



Bacterial endophytes are considered to have a beneficial effect on host plants, improving their growth by different mechanisms. The objective of this study was to investigate the capacity of four endophytic Bacillus strains to solubilize iron phosphate (Fe-P), produce siderophores and indole-acetic acid (IAA) in vitro, and to evaluate their plant growth promotion ability in greenhouse conditions by inoculation into pearl millet cultivated in a P-deficient soils without P fertilization, with Araxá rock phosphate or soluble triple superphosphate. All strains solubilized Fe-P and three of them produced carboxylate-type siderophores and high levels of IAA in the presence of tryptophan. Positive effect of inoculation of some of these strains on shoot and root dry weight and the N P K content of plants cultivated in soil with no P fertilization might result from the synergistic combination of multiple plant growth promoting (PGP) traits. Specifically, while B1923 enhanced shoot and root dry weight and root N P content of plants cultivated with no P added, B2084 and B2088 strains showed positive performance on biomass production and accumulation of N P K in the shoot, indicating that they have higher potential to be microbial biofertilizer candidates for commercial applications in the absence of fertilization.
Most of Brazilian soils, classified as Oxisols,1 show low natural fertility, low pH, and high Al saturation.2 Oxisols have low P availability due to high adsorption of P and the formation of Fe and Al phosphates. The most abundant and least soluble phosphate fraction in pasture, forest, and agricultural Oxisols is iron phosphate (Fe-P), followed by aluminum phosphate (Al-P) and calcium phosphate (Ca-P).3–6
It is well documented that naturally occurring plant growth-promoting bacteria (PGPB) can act in insoluble phosphates form and release a soluble P in the soil solution readily absorbed by plant roots.4,7,8 In addition, PGPB can mitigate abiotic and biotic stresses on plants, including nutrient limitations, heat and drought, exposure to pollutants and antagonistic effect against phytopathogenic microorganisms. The use of PGPB is a favorable strategy in both environmental and economic aspects,7,8 presenting positive results for various crops, including common beans,9 wheat,7,10 maize,11,12 peanuts,12 soybean,13 eggplants, tomatoes, and peppers.14
Most research on PGP microorganisms associated with plants is focused on rhizobacteria; however, there is an increasing interest in the diversity and role of endophytic bacteria for PGP.15 If bacteria are reintroduced in the endophytic form, a more stable relationship can be established between the plant and bacteria, which results in a more efficient promotion of plant growth.16,17
Several rhizospheric or endophytic bacteria belonging to Pseudomonas, Azospirillum, Azotobacter, Burkholderia, Bacillus, Enterobacter, Rhizobium, and Agrobacterium have been reported as PGP microorganisms.18 Among them, Bacillus is frequently reported as a potential biofertilizer due to its multi-functional PGP traits, such as phosphate solubilization,13,14 indole-acetic acid (IAA) production,10,13 siderophore (iron chelator) production,19 and biocontrol ability against plant pathogens.20 In addition, biofertilizers containing Bacillus strains are considered important because of their spore-forming capacity, allowing their adaptation to extreme abiotic conditions, such as extreme temperatures, pH, or pesticide exposure.14Bacillus species have shown to positively affect soybean seed germination by enhancing significantly the root and shoot length, or number of lateral root of the seedling. This effect was related to the production of phytohormone and siderophore and the capability of these bacteria to solubilize phosphate.13 It has also been reported that Bacillus strains significantly promoted seed germination and growth of three test plants, tomato, pepper and eggplant, compared to control plants and the authors attributed this effect to plant-growth promoting traits.14 Other studies also revealed that Bacillus have improved plant growth under drought stress15 and produced a variety of compounds that can be used for the management of a broad range of plant pathogens.16
Previously, in our laboratory, endophytic Bacillus strains isolated from the roots, leaves, and sap of maize showed Ca-P-solubilizing and organic acid-producing ability in vitro.21 Bacterial strains that solubilize P-Ca showed potential as PGPs in soils fertilized with hydroxyapatite rock phosphate (RP) rich in Ca.22 In the present study, four of these Bacillus isolates were selected, representing high (B1920 and B2088) and low (B2084 and B1923) Ca-P-solubilizing ability, to characterize their capacity to produce siderophores and IAA and solubilize Fe-P. Moreover, a greenhouse experiment was conducted to investigate the effects of endophytic Bacillus on pearl millet growth and nutrient uptake.
