


The bioconversion of cellulosic wastes into high-value bio-products by saccharification and fermentation processes is an important step that can reduce the environmental pollution caused by agricultural wastes. In this study, enzymatic saccharification of treated and untreated date palm cellulosic wastes by the cellulases from Geobacillus stearothermophilus was optimized. The alkaline pre-treatment of the date palm wastes was found to be effective in increasing the saccharification percentage. The maximum rate of saccharification was found at a substrate concentration of 4% and enzyme concentration of 30 FPU/g of substrate. The optimum pH and temperature for the bioconversions were 5.0 and 50°C, respectively, after 24h of incubation, with a yield of 31.56mg/mL of glucose at a saccharification degree of 71.03%. The saccharification was increased to 94.88% by removal of the hydrolysate after 24h by using a two-step hydrolysis. Significant lactic acid production (27.8mg/mL) was obtained by separate saccharification and fermentation after 72h of incubation. The results indicate that production of fermentable sugar and lactic acid is feasible and may reduce environmental pollution by using date palm wastes as a cheap substrate.
At present, the conversion of low-value agriculture wastes into valuable commodities, energy, chemicals and microbial protein by saccharification and fermentation processes is not economically feasible, largely due to the costs of cellulosic materials and cellulolytic enzymes, as well as technical problems associated with cellulose saccharification.1–5 There is an increased interest in using thermophilic bacteria Geobacillus stearothermophilus for the production of cellulases and separate saccharification and fermentation of lignocellulosic biomass due to their higher operating temperatures and broad substrate range.6,7 The complete cellulose hydrolysis can be achieved by a combination of three types of cellulases: endoglucanases, which cleave internal glucosidic bonds; exoglucanases, which cleave cellobiosyl units from the ends of cellulose; and glucosidase, which cleaves glucose units from cello-oligosaccharides. Endoglucanases are characterized by their activity toward substituted cellulose derivatives such as carboxymethylcellulose, while exocellulases have been operationally defined by their ability to degrade microcrystalline cellulose.8
Out of the total global production of 7.4 million tons of dates, 5.4 million come from the Arab world.9 The Kingdom of Saudi Arabia (KSA) is a major date-producing country and is ranked second in the world. The total area planted with date palm trees is about 162,000 hectares, while the number of palm trees has reached nearly 23 million.10 Besides fruits, date palm also provides a large number of other products that have a wide range of applications. It can be used as a raw material for certain industrial purposes. Practically all parts of date palm are usable such as trunk, leaves (whole leaves, midribs, leaflets and spines, and the sheath at the leaf base), reproductive organs (spathes, fruit stalks, spikelets and pollens) and many of their extracts.11 Date palm (Phoenix dactylifera) has high values of cellulose (45.3%), hemicellulose, (29.13%), and lignin (25.82%).12 Lignocellulosics cannot be saccharified by cellulases to yield sugar unless they are processed through mechanical, physical, and chemical pre-treatments to remove the inhibitory lignin complex, to reduce the crystallinity and degree of polymerization of cellulose, to increase the surface area available for the enzymes, and to enhance the susceptibility of the substrates to enzymes.13–16 Lactic acid is a valuable organic acid due to its broad applications in pharmaceutical, leather, and food industries, and its potential for the production of biodegradable poly-lactic acid—an environmentally friendly alternative to plastic.17–19 In this study, an optimized production of glucose syrup by enzymatic saccharification of treated and untreated date palm wastes was investigated. Besides this, an attempt was also made to produce lactic acid by separate saccharification and fermentation.
Materials and methodsEnzyme sourceThe bacterial species Geobacillus stearothermophilus Y-1, as a source of cellulases, was isolated from the Najran region, KSA. Culture from agar plate was inoculated into a 50mL tube containing 5mL of nutrient broth and incubated at 50°C in an orbital shaker at 200rpm. This culture was used to inoculate a 250mL Erlenmeyer flask containing 50mL Bushnell Haas medium (BHM).20 The production medium (BHM) consisted of: MgSO4·7H2O (0.2g/L), K2HPO4 (1g/L), KH2PO4 (1g/L), yeast extract (1.0g/L), FeCl3·6H2O (0.05g/L), CaCl2 (0.02g/L), and Tween 80 (0.2%). It was supplemented with 2.0% alkaline-treated date palm leaves as a carbon source. The optimum conditions for cellulase production, when the fermentation period was extended up to 48h, were as follows: cultivation temperature 45°C, pH 7.0, and agitation rate 200rpm (data not shown). The cells and insoluble materials were removed by centrifugation at 10,000rpm for 10min and the cell-free supernatant was used as the enzyme source. The cellulases system contained: FPase 8.105U/mL, CMCase, 12.84U/mL and β-glucosidase 3.74U/mL.
Pre-treatments of date palm wastesDate palm cellulosic wastes (leaves, leaf bases, and fibers) were collected from a date palm plantation in Abha city, KSA and used as the cellulosic substrate. The wastes were ground and pre-treated by two methods: (1) Alkaline pre-treatment: 2N NaOH at 30°C for 48 h21 and (2) acid-steam pre-treatment: 1% H2SO4, 120°C for 100min.22 After the treatment, the wastes were washed thoroughly with tap water until neutralized and oven dried at 70°C. Dried materials were ground through a Wiley Mill (Model 2 Thomas Co., USA) to obtain a particle size ≤1mm. Determination of cellulose, hemicellulose and lignin contents in the treated and untreated wastes was performed according to Saura-Calixto et al.23
Enzymatic saccharification of date palm cellulosic wastesEnzymatic hydrolysis of date palm cellulosic wastes was carried out following the methods of Holtzapple et al.24 Briefly, 2% cellulosic waste was mixed with an appropriate amount of enzyme (20 FPU/g of substrate) in a 100-mL Erlenmeyer flask containing 20mL acetate buffer (pH 5.0), and sodium azide (0.3g/L) was added to inhibit microbial contamination. The enzymatic hydrolysis was carried out for 24h at 50°C using a shaking incubator (100rpm). After the saccharification period, the reaction mixture was centrifuged at 4000rpm for 30min to remove unhydrolyzed substrate and the supernatant was subjected to glucose determination. The effects of incubation temperature, pH, substrate concentration, enzyme concentration, and incubation time on saccharification and glucose production were investigated.
