully elucidated.67
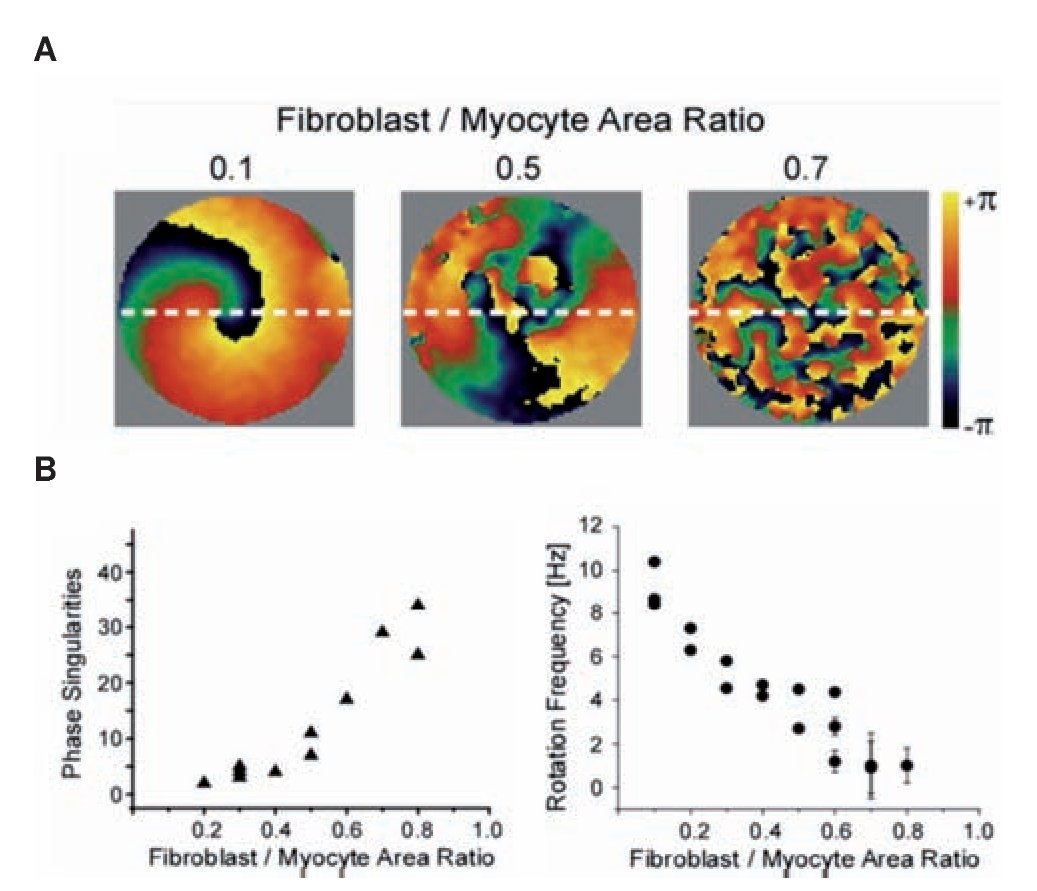
To characterize the effect of fibrosis separately from electrical and other structural remodeling in the myocardium we studied patterns of propagation in monolayers cultured with varying ratios in myocytes and myofibro-blasts.68 The number of myofibroblasts in the co-cultures significantly modified the spatio-temporal characteristics of reentry in the monolayers. Figure 6 shows analysis of propagation patterns and frequency of excitation using optical mapping of such monolayers. Panel A shows representative examples of phase maps during spontaneous activity in the monolayers. Each color represents a phase of the action potential; the convergence of all colors at any given location is a phase singularity (PS).69 Clearly, the number of PSs and wavelets multiplied as the myofibro-blast/myocytes ratio increased from 0.1 to 0.5 and 0.7. In panel B the complexity in the patterns of propagation is quantified by counting the number of PSs and the frequency of reentrant activity; as shown, maximal number of PSs counted in five randomly selected snapshots of a 4-sec-long phase movie (left) and rotation frequency as determined by the DFs across the monolayers (right) are plotted against myofibroblast/myocyte ratio. Each point corresponds to an individual experiment. The graphs demonstrate that as the myofibroblast/myocyte ratio increased, the number of PSs, and thus the complexity of propagation, increased, but reentry frequency decreased. These results are consistent with the results of the optical mapping in the CHF sheep hearts.60 They suggest that fibrosis by itself is an important factor in increasing the complexity of the propagation and the slowing of the excitation frequency, independent of other ionic and structural remodeling that characterize the atria in ventricular tachypaced hearts.70
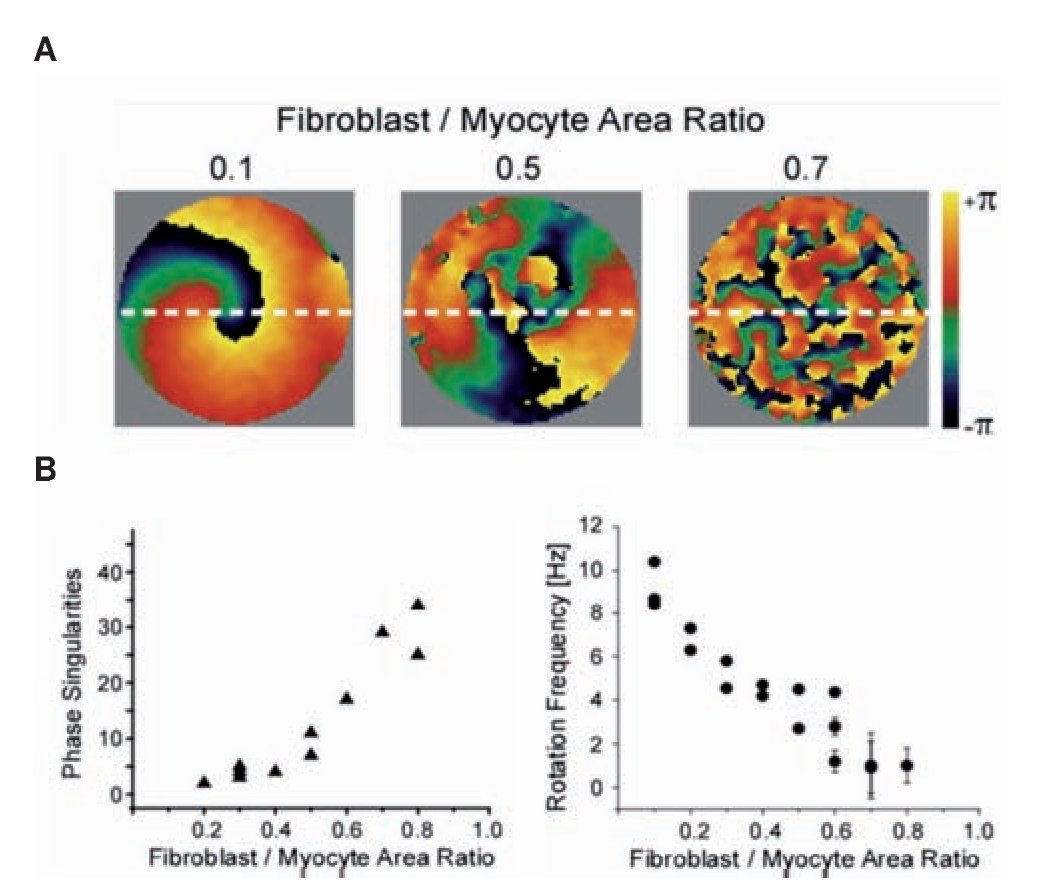
Figure 6. Optical mapping and analysis of complexity and frequency of propagation in hetrocellular mixed monolayers of myocytes and fibrobalsts. A, Phase maps show increasing number of singularity points with increasing number of myofibroblasts in three experiments; myofibroblast/myocyte area ratios: 0.1, 0.5, and 0.7. B, Number of PSs (left, n=14) and rotation frequency and (right, n=19) as function of myofibroblast/myocyte area ratio. Reproduced from Zlochiver et al, Biophys J 2008:95: 4469- 4480.68
The spatial distribution of DFs during AF in patients
