






Cholera continues to be a serious public health issue in developing countries. We analyzed the epidemiological data of cholera from 1976 to 2013 in Shandong Province, an eastern coastal area of China. A total of 250 Vibrio cholerae isolates were selected for PCR analysis of virulence genes and pulsed-field gel electrophoresis (PFGE). The analysis of the virulence genes showed that the positive rates for tcpA and tcpI were the highest among strains from the southwest region, which had the highest incidence rate of cholera. Low positive rates for tcpA, tcpI and ctxAB among isolates from after 2000 may be an influencing factor contributing to the contemporary decline in cholera incidence rates. Spatiotemporal serotype shifts (Ogawa, Inaba, Ogawa, Inaba and O139) generally correlated with the variations in the PFGE patterns (PIV, PIIIc, PIa, PIIIb, PIIIa, PIb, and PII). O1 strains from different years or regions also had similar PFGE patterns, while O139 strains exclusively formed one cluster and differed from all other O1 strains. These data indicate that V. cholerae isolates in Shandong Province have continually undergone spatiotemporal changes. The serotype switching between Ogawa and Inaba originated from indigenous strains, while the emergence of serogroup O139 appeared to be unrelated to endemic V. cholerae O1 strains.
Vibrio cholerae is a Gram-negative bacterium that causes cholera, an acute life-threatening diarrhea disease. To date, over 200 serogroups of V. cholerae have been recognized. However, only the O1 and O139 serogroups are associated with epidemic and pandemic cholera in humans. Based on phenotypic and genotypic differences, the O1 serogroup is further divided into two biotypes, namely, classical and El Tor, and two major serotypes, namely, Ogawa and Inaba. Seven cholera pandemics have occurred globally since 1817, and the first six pandemics are believed to be caused by O1 classical biotype strains.1 It was not until 1961 that O1 El Tor strains became predominant and initiated the seventh cholera pandemic. In 1992, a distinct pathogenic serogroup, O139, was detected in both Bangladesh and India.2 Recently, atypical or variant El Tor biotypes with attributes of both the classical and El Tor biotypes emerged and became predominant globally. The Matlab variants were first isolated in Matlab, Bangladesh between 1991 and 1994.3 Other variant El Tor isolates, including altered El Tor, Mozambique El Tor and hybrid El Tor strains, have been identified in Asia, Africa and America.4–7
During the ongoing seventh cholera pandemic, cholera caused by O1 El Tor arrived in China in 1964 and reached Shandong Province in that same year. After 1964, cholera swept through the entire Shandong Province. Unfortunately, the reports or records from 1966 to 1975 were incomplete due to irregularities in routine work. In 1997, V. cholerae serogroup O139 was isolated from the city of Laiyang for the first time in Shandong Province. Thereafter, serogroup O139 has been the predominant serogroup detected in cholera epidemics in the province. In this study, epidemiological data of cholera in Shandong Province from 1976 to 2013 were collected and analyzed, and on this basis, representative strains of V. cholerae from different areas and years were selected to screen the virulence genes by PCR amplification and study the molecular subtyping using pulsed-field gel electrophoresis (PFGE).
Materials and methodsEpidemiological dataAll records or reports of cholera in Shandong Province since 1976 were collected, and a database including serotype, time, district and epidemic intensity (sporadic or outbreak, meaning a single case or multiple cases, respectively, with an epidemiological link that emerged at the maximum incubation period) was established.
Representative strains and identificationAt least one strain and no more than three strains were chosen from each district with a cholera outbreak. Clinical strains from sporadic cases in various districts were also included. The identification of the isolates was completed. Briefly, lyophilized V. cholerae were enriched in alkaline peptone water for 6h and then streaked on thiosulfate citrate bile salts sucrose (TCBS) agar (Oxoid Ltd, Hampshire, UK) plates and incubated for 24h at 37°C. Typical golden yellow colonies were identified by biochemical reaction and serotyping.