Reducing sugars assayThe amount of reducing sugars released by the enzymatic hydrolysis was estimated by dinitrosalicylic acid (DNS) method.25 The sample (1.0mL) was mixed with 2mL of DNS reagent. The tubes were then heated in a boiling water bath for 5min, after cooling at room temperature; the absorbance was measured at 540nm. The amount of the released reducing sugar was calculated by using a standard curve of glucose, and expressed as mg/mL. The percentage saccharification was calculated using the equation of Mandels and Sternberg26 as follows:
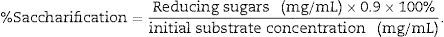
The factor 0.90 was used to convert polysaccharide to monosaccharide accounting for water uptake during hydrolysis. All experiments were carried out in triplicates.
Lactic acid fermentationStrain and growth mediumThe culture of Lactobacillus delbrueckii subsp. Lactis (B. 01357), a homo fermentative lactic acid producer, was utilized in this study. The strain was obtained from the National Collection of Agriculture and Industrial Microorganisms, Budapest, Hungary. The medium used for cell growth contained (g/L): peptone 10.0, meat extract 10.0, yeast extract 5.0, d-glucose 20.0, tween-80 1.0, K2HPO4 2.0, sodium acetate 5.0, tri-ammonium citrate 2.0, MgSO4·7H2O 0.2, MnSO4·4H2O 0.05. The cells from the stock culture were transferred to a sterile growth medium and incubated at 45°C for 24h. The obtained culture was used as inoculum for the successive fermentation process.
Lactic acid production by separate saccharification and fermentationAt first, the cellulosic substrate was hydrolyzed under optimum conditions. The hydrolysate (produced after enzymatic saccharification) was supplemented with nutrients (6% yeast extract, 0.167% sodium acetate, 0.167% (NaPO3)n. 0.1% MgSO4·7H2O, 0.005% FeSO4·7H2O. 0.005% MnSO4·H2O) made to 50mL (pH 5.0) in a 100-mL flask. CaCO3 (5%) was added to prevent acidification. The inoculum size of Lactobacillus delbrueckii B. 01357 was 10% (v/v) and the fermentation was carried out at 50°C for 5 days.

The yield of lactic acid from glucose (%)=concentration of lactic acid produced (g/L)×100%/[initial concentration of glucose in the cellulosic hydrolysate (g/L)]−[final concentration of remaining glucose in the fermentation broth (g/L)].
Lactic acid determinationLactic acid concentration was determined as described by Taylor,27 using pure lactic acid as standard, and concentration was expressed as mg/mL.
Statistical analysisThe obtained data were subjected to analysis of variance using the MSTATC program. The least significant difference (LSD) at p≤0.05 was used to detect differences between the treatments.
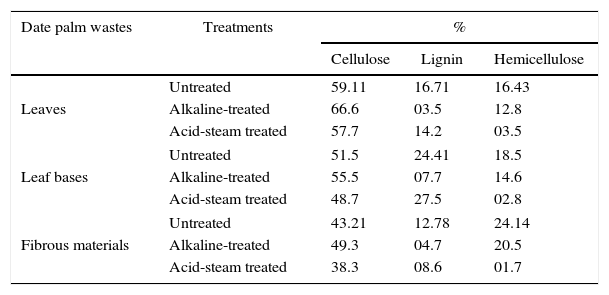
Results and discussionChemical analysis of treated and untreated date palm cellulosic wastesThe amount of carbohydrate polymers and lignin varies among plants, and there are also variations in the constituents in a single plant depending on its age, stage of growth, and other conditions. Cellulose is the dominant structural polysaccharide in plant cell walls, followed by hemicelluloses and lignin.28 The changes in composition of date palm cellulosic wastes were compared before and after the alkaline or acid-steam pre-treatments (Table 1). The maximum lignin content (24.41%) was recorded for leaf bases, followed by leaves (16.71%), and minimum for fibrous date material (12.78%). Date palm leaves contained the highest value of cellulose (59.11%) followed by the leaf bases (51.5%) and trace amounts in fibrous material. For the hemicelluloses content, leaves and leaf bases contained approximately the same amount (16–18%) and the highest value was recorded for fibrous materials (24.14%). These results do not match with those obtained by Ghosh et al.,29 who reported lignin, cellulose, and hemicellulose contents of date palm leaves as 15.3%, 58%, and 20%, respectively. However, Al-Haidary et al.30 reported the main components of the date palm fiber and petiole as cellulose (44.17%), hemicellulose (21.95%), and lignin (12.75%). Conversely, Barreveld11 reported that the main three components in the fibrous parts of date palm, holocellulose, cellulose, and lignin were 48%, 28%, and 28.1%, for leaflets, while for leaf bases, were 54.5%, 22.5%, and 27%, respectively.