Employing the local excitation frequency approach has been demonstrated to be useful in translating mechanistic insights on AF from the experimental to the clinical settings. Several recent studies have characterized the spatial distribution of DF during AF in patients.71-73 Using the CARTO electroanatomic mapping system, Sanders et al73 sequentially acquired 5-sec long bipolar recordings from about 120 points throughout both atria and the CS in 32 patients during sustained AF. The recorded signals were rectified74 and 3-15 Hz band-pass filtered, then fast Fourier transform (FFT) was used to determine the DF of each segment of 4.096 second long with a resolution of 0.24 Hz (Figure 7A).75
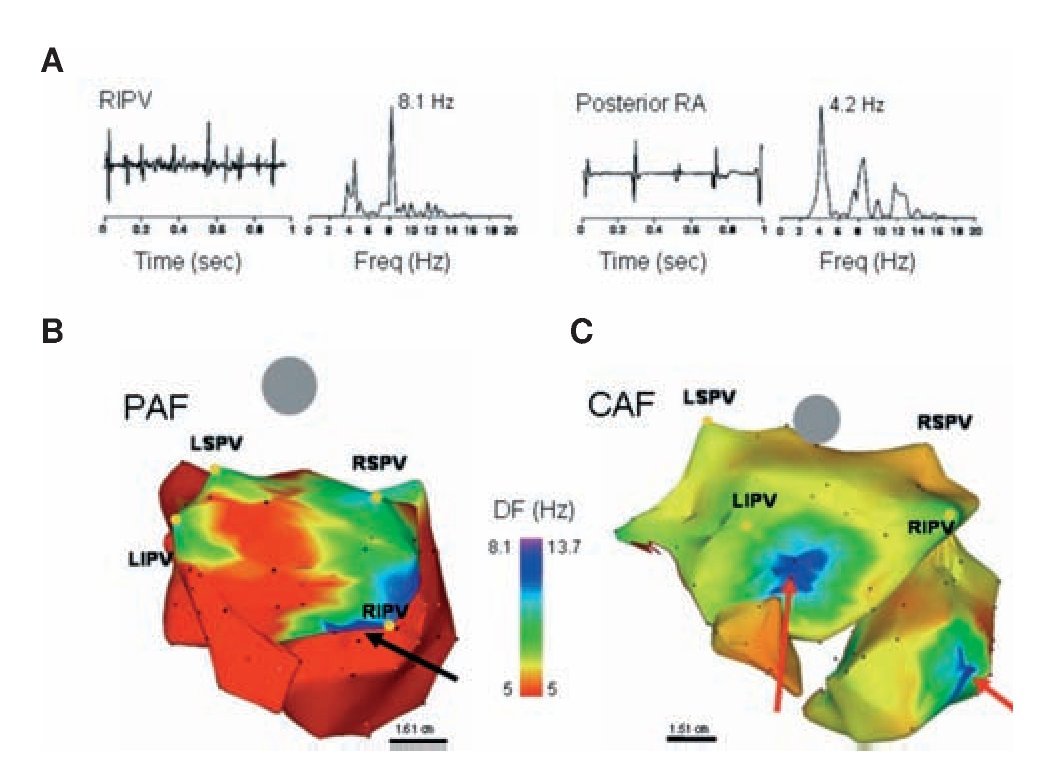
In all patients, electrograms were collected from multiple sites and their corresponding DFs were superimposed on the atrial geometry to generate color DF maps,73 as illustrated in Figure 7. On each DF map, those sites that demonstrated high frequency activity with a gradient of 20% or more relative to the surrounding atrial tissue were defined as high DF (HDF) sites. Figure 7B reproduces a postero-anterior view73 of a DF map in a paroxysmal AF (PAF) patient. While frequency in the majority of the atria and CS was relatively slow (≤5 Hz), the posterior wall of the LA was activated at faster rates (7-8 Hz), with notable HDF sites at each of the PVs. Figure 7C shows a DF map from a patient with permanent AF. Compared with the patient with paroxysmal AF, this patient not only had a higher frequency at the maximal HDF site (13.7 Hz), but also both atria demonstrated higher global frequency of activity and in addition, the HDF sites were located in the atria rather in the PV region.32,73 We concluded that in the cohort of patients studied, paroxysmal AF was characterized by the hierarchical spatial distribution of DFs where the LA and PVs were always the fastest regions. By contrast, in persistent AF, a more uniform distribution of higher DF values was observed, where the highest DFs could not be found in the PV region.22,32,72,73
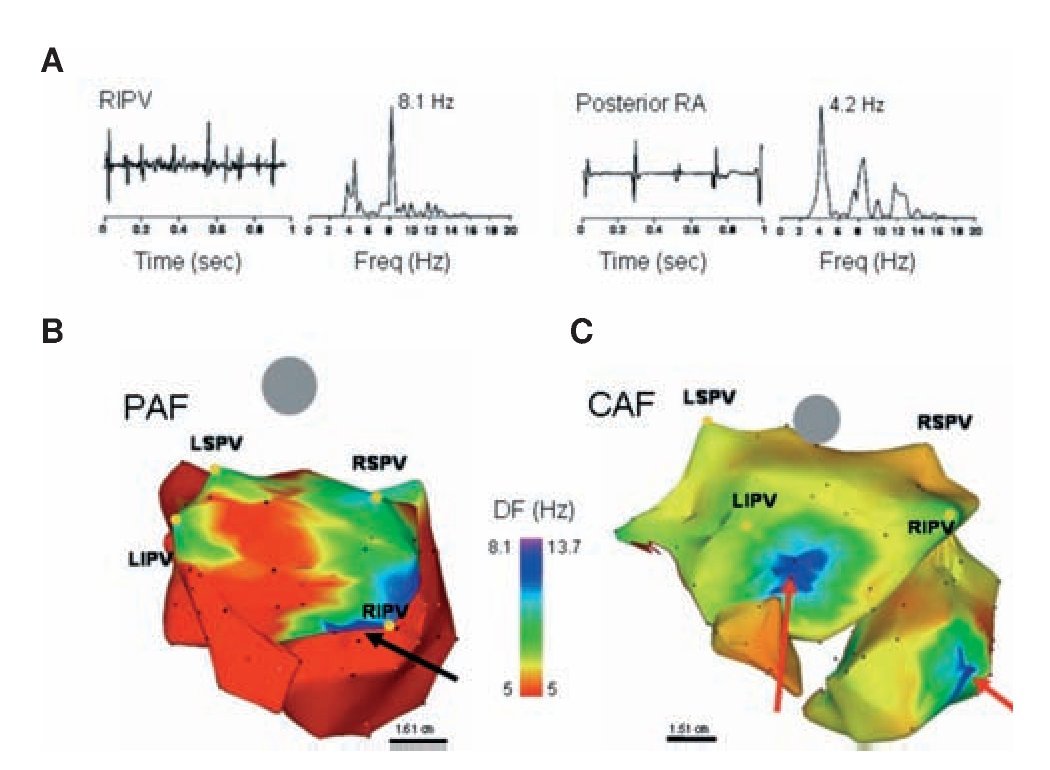
Figure 7. DF analysis in AF patients. A, Bipolar electrograms and corresponding power spectra obtained from the RIPV (left) and posterior RA in a patient with spontaneous paroxysmal AF. Each site shows distinct DF (8.1 and 4.2 Hz in RIPV and RA, respectively) demonstrating the utility of spectral analysis. B, DF map in a patient with paroxysmal AF (Posterior-anterior view; 6 hours). Note HDF sites in each of the PVs. Ablation sequence in this patient was LSPV, LIPV RSPV and RIPV (site of AF termination, black arrow); AFCL increased by 10 ms, 25 ms, 9 ms and 75 ms, respectively, before termination. C, DF map in a patient with permanent AF (24 months). The maximal DF and atrial frequency are higher than the patient in panel A. In addition, HDF sites are located outside the PVs (red arrows). Ablation sequence in this patient was RIPV, RSPV, LSPV and LIPV; AFCL increased by 5 ms, 2 ms, 0 ms and 5 ms respectively. Color-bar, DF scale in Hz. PAF: paroxysmal AF, CAF: permanent AF, LSPV, LIPV, RSPV, RIPV: Left/right superior/inferior pulmonary veins (PVs). Reproduced from Sanders et al, Circulation 2005;112:789-797.73