Pulsed-field gel electrophoresis (PFGE)PFGE was performed according to the PulseNet standardized protocol for V. cholerae.8 Genomic DNA of V. cholerae was digested with 50U of the restriction enzyme Not I (New England Biolabs, Ipswich, MA, USA) at 50°C for 4h. Electrophoresis was performed using a CHEF-DRIII system (Bio-Rad, USA). Images were captured on a Gel Doc 2000 system (Bio-Rad, USA). Tiff images were analyzed using BioNumerics v.6.6 software (Applied Maths). The banding similarity was determined by the Dice coefficient with a 1.0% band position tolerance, and a dendrogram was constructed using the unweighted-pair group method with an arithmetic mean algorithm (UPGMA).
Virulence genes detectingChromosomal DNA was extracted from V. cholerae using a DNeasy Blood & Tissue kit (QIAGEN, Germany) as a DNA template for PCR assay. The six virulence genes (ctxAB, tcpA, tcpI, rtxA, hlyA and toxR) were amplified using a S1000 Thermal Cycler (Bio-Rad, USA) as described previously.9–12
Data analysisDifferences of distributions in serotype and virulence genes among V. cholerae were assessed with chi-square analysis using SAS v.10.1 software. Differences were considered significant for p values <0.05.
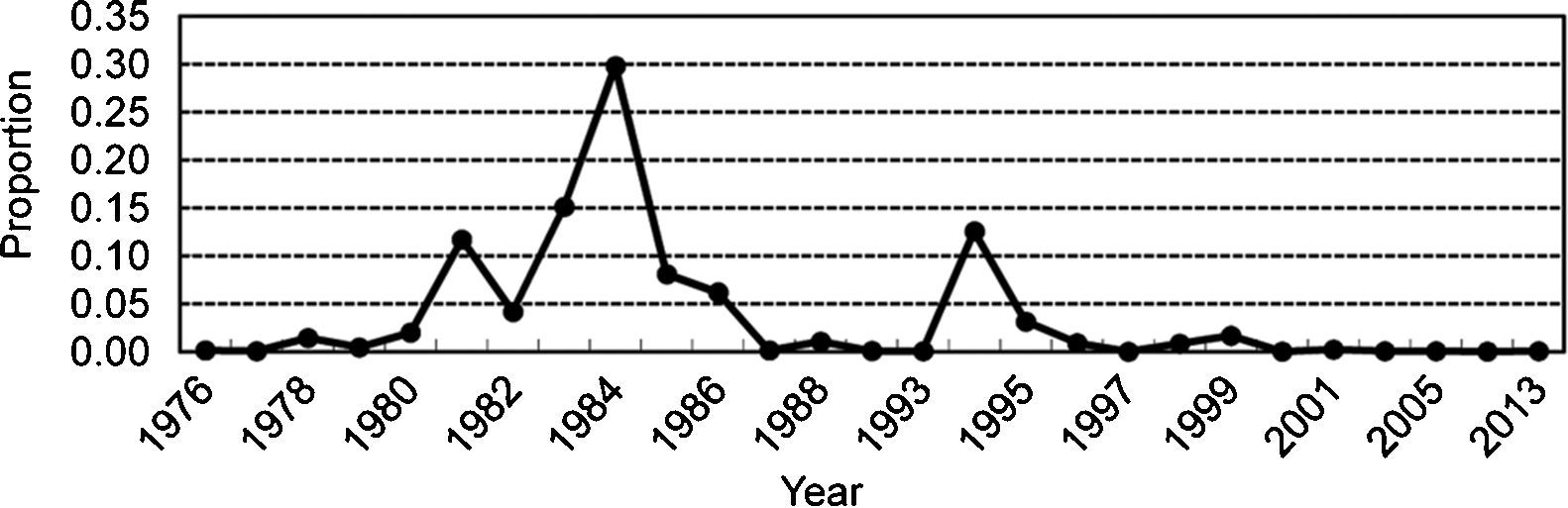
ResultsEpidemiological characteristicsNearly every district of Shandong Province has suffered cholera epidemics since 1976 (Fig. 1). The predominant serotype since 1976 had changed from Ogawa (1976–1979) to Inaba (1980–1989), Ogawa (1993–1999), Inaba (2001), and then O139 (1997–2013). Cities in the southwest (Heze, Zaozhuang, and Jining) and the northeast (Dezhou, Binzhou and Qingdao) of Shandong Province were the hardest hit areas, whereas considerably fewer cholera cases occurred in cities in the central region (Jinan, Zibo, Laiwu and Weifang). Moreover, 58.1% of the patients infected with serotype Ogawa lived in the north, while 85.8% of the patients infected with serotype Inaba lived in the southwest (Table 1).