Chemical composition of treated and untreated date palm cellulosic wastes.
| Date palm wastes | Treatments | % | ||
|---|---|---|---|---|
| Cellulose | Lignin | Hemicellulose | ||
| Leaves | Untreated | 59.11 | 16.71 | 16.43 |
| Alkaline-treated | 66.6 | 03.5 | 12.8 | |
| Acid-steam treated | 57.7 | 14.2 | 03.5 | |
| Leaf bases | Untreated | 51.5 | 24.41 | 18.5 |
| Alkaline-treated | 55.5 | 07.7 | 14.6 | |
| Acid-steam treated | 48.7 | 27.5 | 02.8 | |
| Fibrous materials | Untreated | 43.21 | 12.78 | 24.14 |
| Alkaline-treated | 49.3 | 04.7 | 20.5 | |
| Acid-steam treated | 38.3 | 08.6 | 01.7 | |
[LSD (p≤0.05): cellulose 0.144652, lignin 0.076727, hemicellulose 0.058289].
It is clear from the results shown in Table 1 that alkaline pre-treatment was useful for selectively reducing lignin content in all investigated wastes, compared with acid-steam pre-treatment. The alkaline treatments removed about 79.05%, 68.4%, and 63.22% of the lignin in leaves, leaf bases, and fibrous material, respectively. An increase in cellulose content up to 12.67%, 7.7%, and 14.1% in leaves, leaf bases, and fibrous materials, respectively, was observed after alkaline pre-treatment. These results are consistent with those described by Carrasco et al.22 and Mussatto et al.31 These authors reported that alkali treatment with NaOH is one of the most common methods to delignify agricultural residues that causes swelling, leading to an increase in the internal surface area, decrease in the degree of polymerization and crystallinity, separation of structural linkages between lignin and carbohydrates, and disruption of the lignin structure. The disadvantage of this technique is that it also degrades some fraction of the hemicellulose. Conversely, acid-steam pre-treatment has little effect in reducing lignin content, while it lowers cellulose content. The main effect was an observed reduction of hemicellulose content to 79–93%. Carvalheiro et al.,32 and Kumar et al.33 reported that dilute acid under steam is the most widely used method for breakdown of hemicellulose, but the limitations of this method include incomplete disruption of the lignin–carbohydrate matrix and generation of compounds that may be inhibitory to microorganisms.34
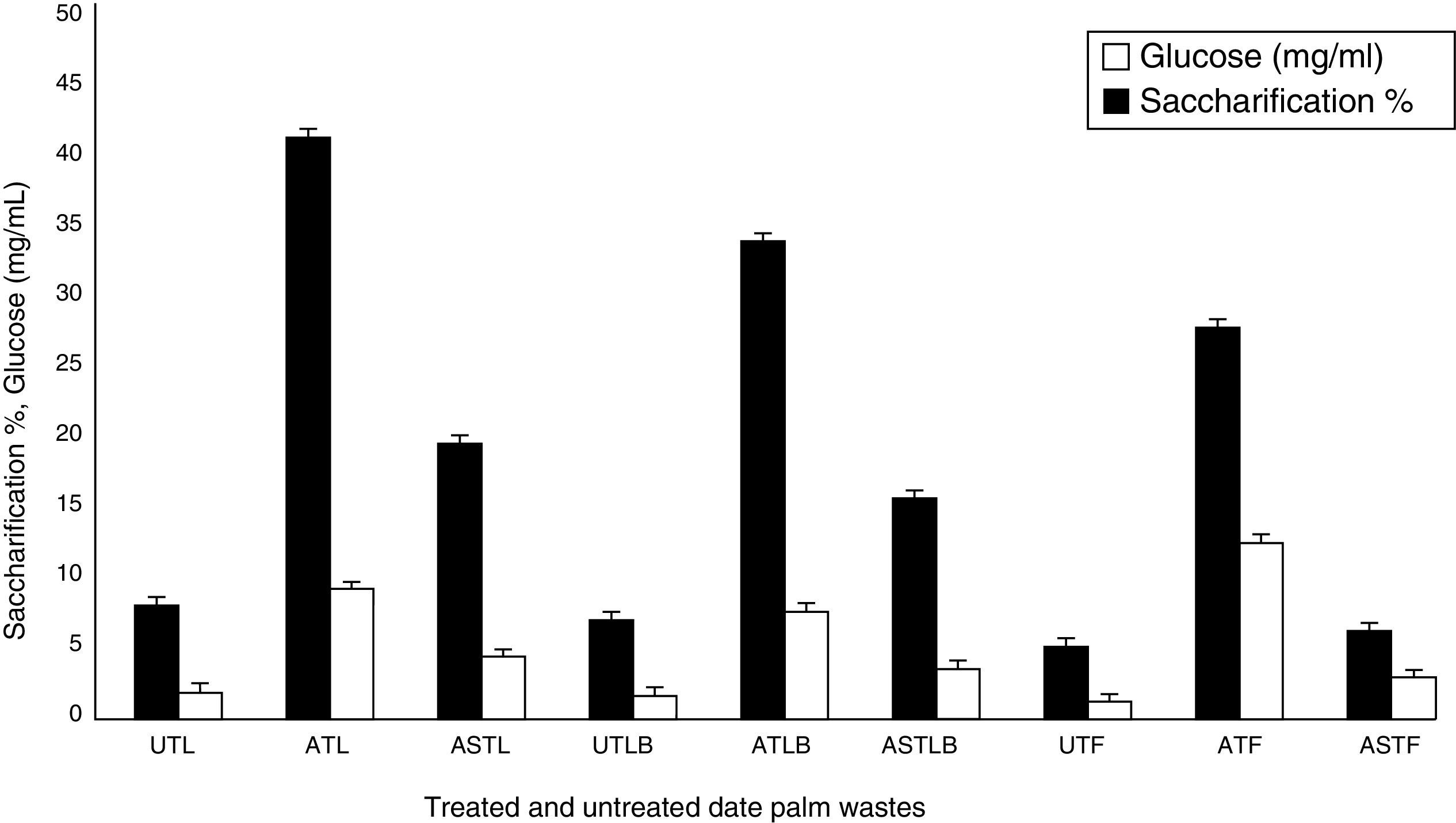
Effect of pre-treatments on date palm wastes for saccharification and glucose productionThe effectiveness of different pre-treatments to alter cellulose structure and to reduce hemicellullose and lignin content in date palm wastes for subsequent use as substrate for enzymatic saccharification was investigated. The results show that untreated substrates were poorly attacked by the cellulase enzymes compared with the pre-treated ones (Fig. 1). The maximum saccharification degrees for untreated substrates were 8.01%, 6.96%, and 5.08% for leaves, leaf bases, and fibrous materials, respectively. Glucose production reached 1.77mg/mL, 1.55mg/mL and 1.14mg/mL for leaves, leaf bases, and fibrous materials, respectively. The low saccharification percentages may be attributed to the natural cellulosic wastes that possess a high amount of lignin, which makes difficult for the large protein molecules of enzymes to penetrate into the tight network constituting the plant cell wall. Therefore, pre-treatment of the lignocellulosic substrates is essential for efficient enzymatic hydrolysis.35,36