The high DF sites and maintenance of AF in patients
AF mapping studies have recognized the presence of temporally and spatially periodic activity17,76-78 emanating from the PV region with regularity,15 suggesting that these structures may have a role in maintaining AF,17 by harboring either localized short cycle length reentrant sources and/or focal automatic activity.79,80 Indeed, in the patient whose DF map is presented in Figure 7B, focal radiofrequency ablation applied to the HDF site near the right inferior PV (RIPV, red arrow) effectively terminated AF.73 A recent study utilizing a morphologically accurate computer model of the atria has demonstrated that the PV region is a preferential site for anchoring rotors.81 In the clinic, paroxysms of short cycle length activity have been observed in the PVs of patients undergoing AF ablation.82-84 In addition, sequential ablation of sites showing the shortest cycle length has been associated with a progressive slowing of AF frequency culminating in termination in 75% of patients with paroxysmal AF.20
Using a blind correlation between atrial DF distribution and ablation, without any attempt at identifying potentially arrhythmogenic sites at the time of the procedure, Sanders et al73 found that ablation at PVs harboring HDF sites resulted in an increase in AFCL (≥ 5 ms) within the CS in 89% of cases. The latter was true in patients with either paroxysmal or permanent AF. However, eventual arrhythmia termination occurred during ablation in 15 (88%) out of 17 patients with paroxysmal but none with permanent AF (p<0.0001). In 13 (87%) of the 15 paroxysmal AF patients arrhythmia termination was associated with ablation at a HDF site; 11 localized to a PV and 2 to the LA roof and the fossa ovalis. A more recent study by Atienza et al85 using a real-time mapping of DF allows targeting specifically HDF sites and further demonstrates that abolishing by ablation a preexisting LA-to-RA DF gradients predicted long term freedom of AF in both paroxysmal and persistent AF patients. In persistent AF patients, Yoshida et al86 recently demonstrated that isolation of the PV region eliminated high DF components and transform the fibrillation into tachycardia with a slower frequency. Interestingly, for reasons which are not entirely clear the frequency of the emerging tachycardia waves had a significantly high power during the preceding fibrillation.86 The aforementioned data, together with those of previous studies by Atienza et al32 and Lazar et al,87 clearly indicate that the HDF sites play a role in the maintenance of AF in a significant number of patients.