Materials and methodsScreening of Fe-P solubilizing abilityFour endophytic Bacillus strains, B1920, B1923, B2084, and B2088, previously isolated from maize sap, roots, and leaves,21 were analyzed for Fe-P solubilization in liquid culture. The strains were reactivated on PDA plates (200gL−1 of potato, 20gL−1 of dextrose and 15gL−1 of agar), using the method of streaking for obtaining pure colonies. One colony of each strain was transferred to trypticase soy broth, incubated overnight at 28°C and subsequently 5×107cellsmL−1 of the bacterial suspension was transferred, in triplicate, to 100mL of National Botanical Research Institute's phosphate growth (NBRIP) medium22 supplemented with FePO4 (5gL–1) and incubated at 28°C for 9 days at 120rpm. After incubation, the cultures were centrifuged at 5000×g for 10min, the supernatant was filtered using Whatman no. 42 paper, the soluble P concentration was determined,23 and the pH was measured.
Siderophore productionThe isolates were assayed for siderophore production using chrome azurol sulphonate (CAS) agar plate assay,24 modified by Pérez-Miranda et al.25 Briefly, 10μL of an overnight culture (aprox. 107cellmL−1) of each isolate grown in a LB medium was transferred to an agar nutrient medium and incubated at 37°C for 16h. After, the siderophore production was detected using an overlay technique, in which a modified CAS medium was applied over the plates containing the cultivated microorganism and incubated in the dark at 25°C for four days. A change in color in the overlaid medium indicates the type of siderophore produced, i.e., from blue to purple for siderophores of the catechol type, to orange for siderophores of the hydroxamate type, or to light yellow for siderophores of the carboxylate type.
Indole-acetic acid (IAA) productionTryptophan-dependent IAA production was measured using the colorimetric method by Patten and Glick.26 Briefly, single bacterial colonies of each endophytic strain were inoculated into TSB medium supplemented with 1.0mgmL−1 of l-tryptophan as an IAA precursor and incubated at 30°C for 5 days at 100rpm in the dark. After centrifugation at 5500×g for 10min, a 0.1mL aliquot of the supernatant was mixed with 0.1mL of Salkowsky's reagent27 and incubated at room temperature for 20min in the dark. The level of IAA present in the culture supernatant was determined in triplicate by colorimetric measurement at 540nm and compared with a standard curve.
Greenhouse experimentA pot experiment was carried out in a greenhouse to evaluate the effects of the Bacillus strains on shoot and root biomass (dry weight) and N, P, and K mobilization using pearl millet BRS1501 (Pennisetum glaucum) as the test crop. Pearl millet was chosen because presents fast development with short life cycle responding particularly well to nutrient deficiency with high capacity of absorption and cycling of nutrients. The experiment consisted of a factorial of three P treatments and four bacteria strains, as well as an uninoculated control treatment consisting of seed coated with sterilized mineral coal and cassava starch gum without bacteria, arranged in a completely randomized design with four replicates. The experiment was performed using 4kg of red clay-texture oxisol, pH 5.2 (soil to water ratio of 1:2.5 (w/v)), containing 0.4cmolcdm−3 Al, 2.5cmolcdm−3 Ca, 0.2cmolcdm−3 Mg, and 30mgdm−3 K, cation exchange capacity of 11.8cmolcdm−3, base saturation of 23.2%, and clay content of 74dagkg−1. The original content of available P in the soil was 2.2mgdm−3.