![Effect of pre-treatments on saccharification and glucose production by date palm wastes (UTL, untreated leaves; ATL, alkaline-treated leaves; ASTL, acid-steam-treated leaves; UTLB, untreated leaf bases; ATLB, alkaline-treated leaves bases; ASTLB, acid-steam-treated leaf bases; UTF, untreated fibrous; ATF, alkaline-treated fibrous; ASTF, acid-steam-treated fibrous) [LSD (p≤0.05): saccharification 0.035754, glucose 0.027816].](https://static.elsevier.es/multimedia/15178382/0000004700000001/v1_201602040959/S1517838215000167/v1_201602040959/en/main.assets/gr1.jpeg?xkr=ue/ImdikoIMrsJoerZ+w997EogCnBdOOD93cPFbanNerEcKXq/4gYMpNkXqSlVDY0lXCTapXrEOq5lCiG1bYbVRoz4b5rC/mtt5Z76dY4YaRNtQekK/QrNbPG6UsDyr4ZJ64xH79Pa0LNZ49YBb3E4vTKT/q/G8gNaEWafrN2gcQgUJMpSmzQR/S+VDYQeOXZiBdlYMrk+DESQxFCcCAU3L63HLMMXIWwsqgSnGzd7KylfPT7WP2H/0cJNWhxD0+tuqlh/JIhw715BOEieuMrCPbFKH4FkbuopqX9PPWfuo= "Effect of pre-treatments on saccharification and glucose production by date palm wastes (UTL, untreated leaves; ATL, alkaline-treated leaves; ASTL, acid-steam-treated leaves; UTLB, untreated leaf bases; ATLB, alkaline-treated leaves bases; ASTLB, acid-steam-treated leaf bases; UTF, untreated fibrous; ATF, alkaline-treated fibrous; ASTF, acid-steam-treated fibrous) [LSD (p≤0.05): saccharification 0.035754, glucose 0.027816].")
Effect of pre-treatments on saccharification and glucose production by date palm wastes (UTL, untreated leaves; ATL, alkaline-treated leaves; ASTL, acid-steam-treated leaves; UTLB, untreated leaf bases; ATLB, alkaline-treated leaves bases; ASTLB, acid-steam-treated leaf bases; UTF, untreated fibrous; ATF, alkaline-treated fibrous; ASTF, acid-steam-treated fibrous) [LSD (p≤0.05): saccharification 0.035754, glucose 0.027816].
The optimum amount of sugar was obtained with alkaline-treated substrates. The maximum saccharification rate (41.40%) and glucose production (9.2mg/mL) were obtained for alkaline-treated leaves (ATL). Alkaline treatment may increase the available surface area and reduce the cellulose crystallinity in leaves compared with leaf bases. These results concord with those of Uzunlu et al.,37 who found that alkaline pre-treatment of poppy stalks resulted in an increased saccharification degree. Moreover, Akhtar et al.35 reported that pre-treatment of wheat straw, rice straw, and bagasse with 2% NaOH was most effective for increasing the enzymatic saccharification by Bacillus subtilis cellulases, with saccharification rates of 33.0%, 25.5%, and 35.5%, respectively. On the other hand, acid-steam substrates appeared to be the next alternative. The maximum saccharification degree (19.57%) and glucose production (4.37mg/mL) were observed with date palm leaves, followed by leaf bases, 15.66% and 3.47mg/mL, respectively. These results are in contrast with those obtained by Saha et al.,38 who reported that the conversion degree for enzymatic hydrolysis of cellulosic wastes pre-treated by diluted H2SO4 at an elevated temperature was not less than 80%. These results indicate that the main factor affecting enzymatic saccharification was the nature of the alkaline pre-treatment, which can dissolve part of the hemicelluloses and lignin and lead to swelling of the cellulose. Therefore, alkaline treated date palm leaves were chosen as the best substrate for the subsequent saccharification experiments.