Activation frequency and driver mechanisms in patients
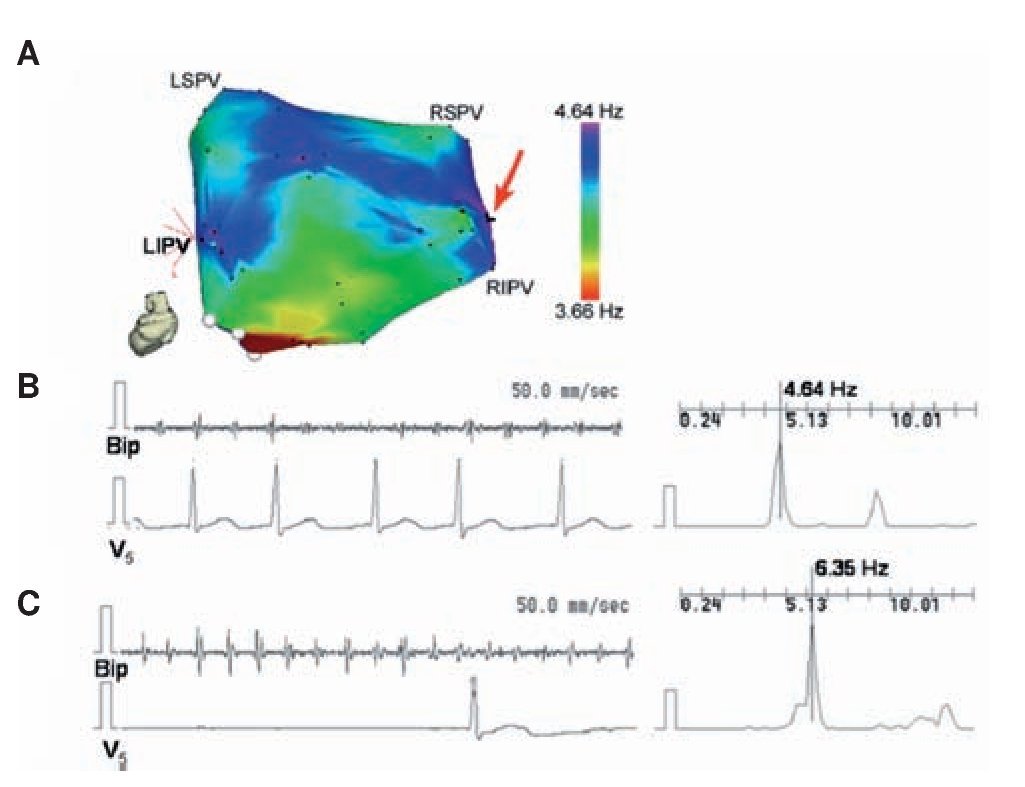
Identifying rotors as drivers of AF directly in patients is very difficult with the existing mapping techniques.88,89 Yet, the ACh dose dependent acceleration of rotor frequency29 shown in Figure 3 offered an idea that enabled translating animal experiments to the patient and gave us the opportunity to obtain evidence, albeit indirect, for the presence of rotors as drivers through pharmacologic means. Translation was made possible also by the fact that adenosine, which is widely used in the clinic, is known to activate the same Kir3.x subfamily of inward rectifier potassium channels as ACh.90-92 By increasing K+ conductance in the atrium, both ACh and adenosine hyperpolarize the cell membrane, abbreviate the action potential duration and the refractory period, and inhibit spontaneous pacemaker discharge as well as early and delayed depolarizations.90,91 On the other hand they both accelerate reentrant activity.29 Thus, in a recent study32 we used adenosine to test the hypothesis that localized reentry maintains AF also in humans. We determined the effects of adenosine infusion on DF at varying locations of both atria with the idea that adenosine-induced acceleration reveals reentry as the mechanism of AF maintenance and rules out an automatic or triggered mechanism. We generated baseline DF maps of the LA using novel real-time spectral analysis software that allowed determination of the specific high DF (HDF) sites likely to harbor the AF drivers73 in AF patients. Then the adenosine effect was measured at the primary and secondary HDF sites in the LA. Figure 8 shows a representative example in PAF where the AF frequency at baseline was relatively slow (<5 Hz) and 3 HDF sites were identified with the primary HDF site being located near the RIPV (red arrow). Panels B and C show that while the adenosine infusion practically abolished the ventricular activity as detected by V5, the DF at the primary HDF site accelerated from 4.64 at baseline to 6.35 Hz at the peak of the adenosine effect. An additional adenosine infusion performed while measuring activity at a secondary HDF site also showed an increase in DF, but to a lesser extent. Interestingly, in this patient the arrhythmia terminated during post-mapping ablation at the primary HDF site, supporting again the critical role of such sites as AF drivers.73 Compared to baseline, adenosine significantly accelerated the primary and secondary HDF sites in PAF patients from about 5 to 6.7 Hz,32 demonstrating that the sites involved in the maintenance of AF are affected by adenosine.