Approximately 10 days before planting, fertilization was carried out with a nutrient solution without P28 (285.8mgdm−3 NH4NO3, 382.4mgdm−3 KCl, 123.9mgdm−3 (NH4)2SO4, 2.9mgdm−3 H3BO3, 7.9mgdm−3 CuSO4·5H2O, 9.2mgdm−3 MnSO4·H2O, 8.3mg dm−3 ZnCl2, 0.5mgdm−3 (NH4)6Mo7O24·4H2O), which was replaced by a half-strength of the same nutrient solution 20 days after sowing. The P sources were triple superphosphate (TSP) and Araxá rock phosphate (RP), both at 300mgPdm−3 soil and a treatment with no P added. Twenty seeds of pearl millet, coated with appropriate bacterial inocula, were sown in each pot, and the seedlings were later thinned to eight plants per pot. The bacterial inocula were prepared as follows: cells from 50mL cultures incubated for 96h in LB medium were harvested by centrifugation at 10,000×g for 10min, resuspended in a 0.85% (w/v) NaCl solution and the optical densities were adjusted to 1.0 absorbance at 540nm corresponding to 108cellsmL−1. Subsequently, the suspensions were added to the sterilized mineral coal (inoculum carrier) in the proportion of 109cellsg–1 of mineral coal. The inoculant (bacteria+mineral coal) was pelletized onto millet seeds with 4% (w/w) cassava starch gum at a final concentration of 108cellsseed–1.
Harvesting was carried out 50 days after planting, and roots and shoots were separated and dried in a forced air circulation oven at the temperature of 65°C until constant weight in order to obtain dry matter. Then, the plant material was ground in a Wiley mill, and chemical analyses were conducted for determining the N P K concentration in pearl millet shoots and roots in the Laboratory of Plant Chemical Analysis at Embrapa Milho e Sorgo using ICP-OES.29 The N P K content was calculated by multiplying the N P K concentration with the dry weight, which was performed separately for roots and shoots.
Statistical analysesData were subjected to variance analysis using the SISVAR software, version 5.6,30 and the biological means were compared by the least significant difference (LSD) test at 5% probability.
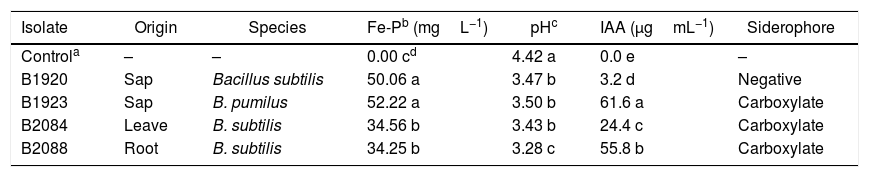
ResultsPGP potential in vitroA high variability of Fe-P solubilization among the strains was observed after incubation at 28°C (Table 1). In the absence of bacteria (control), the concentration of soluble P in the growth medium was very low, and no significant decrease was observed in pH after 9 days. Solubilized Fe-P measurements ranged from 34.25 to 52.22mgL–1, and the highest solubilization was achieved by the B1923 and B1920 strains, while the lowest was achieved by the B2084 and B2088 strains. All strains that solubilized Fe-P reduced the pH of the media compared to the control treatment, with a significant and negative correlation between the amount of soluble P and the final pH of the media (r=−0.84; p<0.05). In addition to their phosphate-solubilizing potential, all strains, except B1920, produced the carboxylate type of siderophores when grown on CAS medium agar plates (Table 1).