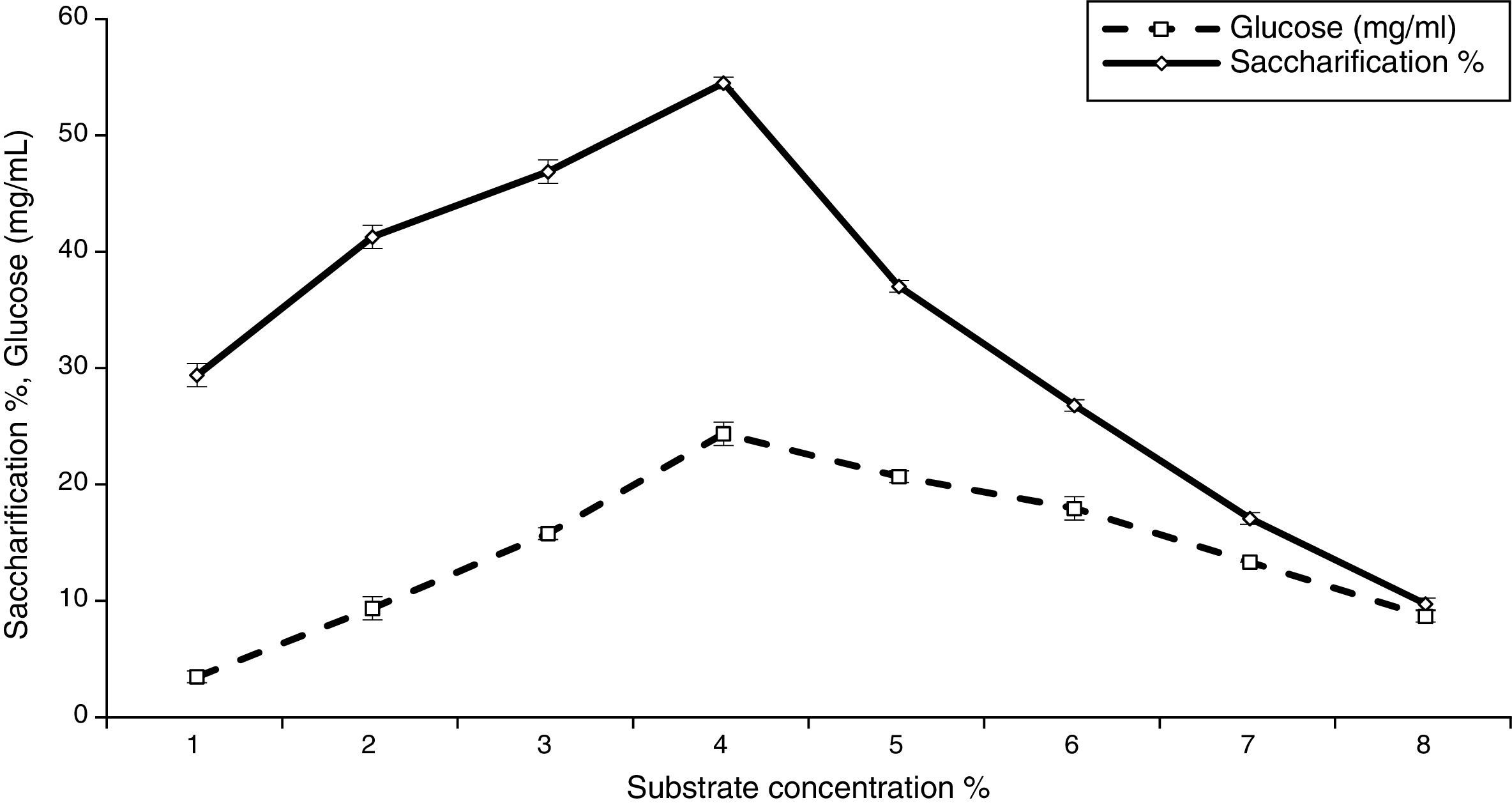
Effect of substrate concentration on saccharification and glucose productionThe results illustrated in Fig. 2 indicate that as the concentration of alkaline-treated date palm leaves (ATDPL) was increased, the sugar production and saccharification percentages were increased up to 4% for ATDPL. Similar results were reported by Singh et al.,39 who obtained the maximum glucose production with 4% alkaline-treated bagasse. Furthermore, Akhtar et al.35 reported the optimum substrate concentration for maximum enzymatic saccharification of alkaline pre-treated wheat straw, rice straw, and bagasse as 4%, while using Bacillus subtilis cellulases. The increase in substrate concentration decreased the saccharification percentage, which might be caused by poor stirring, enzyme inhibition by saccharification products, and decreased synergistic action between cellulases enzymes, as mentioned by Krishna and Chowdary40 and Wen et al.41 However, Mahamud and Gomes36 reported maximum hydrolysis of alkaline-treated sugarcane bagasse at a 2.5% substrate concentration. Furthermore, Zhang et al.42 reported that the feasible substrate concentration for saccharification of steam explosion pre-treated corn stover was about 3% and an increase in substrate concentration from 4 to 6% limited the rate of hydrolysis. Moreover, Ouyang et al.43 obtained maximum saccharification yield (90%) when used 3% corncob residue concentration.