Distributions of serotypes and virulence genes among V. cholerae isolates from Shandong Province.
| Factor | Percentage of isolates carrying virulence gene | Percentage of serotype | |||||||
|---|---|---|---|---|---|---|---|---|---|
| hlyA | ctxAB | toxR | tcpI | rtxA | tcpA | Ogawa | Inaba | O139 | |
| Source | |||||||||
| Patient | 100 | 76.6a | 100 | 90.4a | 99.1 | 92.2a | |||
| Environment | 100 | 21.9 | 100 | 56.3 | 100 | 56.3 | |||
| Serotype | |||||||||
| Inaba | 100 | 70.4 | 100 | 92.6 | 99.1 | 95.4 | |||
| Ogawa | 100 | 83.2 | 100 | 89.1 | 99 | 89.1 | |||
| O139 | 100 | 77.8 | 100 | 77.8 | 100 | 88.9 | |||
| Temporal distribution | |||||||||
| 1976–1979 | 100 | 55.6a | 100 | 61.1a | 100 | 61.1a | 10.9a | 0.8a | 0a |
| 1980–1989 | 100 | 68.8 | 100 | 92.7 | 99.1 | 93.6 | 51.3 | 96.2 | 0 |
| 1990–1999 | 100 | 94.8 | 100 | 97.4 | 98.7 | 97.4 | 37.5 | 0.8 | 7.1 |
| After 2000 | 100 | 64.3 | 100 | 71.4 | 100 | 92.9 | 0.2 | 2.3 | 92.9 |
| Spatial distribution | |||||||||
| East | 100 | 84.6 | 100 | 82.1a | 100 | 84.6a | 14.8a | 3.4a | 21.4a |
| West | 100 | 73.5 | 100 | 98.5 | 100 | 98.5 | 4.4 | 49.2 | 14.3 |
| South | 100 | 76.7 | 100 | 91.7 | 96.7 | 93.3 | 12.1 | 36.6 | 35.7 |
| North | 100 | 70 | 100 | 85 | 100 | 90 | 58.1 | 4.2 | 0 |
| Central | 100 | 77.4 | 100 | 83.9 | 100 | 87.1 | 10.7 | 6.5 | 28.6 |
| Epidemic patterns | |||||||||
| Sporadic | 100 | 77 | 100 | 86.1a | 99.2 | 88.5a | 39a | 19.4a | 100a |
| Outbreak | 100 | 76 | 100 | 95.8 | 99 | 96.9 | 62 | 80.6 | 0 |
The eastern region of Shandong Province includes Qingdao, Yantai, Weihai and Rizhao; the west includes Liaocheng and Heze; the south includes Jining, Zaozhuang and Linyi; the north includes Dezhou, Binzhou and Dongying; and the central region includes Jinan, Zibo, Laiwu, Taian and Weifang.
Shandong Province has witnessed two peaks in cholera epidemics since 1976 (Fig. 2). One epidemic was caused by serotype Inaba in the southwestern region during 1983–1984, and the other was caused by serotype Ogawa located mainly in the northern districts in 1994. Unlike explosive epidemics caused by serogroup O1 prior to 2001, cholera caused by serogroup O139 during 1997–2013 was usually sporadic. Accordingly, the number of cholera cases decreased significantly in the 21st century.