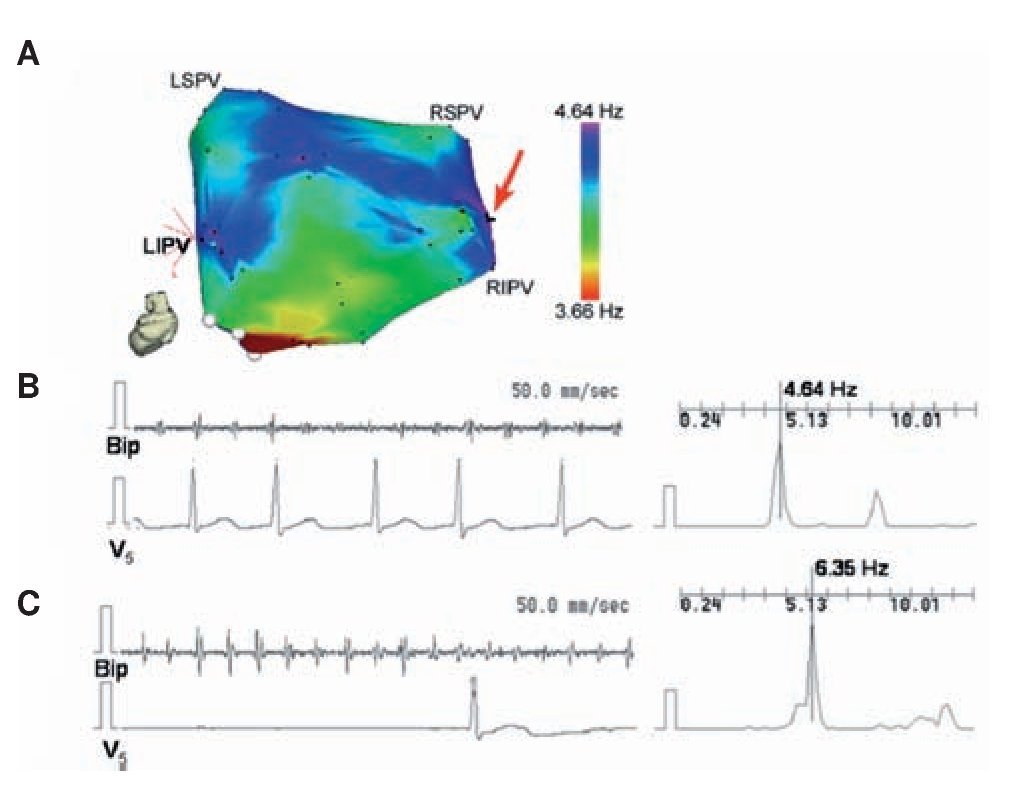
Figure 8. Accelerating effect of adenosine on HDF site. A, LA posterior view DF map from a paroxysmal AF patient. The DF map was produced by the real-time frequency-mapping CARTO system before infusion of adenosine. Red arrow indicates primary HDF site near the RIPV. B, Baseline recording at the primary HDF site with its power spectrum and simultaneous V5 reference. C, Recording at the primary HDF site with power spectrum and simultaneous V5 reference during peak adenosine effect showing increase of DF. LSPV, LIPV, RSPV, RIPV: Left/right superior/inferior pulmonary veins (PVs); Bip, bipolar catheter. Reproduced from Atienza et al, Circulation 2006;114:2434-2442.32
In a larger cohort of paroxysmal and persistent AF patients, Atienza et al32 analyzed the effect of adenosine on the activation rate in specific regions at the junction of the PV and the LA (PV-LAJ), the roof of the RA (HRA) and the CS. In general, patients with persistent AF demonstrated significantly higher maximal baseline DFs than paroxysmal AF patients (p<0.001). However, adenosine infusion in persistent AF patients increased local DFs only in the HRA. The increase in the PV-LAJ was not statistically significant and there was no change in the CS DF. In sum, adenosine infusion increased frequency primarily at sites that activated at the highest rate at baseline. In paroxysmal AF patients, adenosine increased activation frequency in the PV-LAJ. In persistent AF patients, the highest-frequency sources accelerated by adenosine were located in either atria, but not at PV sites. Thus, the DF increase in response to adenosine is consistent with reentrant drivers maintaining AF that have different locations in paroxysmal compared with persistent AF patients.32
AF and fibrosis in post-operative patients
As much as 30% of patients undergoing cardiac surgery develop post-operative AF, which leads to higher morbidity, mortality, and prolonged hospitalization.93,94 While several factors likely contribute to postsurgical AF (e.g., stretch, inflammation, pericarditis, ischemia, autonomic influences), the exact mechanisms of initiation and maintenance are unknown. Structural remodeling of the atria by fatty infiltration, fibrosis, and iron has been implicated in the pathogenesis of chronic AF67 and postsurgical AF.95 Fatty infiltrates and fibrosis form barriers around myocytes creating conditions favorable for reentry.60,67 However, in the literature, biopsies were obtained mostly from the RA of individuals undergoing cardiac surgery95,96 and have demonstrated mixed results regarding the importance of preoperative structural and electrophysiological changes in patients who develop postoperative AF.97