Isolate, local of origin, species, Fe-P solubilized, pH, IAA and siderophore production by four endophytic Bacillus strains after 9 days of growing at 28°C.
| Isolate | Origin | Species | Fe-Pb (mgL−1) | pHc | IAA (μgmL−1) | Siderophore |
|---|---|---|---|---|---|---|
| Controla | – | – | 0.00 cd | 4.42 a | 0.0 e | – |
| B1920 | Sap | Bacillus subtilis | 50.06 a | 3.47 b | 3.2 d | Negative |
| B1923 | Sap | B. pumilus | 52.22 a | 3.50 b | 61.6 a | Carboxylate |
| B2084 | Leave | B. subtilis | 34.56 b | 3.43 b | 24.4 c | Carboxylate |
| B2088 | Root | B. subtilis | 34.25 b | 3.28 c | 55.8 b | Carboxylate |
The bacterial strains were able to produce IAA in the presence of l-tryptophan with a significant difference, and the yield ranged from 3.2 (B1920) to 61.6mgL–1 (B1923) (Table 1).
PGP potential in the greenhousePlants fertilized with soluble phosphate significantly produced the highest shoot and root biomass and N P K content. In general, no significant differences were observed between the biomass and nutrient content of plants cultivated with no P added or RP fertilization (Tables 2 and 3).
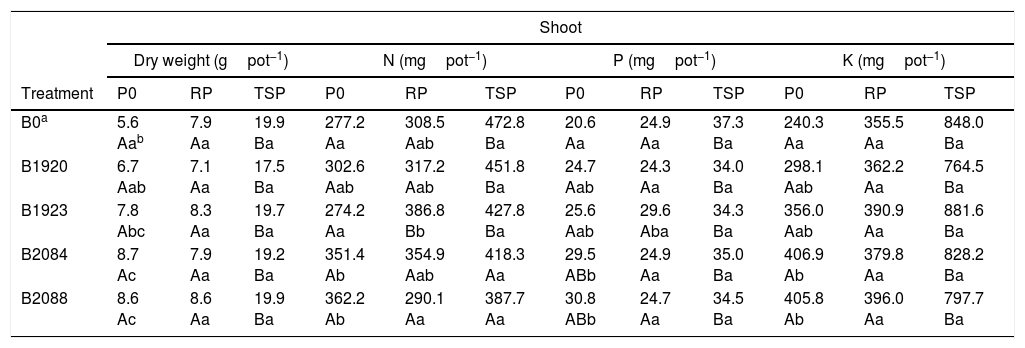
Shoot biomass (dry weight) and N P K content of pearl millet inoculated with endophytic Bacillus strains after 50 days of cultivation under greenhouse conditions with no P added (P0), Araxá rock phosphate (RP) and soluble P (TSP). Results are means SD (n=4).
| Shoot | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dry weight (gpot−1) | N (mgpot−1) | P (mgpot−1) | K (mgpot−1) | |||||||||
| Treatment | P0 | RP | TSP | P0 | RP | TSP | P0 | RP | TSP | P0 | RP | TSP |
| B0a | 5.6 Aab | 7.9 Aa | 19.9 Ba | 277.2 Aa | 308.5 Aab | 472.8 Ba | 20.6 Aa | 24.9 Aa | 37.3 Ba | 240.3 Aa | 355.5 Aa | 848.0 Ba |
| B1920 | 6.7 Aab | 7.1 Aa | 17.5 Ba | 302.6 Aab | 317.2 Aab | 451.8 Ba | 24.7 Aab | 24.3 Aa | 34.0 Ba | 298.1 Aab | 362.2 Aa | 764.5 Ba |
| B1923 | 7.8 Abc | 8.3 Aa | 19.7 Ba | 274.2 Aa | 386.8 Bb | 427.8 Ba | 25.6 Aab | 29.6 Aba | 34.3 Ba | 356.0 Aab | 390.9 Aa | 881.6 Ba |