Representative strains
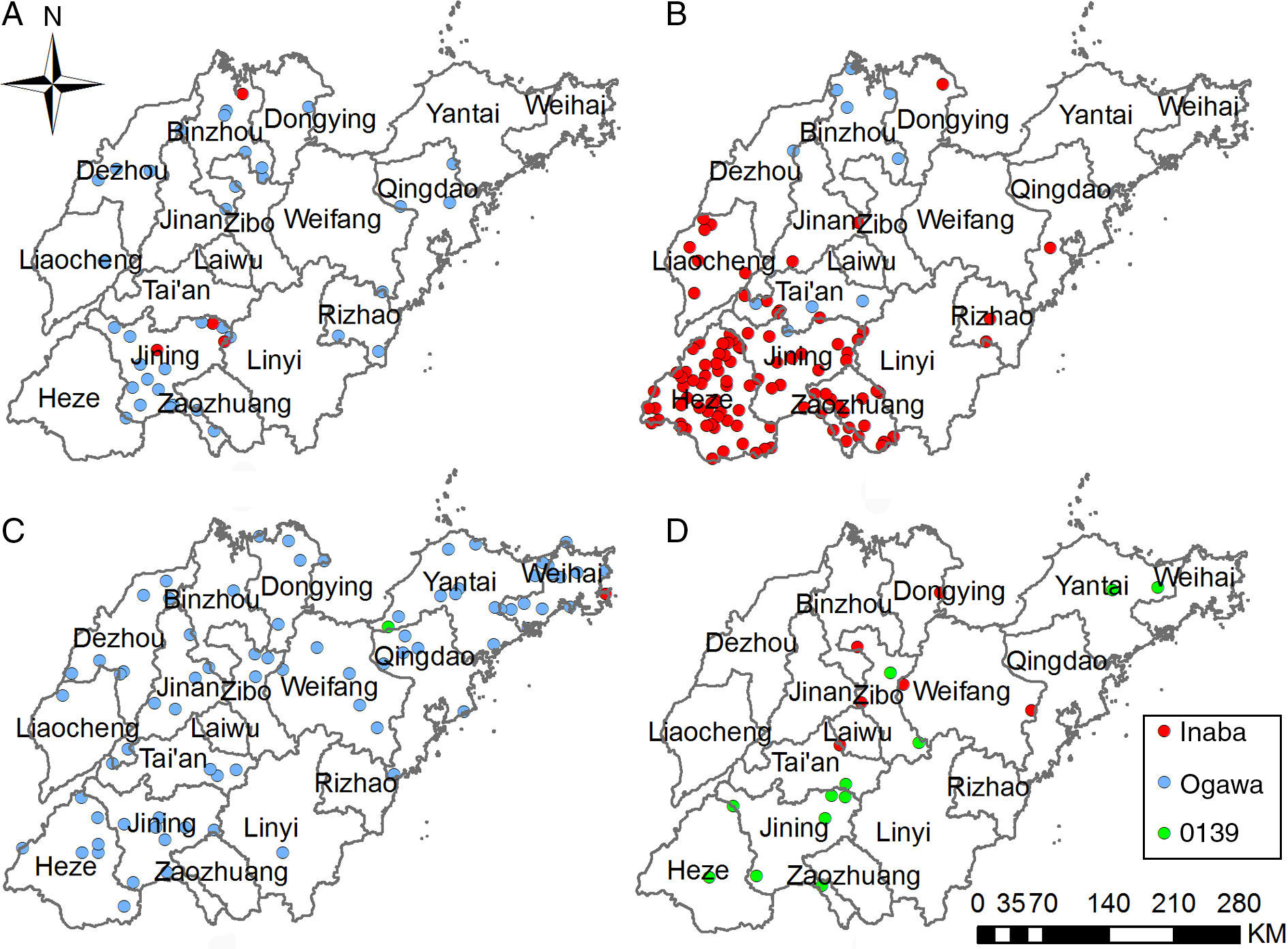
A total of 218 representative clinical strains were selected between 1976 and 2013 from 17 districts in Shandong Province. We also included 32 environmental strains as controls. Among these 250 isolates, 121 strains belonged to serotype Ogawa, 116 strains belonged to serotype Inaba, and 13 strains belonged to serogroup O139. Fig. 3 shows the locations of representative V. cholerae strains with different serotypes from different decades.
 representative strains from 1976 to 1979 were mainly serotype Ogawa and located in the south and north of Shandong Province; (B) representative strains from 1980 to 1989 were mainly serotype Inaba and located in the southwestern regions; (C) representative strains from 1990 to 1999 were mainly serotype Ogawa and spread across the entire province; and (D) representative strains from the 21st century were mainly geographically fragmented O139 strains.")