We recently studied 44 patients recovering from coronary artery bypass grafts (CABG) and/or valve replacement/repair surgery.98 Of those, 31 remained in normal sinus rhythm (SR), 12 developed new-onset AF, and one developed new-onset atrial flutter. Of the 44 patients, a subset of 27 was instrumented with epicardial wires for evaluation of LA and RA DFs should they develop AF. Two temporary cardiac pacing wires were sutured onto the LA surface, and the opposing end was brought through the skin. The first recording wire was placed posteriorly at the junction of the LA and LA appendage (LAA). The second wire was placed on the posterior aspect of the LAA approximately 2 cm away from the first one. Two RA leads were placed similarly on the RA appendage and RA free wall, approximately 3 cm apart.98 At the onset, we investigated whether the spatial organization of AF that develops spontaneously after the cardiac surgery is associated with stable LA versus RA differences in DF that persist over a 1-hour period. On average, DF values from 5-second long unipolar recordings (following removal of ventricular activity) were consistent with the DF distribution across the atria in paroxysnmal AF patients.73 LA DFs were significantly greater than RA DFs in all AF patients regardless of the individual LA or RA leads considered (LA: 6.29±0.6 and 6.23±0.6, RA: 4.58±0.7 and 4.55±0.63 Hz. p<0.01 LA vs. RA). The time course of the mean DF values at 10-minute intervals from the LA and RA in five patients who remained in AF for >1 hour shows the LA-to-RA DF difference throughout the 60-minute period despite a slight deceleration in the LA DF.98
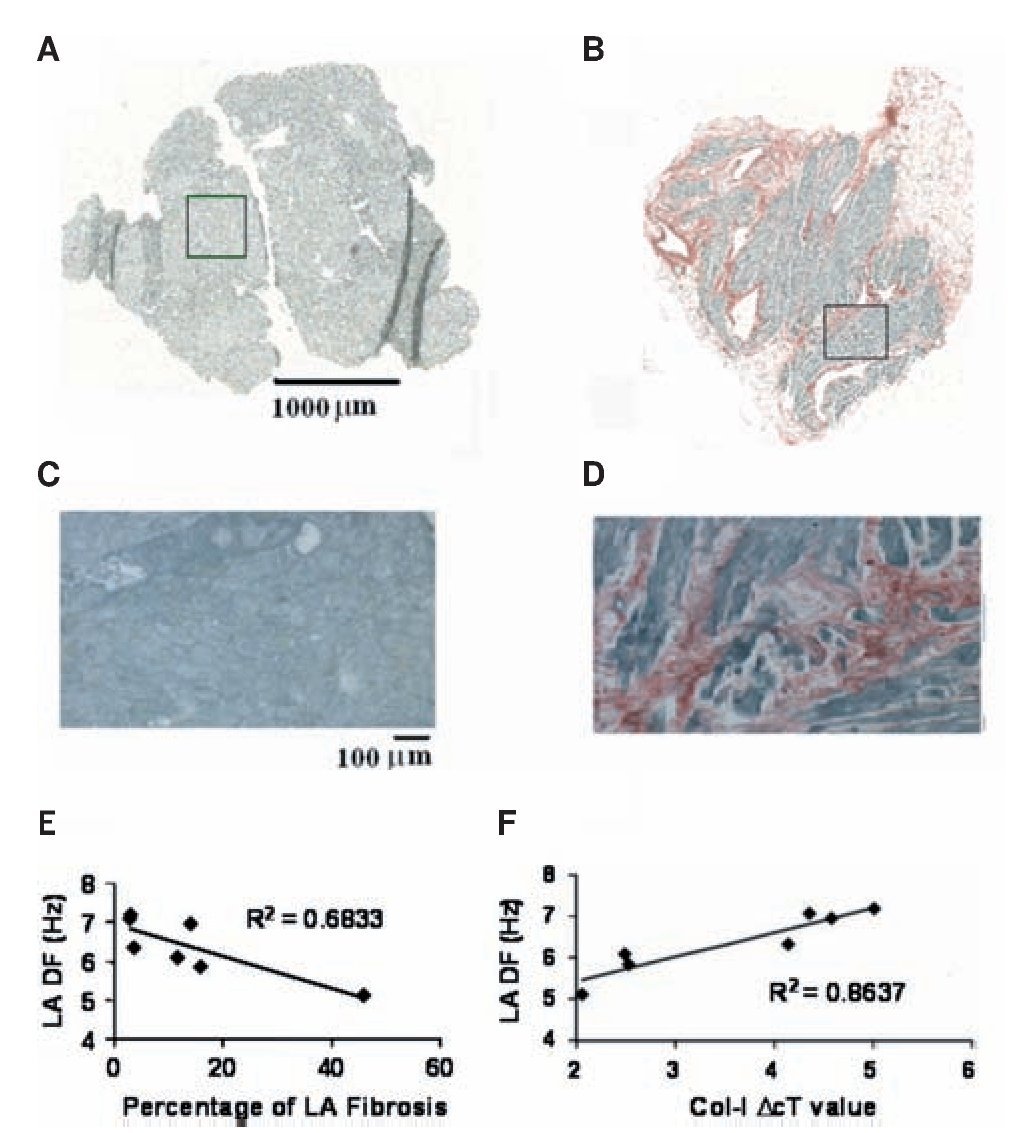
Three- to five-millimeter biopsies were obtained from both LA and RA appendages in all 44 patients during the surgery.98 Panels A-D in Figure 9 show micrographs of 6 mm sections from those specimen stained with picrosirius red. Those examples demonstrate the range of fibrosis seen in our samples. There was no significant difference between the LA and RA biopsies in the area of tissue available for analysis. However, the percent area that was occupied by fibrosis was higher in the LA of patients who developed AF. A statistically significant difference in LA fibrosis was found between those patients who developed AF and those who remained in SR. In contrast, there was no difference in RA fibrosis between these two groups as reported previously.96 Using quantitative reverse transcription-polymerase chain reaction (RT-PCR) analysis we determined LA vs. RA differences in mRNA levels for genes coding for fibrosis.98 We asked whether preoperative differences in atrial structure correlate with the development of postoperative AF. We found that collagen-I (a marker for fibrosis99) expression was similar in patients who developed AF and those who remained in SR. However, in Figure 9E-F, examination of the impact of atrial fibrosis and collagen-I expression on the AF DF in the seven patients who developed AF and had intracardiac recordings demonstrated that an increasing percentage of LA fibrosis resulted in slower LA frequencies. This resulted in an R2 value of 0.68 (P=0.02). However, this did not remain significant after Bonferroni correction. Nevertheless, as Figure 9F demonstrates, when examining collagen-I expression in LA tissue there was a strong correlation between collagen-I expression and LA DF (lower DcT values represent increased collagen-I expression), where the R2 value was 0.86 (p = 0.002), which remained significant after Bonferroni correction.