| B2084 | 8.7 Ac | 7.9 Aa | 19.2 Ba | 351.4 Ab | 354.9 Aab | 418.3 Aa | 29.5 ABb | 24.9 Aa | 35.0 Ba | 406.9 Ab | 379.8 Aa | 828.2 Ba |
| B2088 | 8.6 Ac | 8.6 Aa | 19.9 Ba | 362.2 Ab | 290.1 Aa | 387.7 Aa | 30.8 ABb | 24.7 Aa | 34.5 Ba | 405.8 Ab | 396.0 Aa | 797.7 Ba |
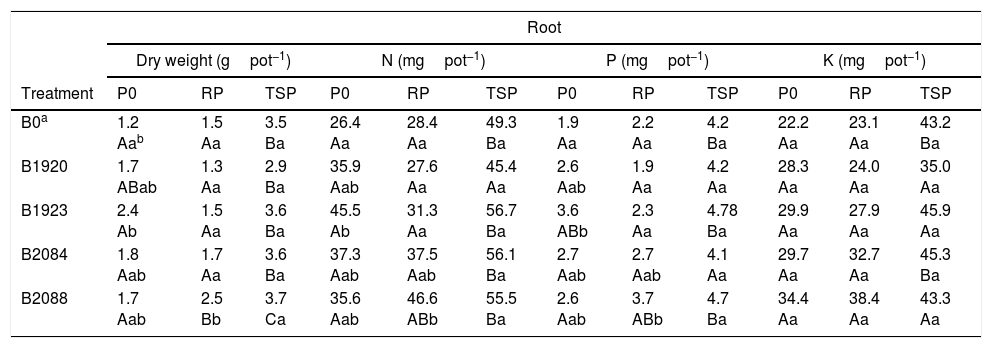
Root biomass (dry weight) and N P K content of pearl millet inoculated with endophytic Bacillus strains after 50 days of cultivation under greenhouse conditions with no P added (P0), Araxá rock phosphate (RP) and soluble P (TSP). Results are means SD (n=4).
| Root | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dry weight (gpot−1) | N (mgpot−1) | P (mgpot−1) | K (mgpot−1) | |||||||||
| Treatment | P0 | RP | TSP | P0 | RP | TSP | P0 | RP | TSP | P0 | RP | TSP |
| B0a | 1.2 Aab | 1.5 Aa | 3.5 Ba | 26.4 Aa | 28.4 Aa | 49.3 Ba | 1.9 Aa | 2.2 Aa | 4.2 Ba | 22.2 Aa | 23.1 Aa | 43.2 Ba |
| B1920 | 1.7 ABab | 1.3 Aa | 2.9 Ba | 35.9 Aab | 27.6 Aa | 45.4 Aa | 2.6 Aab | 1.9 Aa | 4.2 Aa | 28.3 Aa | 24.0 Aa | 35.0 Aa |
| B1923 | 2.4 Ab | 1.5 Aa | 3.6 Ba | 45.5 Ab | 31.3 Aa | 56.7 Ba | 3.6 ABb | 2.3 Aa | 4.78 Ba | 29.9 Aa | 27.9 Aa | 45.9 Aa |
| B2084 | 1.8 Aab | 1.7 Aa | 3.6 Ba | 37.3 Aab | 37.5 Aab | 56.1 Ba | 2.7 Aab | 2.7 Aab | 4.1 Aa | 29.7 Aa | 32.7 Aa | 45.3 Ba |
| B2088 | 1.7 Aab | 2.5 Bb | 3.7 Ca | 35.6 Aab | 46.6 ABb | 55.5 Ba | 2.6 Aab | 3.7 ABb | 4.7 Ba | 34.4 Aa | 38.4 Aa | 43.3 Aa |
Bacterial isolates B1923, B2084, and B2088 significantly enhanced millet shoot dry biomass when grown with no P fertilization (P0) compared to the control treatment. Positive effects were also observed on shoot N P K content of plants inoculated with B2084 and B2088 strains and cultivated under P0 (Table 2). Overall, in average B2084 and B2088 significantly increased shoot biomass by around 55% and N P K content by 30, 50, and 70%, respectively, compared to control treatment with no P fertilization.