Effect of enzyme concentration on saccharification and glucose production![Effect of substrate concentration on saccharification and glucose production [LSD (p≤0.05): saccharification 0.025365, glucose 0.024607].](https://static.elsevier.es/multimedia/15178382/0000004700000001/v1_201602040959/S1517838215000167/v1_201602040959/en/main.assets/gr2.jpeg?xkr=ue/ImdikoIMrsJoerZ+w997EogCnBdOOD93cPFbanNerEcKXq/4gYMpNkXqSlVDY0lXCTapXrEOq5lCiG1bYbVRoz4b5rC/mtt5Z76dY4YaRNtQekK/QrNbPG6UsDyr4ZJ64xH79Pa0LNZ49YBb3E4vTKT/q/G8gNaEWafrN2gcQgUJMpSmzQR/S+VDYQeOXZiBdlYMrk+DESQxFCcCAU3L63HLMMXIWwsqgSnGzd7KylfPT7WP2H/0cJNWhxD0+tuqlh/JIhw715BOEieuMrCPbFKH4FkbuopqX9PPWfuo= "Effect of substrate concentration on saccharification and glucose production [LSD (p≤0.05): saccharification 0.025365, glucose 0.024607].")
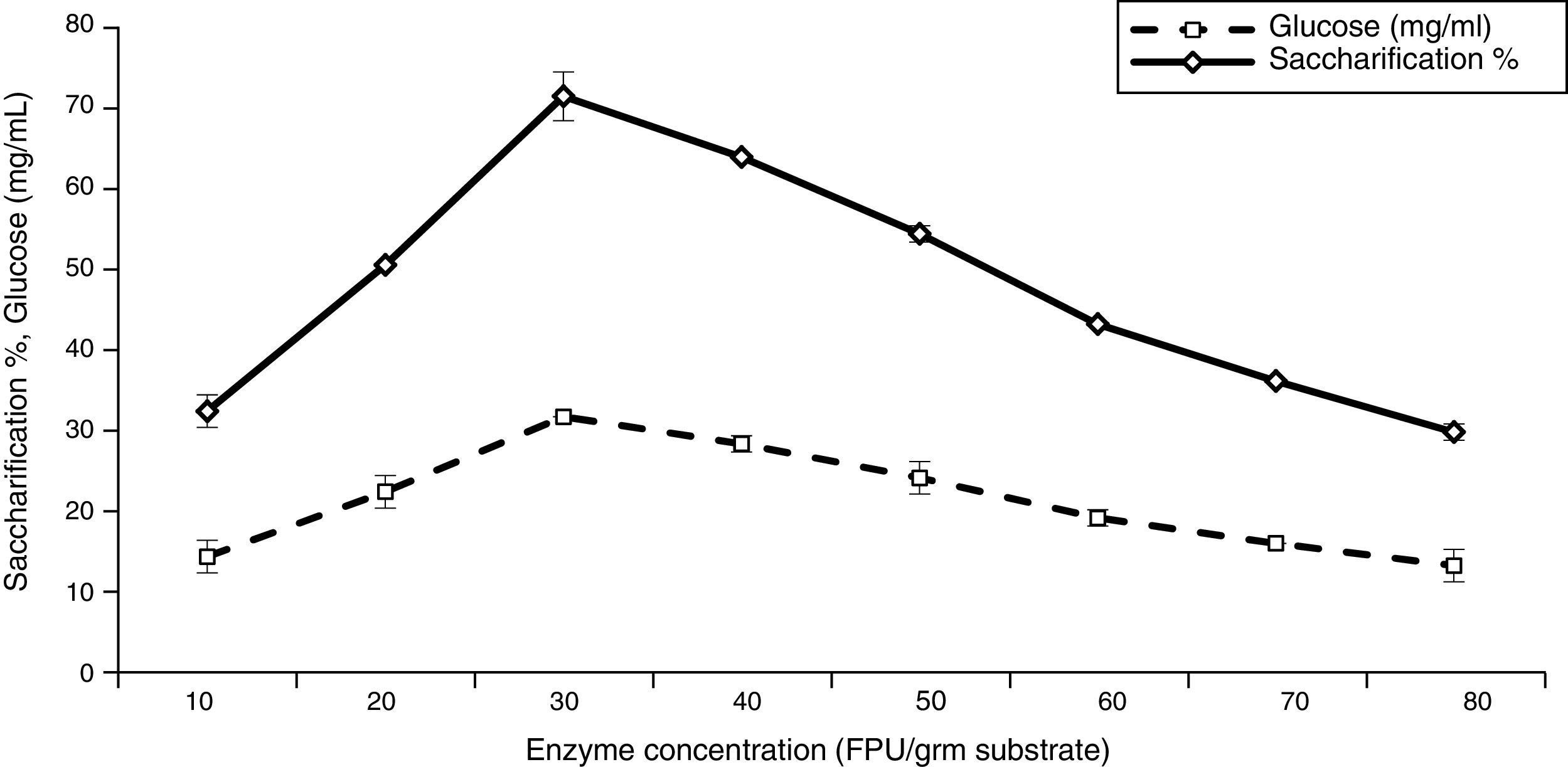
The effects of enzyme concentration on the saccharification percentage and sugar production was investigated and found to be in a range of 10–80FPU/g substrate (Fig. 3). The results revealed that the saccharification degree depended strongly on the enzyme-substrate ratio. The maximum saccharification degree and sugar production were gradually increased up to 71.03% and 31.57mg/mL, respectively, at 30FPU/g substrate. A further increase in the enzyme load did not improve sugar yield. High cellulase concentrations may counteract the saccharification by increasing the rate of transglycosylation reactions44 along with hydrodynamic instability, improper mixing, and suspension of slurry.35 As the cost of cellulase contributes significantly to the cost of the hydrolysis process, the enzyme dosage should be minimized as much as possible.45 Furthermore, Krishna and Chowdary40 found a cellulase concentration of 120 FPU/g substrate as optimal for enzymatic saccharification; and an increase in enzyme content from 40 to 120FPU increased the yield only by 10%. Conversely, Zhang et al.42,46 reported that the optimum enzyme concentrations for saccharification of ammonia pre-treated corncob and steam explosion pre-treated corn stover were 50 and 20FPU/g substrate, respectively. In contrast, Ouyang et al.43 suggested that the enzyme dosage for saccharification should not to exceed 5–8FPU/g substrate. They obtained maximum saccharification yield (90%) when the enzyme concentration was 8FPU/g substrate of corncob residue.