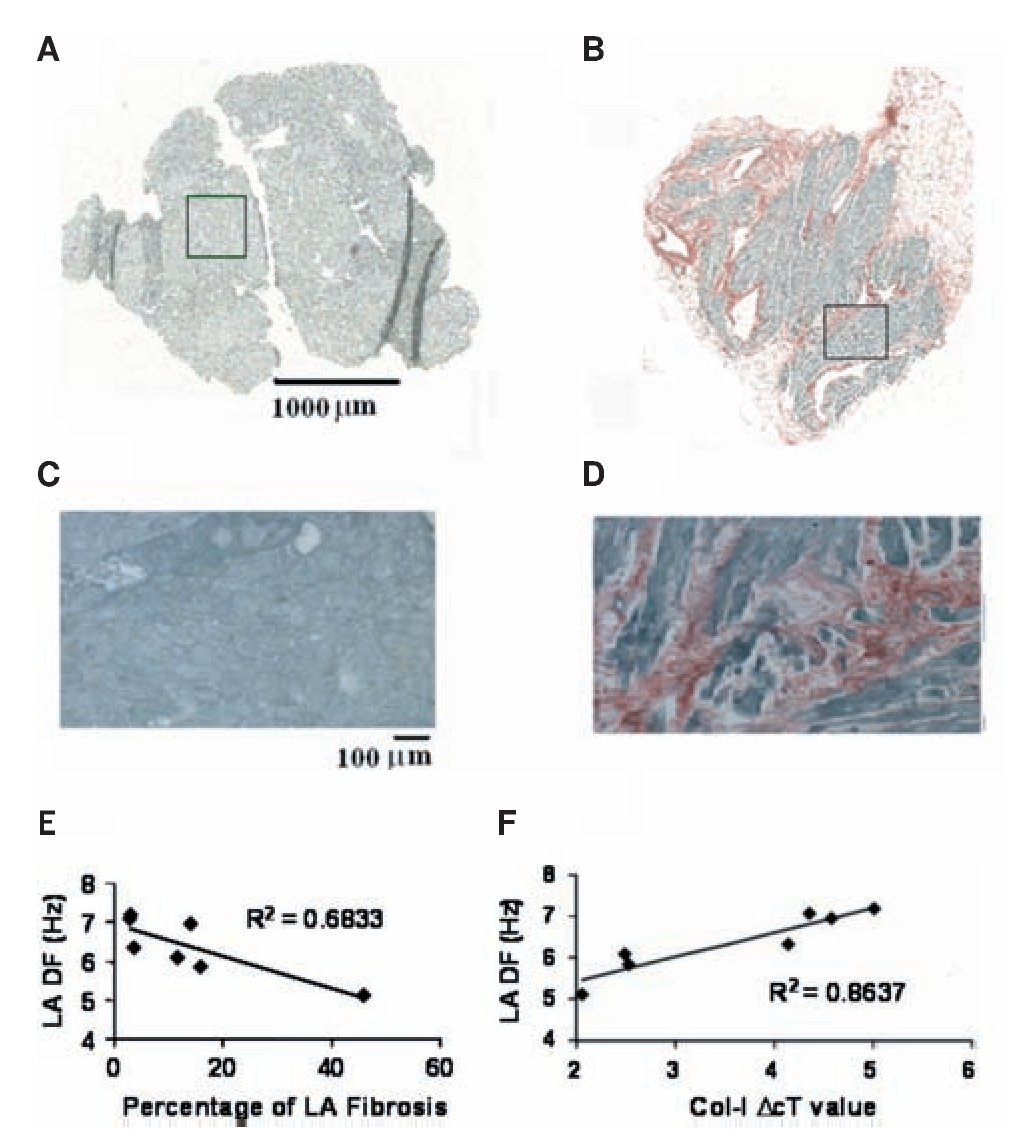
Figure 9. Fibrosis and frequency of excitation during AF in post heart surgery patients. A and B, Montage images of atrial tissue after staining with picrosirius red (fibrosis-red, muscle-green). A, LA biopsy from a 30-year-old patient who remained in SR shows no evidence of fibrosis. B, LA biopsy from a 74-year-old patient who developed AF shows 16% atrial fibrosis. C, Enlarged (x10) boxed region in A demonstrates a lack of fibrosis. D, Enlarged (x10) boxed region in B demonstrates the highest amount of fibrosis. E, Relationship between LA fibrosis and LA DF in seven postoperative AF patients. F, Relationship between LA collagen-I expression (where lower DcT values represent increased collagen-I expression) and LA DF in seven patients who developed postoperative AF. Reproduced from Swartz et al, Heart Rhythm 2009;6:1415-1422.98
In sum, Increasing amounts of LA fibrosis, particularly from collagen-I, are associated with a lower LA DF during AF after cardiac surgery. The accumulation of fibrosis has been found to modify the electrophysiologic properties of the atrium by reducing the velocity of the impulse and providing a substrate for reentry and AF initiation.60,99,100 The finding of reduced DF with increased presence of fibrosis is similar to the reduced frequency of reentry in monolayers with increasing myofibroblasts/myocytes area ratio. Thus, one may speculate that fibrosis in post-operative patients is sufficient to alter the dynamics of an ensuing AF independently of other remodeled factors associated, for example, with age or heart failure.
Closing remarks
The experimental and numerical studies on AF described here were conducted using sheep and simplified computer models. Obviously, results obtained from animal hearts should be cautiously extrapolated to humans. Similarly, one must be extremely cautious when attempting to generalize the applicability of theoretical concepts derived from numerical experiments. Nevertheless, as discussed above, knowledge derived from spectral analysis in the sheep atria and cell cultures may be used to link ionic and structural mechanisms of AF in experiments and patients. In general, the evidence that rotor-like activity is the driving force that maintains both atrial and ventricular fibrillation is compelling.17,29,31,32 and in that regard the analysis of excitation frequency arguably contributed to the recognition that time independent rectifying K+ currents play an important role in the dynamics of rotors and AF.