On the other hand, bacterial inoculation did not increase the shoot biomass or N P K content of plants cultivated with RP and TSP. Similar results were observed for root biomass and N P content, wherein the isolates B1923 and B2088 outperformed the others when the plants were cultivated under P0 and RP, respectively (Table 3). The relative increase in root biomass, N P content was 100, 72, and 89%, respectively, for plants inoculated with B1923 and cultivated under P0 and 66, 64, and 68%, respectively, for the same parameters in plants inoculated with B2088, but fertilized with RP, when compared to the control treatment (Fig. S1 and Table 3).
DiscussionThe efficiency of phosphate solubilization by microorganisms depends on the nature of the P source and the organisms involved in the process. We selected from a previous screening in vitro, four Bacillus isolates that contrasted for Ca-P solubilization21 to be characterized for their capacity to solubilize Fe-P, that is the most abundant and least soluble P fraction in Brazilian Oxisols.3,4 In addition, we assessed siderophore and IAA production, followed by a greenhouse test to assess their effect on plant growth promotion.
Our results of soluble P released by Bacillus grown on a medium containing FePO4 varied from 34.25 to 52.22mgL−1 and were lower than the previously reported values for Ca-P solubilization by the same isolates, which ranged from 112 to 179mgL−1.21 Similar results, reported previously with different bacterial species, have shown that Fe-P and Al-P are less soluble than Ca-P.31,32 The highest Fe-P solubilization has been documented in rhizobacteria and fungi4,32,33; however, our isolates showed higher solubilization than reported for endophytic bacteria isolated from peanuts, that ranged from 13.9±0.3 to 37.4±2.0mgL−1.12 In our study, the inoculated strains decreased the growth medium pH and simultaneously increased the concentration of soluble phosphate obtained from FePO4. Abreu et al.21 measured organic acids secreted by the same bacteria, and the results suggested that culture medium acidification might be one of the mechanisms involved on Ca-P solubilization. These organic acids can chelate cations associated with phosphate, thus making P available to plants.8,34,35 However, the mechanisms behind Fe-P solubilization remain largely unknown. Highly weathered Brazilian soils under different agrosystems predominantly consist of Fe and Al oxides, independent of the vegetation, facilitating the formation of Fe-P and Al-P. This makes P often low in the soil, limiting crop yield,4 emphasizing the importance of screening for soil microorganisms that exhibit a Fe-P solubilization ability.
The effects of inoculating the four bacterial strains onto pearl millet grown with different P-sources under greenhouse conditions varied, depending on P treatment and the strain used. Compared to uninoculated controls grown with no P added, the strains B1923, B2084, and B2088 promoted increase in dry weight of shoots, and two of them, B2084 and B2088, increased shoot N P K content. Interestingly, B2084 and B2088 produced high concentrations of gluconic acid (324 and 171mM, respectively) and solubilized phosphate, with a reduction of pH, when cultivated in Ca-P21 or Fe-P (Table 1) media. Gluconic acid production has been frequently described as the most efficient mechanism related to P solubilization by bacteria. For instance, Pisum sativum inoculated with strains of endophytic Pseudomonas fluorescens, which is capable of producing gluconic acid (14–169mM) and solubilizing phosphate, presented higher growth compared to uninoculated plants.36 These strains probably exude gluconic acid into the rhizosphere of the inoculated plants, thus playing an important role in the solubilization of natural P fixed in soil particles and allowing the plant to subsequently assimilate the soluble P.
On the other hand, although the inoculation of plants with strains B1923, B2084, and B2088 and cultivated with Araxá RP increased shoot dry weight, this effect was not statistically significant, indicating that the amount of P solubilized by these bacteria was probably not sufficient to significantly increase the shoot parameters evaluated in our shot-term experiment. Long-term field experiments are likely to show more clearly the effect of RP solubilization by these bacteria on P accumulation in grain and shoot biomass.