Effect of pH value on saccharification and glucose production![Effect of enzyme substrate ratio on saccharification and glucose production [LSD (p≤0.05): saccharification 0.023826, glucose 0.0261].](https://static.elsevier.es/multimedia/15178382/0000004700000001/v1_201602040959/S1517838215000167/v1_201602040959/en/main.assets/gr3.jpeg?xkr=ue/ImdikoIMrsJoerZ+w997EogCnBdOOD93cPFbanNerEcKXq/4gYMpNkXqSlVDY0lXCTapXrEOq5lCiG1bYbVRoz4b5rC/mtt5Z76dY4YaRNtQekK/QrNbPG6UsDyr4ZJ64xH79Pa0LNZ49YBb3E4vTKT/q/G8gNaEWafrN2gcQgUJMpSmzQR/S+VDYQeOXZiBdlYMrk+DESQxFCcCAU3L63HLMMXIWwsqgSnGzd7KylfPT7WP2H/0cJNWhxD0+tuqlh/JIhw715BOEieuMrCPbFKH4FkbuopqX9PPWfuo= "Effect of enzyme substrate ratio on saccharification and glucose production [LSD (p≤0.05): saccharification 0.023826, glucose 0.0261].")
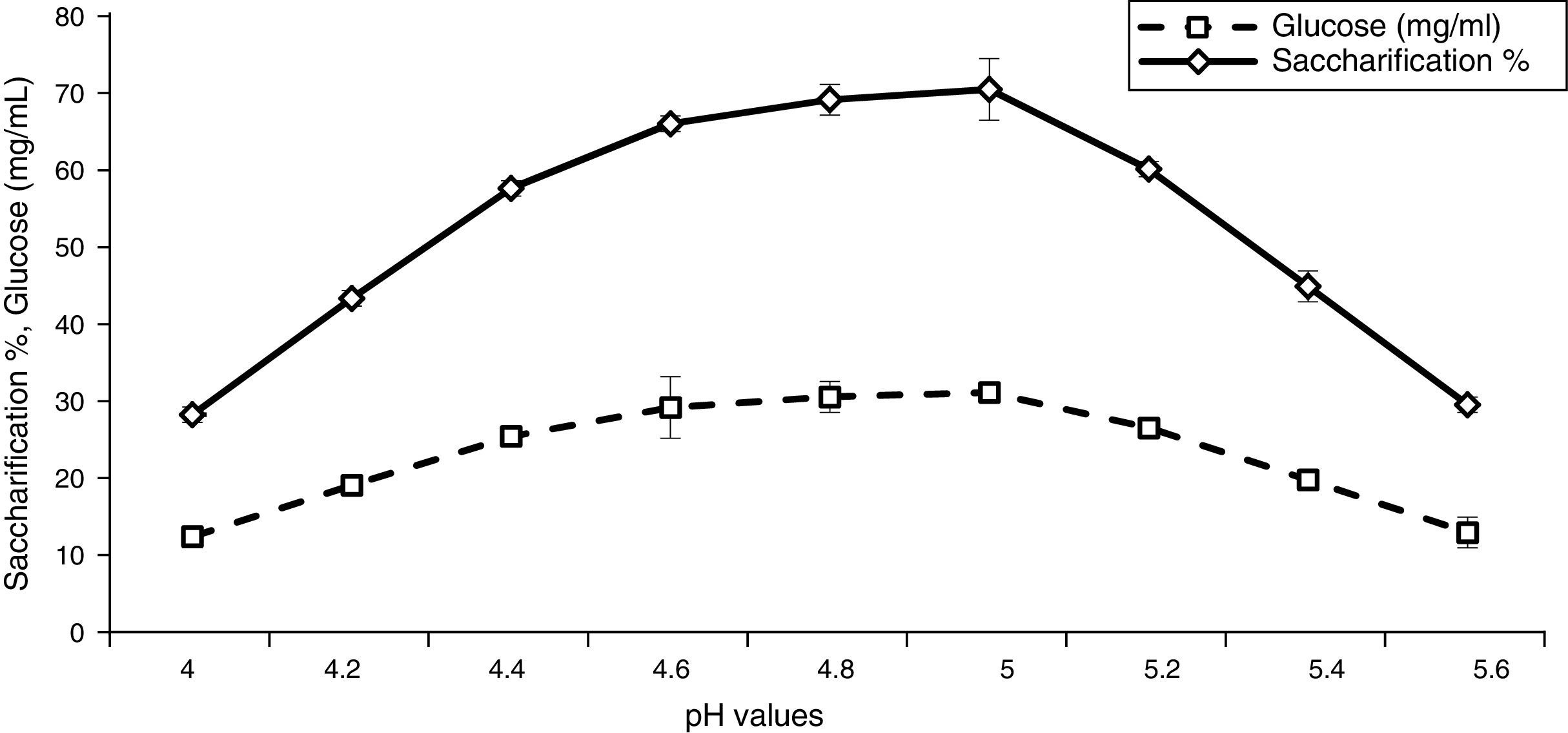
The results illustrated in Fig. 4 indicate that the sugar and saccharification percentages were increased significantly at pH 5.0. These results are consistent with few earlier reports such as Singh et al.39 and Krishna and Chowdary.40 They found that pH has a significant effect on the hydrolytic behavior of cellulases; the hydrolytic reaction is possible only after enzyme–substrate complex