Indeed, in agreement with our animal and numerical studies on rotor frequency in cholinergic AF in isolated sheep hearts presented in Figures 3-5,17,29 as well as our human studies,32,98 a recent publication describes measurement of IK1 and IK,ACh in cardiomyocytes from RA and LA atrial appendages of SR- and AF-patients undergoing cardiac surgery.101 In that recent study basal inward-rectifier background current was ~2-fold larger in chronic AF vs. SR patients, without RA-LA differences. In PAF, basal current was ~2-fold larger in LA vs. RA, indicating a left-to-right atrial gradient. These results support the hypothesis that a left-to-right gradient in inward-rectifier background current contributes to high-frequency reentrant sources in LA that maintain PAF.101 The findings on K+ currents may allow investigators and physicians to device new antiarrhythmic therapies that are more effective than currently available means in either preventing rotor formation, or terminating rotors after they have had time to stabilize. For instance, our numerical simulations on AF in humans demonstrate that sustained rotors in the atria depend on the conductance level the inward-rectifiers IK1 and IK,ACh.32,102 At normal atrial condition compatible with PAF a presence of IK,ACh is a major player in rotor acceleration and stabilization.29,30,32 However, in persistent AF the increase in IK1 plays a further major role in the dynamics of driving rotors. By altering numerically the rectification profile of IK1 reentrant activity was either slowed down or abolished. Hence, one could envision that, in the foreseeable future, development of a new generation of safe and effective antifibrillatory K+ channels modifying drugs could lead to protection against AF in patients.
An alternative approach for drug-refractory AF in patients is ablation. However this approach can be challenging in certain groups of patients.5 In the future, the combined use of time and frequency domain measures, including electrogram fractionation,103 principal value decomposition and DF mapping,104 should help elucidate mechanism of impulse propagation. The ability to determine whether AF is maintained by discrete sources and to identify their location may help improve the efficacy of ablation procedures. Indeed, DF analysis of bipolar recordings obtained during AF ablation is strongly suggestive of specific high DF sites representing the drivers that maintain AF in certain groups of patients.73,85 Thus, the identification of the drivers and characterization of their dynamics may provide an efficient mean of developing more patient-specific ablation procedures with well defined end-points85,105 potentially leading to increased efficacy and safety.106,107
Acknowledgement
Supported in part by National Heart, Lung and Blood Institute grants PO1-HL39707, PO1 HL087226 and RO1 HL087055 as well as the Gelman Innovation/Innovator Grant Award from the University of Michigan.
Corresponding author: Omer Berenfeld,
Center for Arrhythmia Research. University of Michigan. 5025 Venture Drive. Ann Arbor, MI 48108, USA.
Telephone: (734) 998-7560
E-mail:oberen@umich.edu
Received on August 30, 2010;
accepted on September 30, 2010.