Locations of representative V. cholerae strains with different serotypes categorized by decades in Shandong Province: (A) representative strains from 1976 to 1979 were mainly serotype Ogawa and located in the south and north of Shandong Province; (B) representative strains from 1980 to 1989 were mainly serotype Inaba and located in the southwestern regions; (C) representative strains from 1990 to 1999 were mainly serotype Ogawa and spread across the entire province; and (D) representative strains from the 21st century were mainly geographically fragmented O139 strains.
All 250 isolates were analyzed using PCR to detect the presence of ctxAB, tcpA, tcpI, toxR, rtxA and hlyA (Table 1). The positive rates of ctxAB, tcpI and tcpA among clinical isolates (76.6%, 90.4% and 92.2%, respectively) were much higher than those among environmental strains (21.9%, 56.3% and 56.3%, respectively). Nearly all the tested strains carried the toxR, rtxA and hlyA virulence genes. Many more of the strains from outbreaks possessed the tcpI and tcpA virulence genes compared to the strains from sporadic cases. However, no significant difference was observed in the distribution of ctxAB between outbreak strains and sporadic isolates. In addition, the positive rates for tcpA and tcpI were highest in the southwestern region. In the clinical strains, the detection rates of ctxAB, tcpA and tcpI increased from 1976 to 2000 and decreased after 2000. Interestingly, the prevalences of the six virulence genes were unrelated to serotype.
Clustering analysis and genetic relatednessAll 250 isolates were differentiated into 179 pulsotypes using PFGE with NotI-digested genomic DNA. UPGMA clustered the pulsotypes into 4 major groups, namely, PI, PII, PIII and PIV (Fig. 4). A general association between serotype and PFGE types was observed, and the serotype switched according to year and region of isolation (Fig. 5).


From 1976 to 1979, the predominant serotype Ogawa was distributed in the northeastern districts. Most clinical strains from northern regions in 1976 and 1977 and eastern parts in 1979 clustered into group PIV, which was also true for 84.3% of the environmental isolates. All the isolates in group PIV were absent of ctxAB and tcpA and showed 67.95% similarity. The clinical isolates from the northern areas in 1978 harbored ctxAB and tcpA and formed subgroup PIIIc.
Cholera epidemics during 1980–1989 were located in the southwestern districts with the predominant serotype Inaba. During the 1980s, 89% of the clinical strains and one environmental strain clustered into subgroup PIa with 69 pulsotypes. Strains collected from the same district in different years or from adjoining districts at the same time typically gave an identical pulsotype (PIa1–PIa12). Additionally, the overall similarity of this cluster was 93.3%.
Serotype Ogawa replaced serotype Inaba and prevailed in the northeast of Shandong Province from 1993 to 1999. All the strains from 1993 to 1997 in the northeastern areas and 2 strains in 1986 clustered into subgroup PIIIb. However, in the following two years, cholera cases were distributed throughout the entire province and all the strains formed subgroup PIIIa.
Serotype Inaba strains reemerged in the central and northern districts in 2001. All the strains from 2001 and one environmental strain from 1994 were grouped together. The O139 strains from 1997 to 2013 formed subgroup PIIa, in which 2 clinical strains collected from Zibo in 2004 shared the same pulsotype with one clinical isolate collected from Jining in 2005 (PIIa1). Additionally, strains isolated from one patient in 2013 and from the cooking utensils used by this patient showed the same PFGE pattern (PIIb1).