In our study, two Bacillus strains also caused a significant increase in root dry weight and N P content in plants cultivated in soil without P added (B1923) or amended with RP (B2088). The difference between the performance of the two isolates in the greenhouse experiment was probably a consequence of their different P-solubilizing mechanisms, measured in vitro, since B2088 is an efficient Ca-P solubilizer21 and B1923 showed higher Fe-P solubilization (Table 1). This characteristic could be important in the selection of bacteria to be used as inoculants for plants cultivated in different soils or soils amended with RP. It is well known that the rate of RP solubilization is influenced by different physicochemical characteristics that depend on the source material and particle size. Araxá is an RP of igneous or metamorphic origin that exhibits a high crystallization level and low citric acid solubility,37 which makes it difficult to solubilize, particularly in the first year of cultivation. In addition, fluoride present in Araxá RP limited the solubilization by Aspergillus niger by negatively affecting metabolic processes involved in phosphate solubilization, such as the growth of the fungus, its citric acid production, and medium acidification.38
Most of the Fe-P-solubilizing strains also produced siderophores (Table 1), which are iron-binding compounds that can chelate ferric iron and thereby make it available for microbial and plant cells. It is known that siderophores have the potential for plant pathogen biocontrol,13,39 as well as the potential to solubilize iron from minerals, thus increasing the availability of P to plants.40,41–43 Recently, Ghosh et al. (2016)44 reported high siderophore production by three Burkholderia strains that produced soluble phosphate when ferric phosphate was used as the sole phosphate source. However, the specific contribution of siderophores to P solubilization mechanisms in soil is yet to be elucidated.45
In addition to the siderophore-producing ability, an in vitro screening revealed that all Bacillus isolates, when grown with tryptophan in the culture medium, were able to produce IAA, in a range between 3.2 and 61.6μgmL–1, revealing a substantial variability among isolates (Table 1). Other species of Bacillus and Lactobacillus presented similar results, with reported IAA production between 30 and 60μgmL–110 and 0.81 and 86.82μgmL–1.13 IAA production is widespread among bacteria and has been used as a criterion for selecting effective PGP bacteria.46 Bacteria that produce IAA and promote growth were described for different crops such as maize, peanut, wheat, soybean, and rice.46–49 In general, high levels of bacterial IAA are associated with lateral and adventitious root formation, while low IAA levels often stimulate root elongation.50 An extended root surface combined with a more efficient P solubilization ability improves nutrient acquisition and water uptake. IAA-producing bacteria also stimulate shoot growth; however, it is not clear whether IAA has a direct effect on shoot growth or an indirect effect, i.e., by stimulating the root system and consequently enhancing nutrient and water uptake.51
Most PGPB endophytes are facultative, changing between the free-living and endophytic stages depending on different factors such as the soil, microbial competitors, and plant nutrients.52 This facultative endophytic lifestyle enables multi-traits PGP, resulting in more competitive bacteria than rhizospheric microorganism as they can be inoculated in seeds and spread systemically through the plant reducing the need of continuous inoculations.53
In conclusion, we observed a positive effect of the inoculation of three endophytic Bacillus (B1923, B2084 and B2088) on pearl millet growth and nutrient uptake, indicating that these strains have potential to be used as plant inoculants in highly weathered tropical soils. B1923 strain was able to produce IAA and solubilize Fe-P in vitro and this trait was expressed under phosphate limiting conditions in the soil (no P added) resulting in enhanced shoot and root dry weight and root N P content of pearl millet plants. Strains B2084 and B2088, despite of solubilize less Fe-P than B1923, showed positive performance on biomass production and accumulation of nutrients N P K in the shoot, which represents the most relevant response of these microorganisms in the productive environment. Based on this, we indicated that the strains B2084 and B2088 have higher potential to be microbial biofertilizer candidates for commercial applications in the absence of fertilization.
Conflicts of interestThe authors declare that they have no conflict of interest.
This work was supported by Fapemig (Grant No. APQ-01867-14), MPCPAgro (CNPq 465133/2014-4, Capes) and Embrapa (Grant No. 01.13.05.001.02-04).