DiscussionFrom epidemiological data, 2 different multi-year cholera epidemics were distinguished in Shandong Province since 1976 (Fig. 2), which shared the same pattern with the entire nation. One epidemic was mainly caused by serotype Inaba in the southwestern regions from 1980 to 1989, and the other epidemic was caused by serotype Ogawa mainly in the northern districts from 1993 to 1999. Poor economic level and sanitary condition may have resulted in the 2 multi-year epidemics that corresponded with the characteristic geographic distribution.13V. cholerae exists within an aquatic natural reservoir. Environmental factors, such as climate and weather, play an important role in the evolution of V. cholerae and are probably involved in the different geographical distributions between serotype Ogawa and Inaba in Shandong Province.14
In clinical strains collected prior to 2000, the detection rates of ctxAB, tcpA and tcpI increased over time, suggesting that selective pressures of the environment may exist in Shandong Province.15 Larger numbers of clinical strains carried the ctxAB, tcpI and tcpA genes compared to environmental strains, supporting the fact that cholera toxin (CTX) and toxin coregulated pilus (TCP) are the principal virulence factors of V. cholerae.16 Therefore, the low positive rates for ctxAB (64.3%) and tcpI (71.4%) in the isolates from after the year 2000 would likely be an influencing factor in the decline in cholera incidence rates (Table 1). However, 23.4% of the clinical isolates were devoid of ctxAB, indicating that other pathogenic mechanisms, such as hlyA and RTX genes encoding hemolysin or cytolysin, may play some role in the disease process.17,18 Three genes, toxR, rtxA and hlyA, were carried in nearly all the tested strains, suggesting that they are essential for pathogenicity, especially in strains that lack ctxAB.19
The positive rates for tcpA and tcpI among the outbreak strains were higher than those from the sporadic isolates. Meanwhile, the positive rates for tcpA and tcpI were highest in the southwestern region, which had suffered the most severe epidemics compared to the rest of the province. This distribution pattern proved that the virulence genes tcpA and tcpI were associated with the epidemic intensity of cholera in Shandong Province. Overall, the detection rates for ctxAB, tcpA and tcpI among cholera strains varied in different decades and regions, which relates to the spatiotemporal characteristics of cholera in Shandong Province.
PFGE analysis divided V. cholerae isolates from Shandong Province into four groups based on serotype and expression of virulence genes (Fig. 4). The PFGE grouping pattern changed with time and space, which was in agreement with the serotype transition of V. cholerae. The non-toxigenic isolates, containing the majority of the environmental strains and a few of the clinical strains, exhibited greater diversity than the toxigenic isolates. However, the environmental strain from 1981, possessing ctxAB and tcpA, had the same pulsotype as one clinical strain from 1983, indicating that V. cholerae strains were autochthonous to the aquatic environment. The clinical Ogawa strain from 1993 showed 98% similarity with 2 strains from 1986, revealing their clonal relation. A close genetic relationship (96.8% similarity) was also observed between a serotype Inaba strain that remerged in 2001 and one environmental strain from the 1990s. These results demonstrated that the evolution of indigenous V. cholerae led to a shift between serotypes Inaba and Ogawa.
Serogroup O139 first appeared in Shandong Province in 1997 and has replaced O1 strains in sporadic cases since 2002. The PFGE patterns of the O139 strains differed from the profiles displayed by the O1 strains of clinical and environmental origin, which agreed with the presumption that the progenitor isolate of serogroup O139 in China was introduced from India.20 Further research, such as whole genome sequencing, may help in understanding the origin of the O139 strains from Shandong Province.21 The O139 strains collected in 2013 were separated from those collected in the past ten years with a unique pulsotype predominating in China for the most recent two years, which indicated that the O139 strains from 2013 were imported from other provinces of China.
In summary, the distributions of serotype and virulence genes in V. cholerae isolates collected from Shandong Province, which were associated with molecular PFGE typing, have alternated continually since 1976. This information could be useful for understanding the evolution of V. cholerae and for controlling new potentially epidemic clones.
Conflicts of interestThe authors declare no conflicts of interest.
This work was supported by Grant No. 81202260 of the National Natural Science Foundation of China (http://www.nsfc.gov.cn), and Grant Nos. 2012ZX10004-201-002 and 2012ZX10004215 of the National Science and Technology Major Project of China (http://www.nmp.gov.cn).