formation, and the effect of pH on the both adsorption and hydrolysis is similar that usually occur at around pH 4.8. Moreover, Mahamud and Gomes36 reported maximum hydrolysis of alkali-treated sugarcane bagasse at pH 5.0. Furthermore, Baig et al.13 reported that pH 6.0 was optimal for enzymatic saccharification of steam-treated leaves and pseudo-stem of banana. Conversely, Karmakar and Ray47 reported pH 7.0 as optimal for saccharification of various agro-wastes, such as orange peel, sugarcane bagasse, dried flower, water hyacinth, and coconut shell, while using cellulase enzymes from Rhizopus oryzae PR 7. They also observed that the optimal pH was higher than the acidic pH optima of CMCase from Trichoderma viride. However, similar to that reported from Bacillus subtilis.35
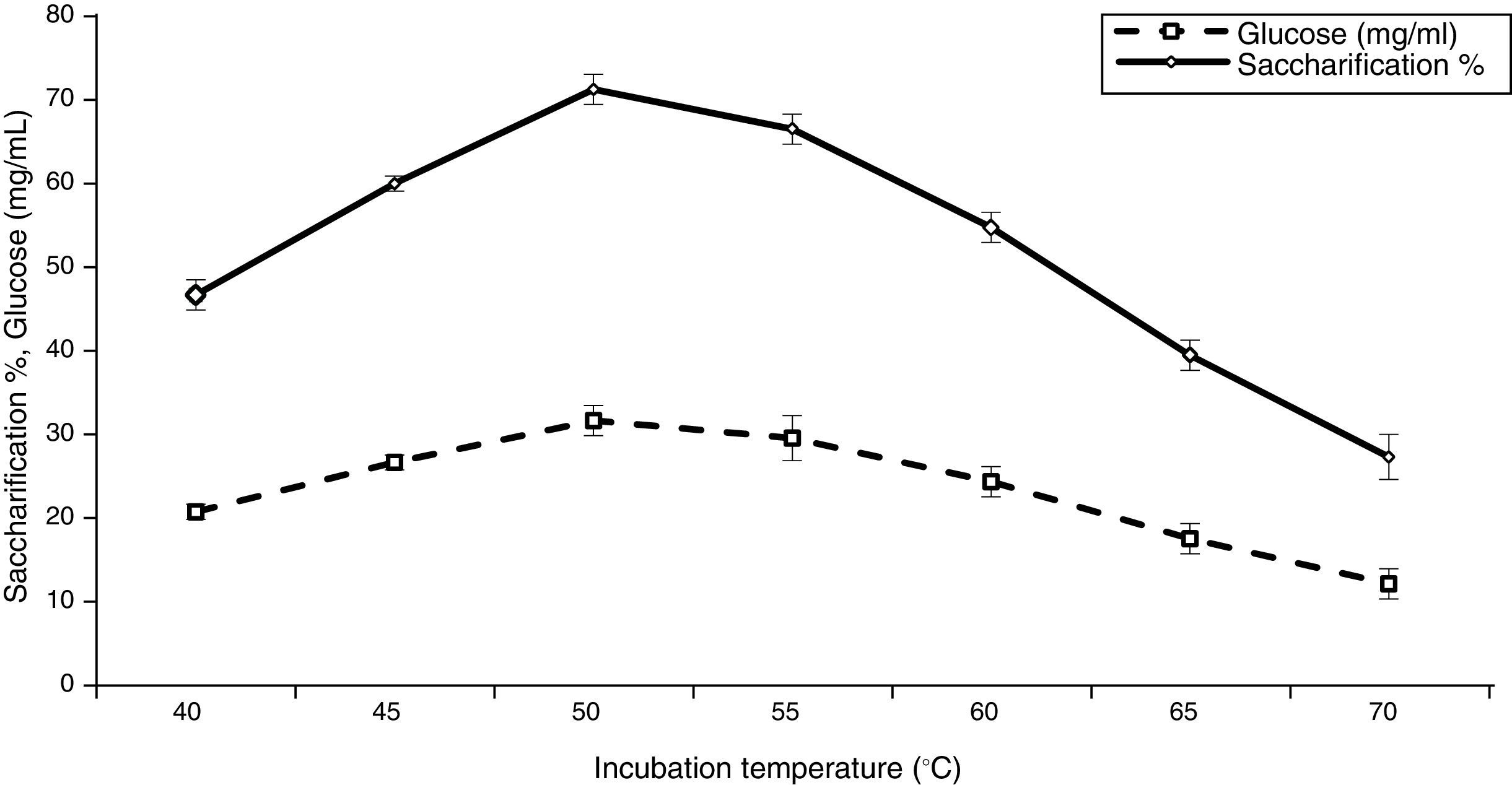
Effect of temperature on saccharification and glucose production![Effect of pH value on saccharification and glucose production [LSD (p≤0.05): saccharification 0.031768, glucose 0.050895].](https://static.elsevier.es/multimedia/15178382/0000004700000001/v1_201602040959/S1517838215000167/v1_201602040959/en/main.assets/gr4.jpeg?xkr=ue/ImdikoIMrsJoerZ+w997EogCnBdOOD93cPFbanNerEcKXq/4gYMpNkXqSlVDY0lXCTapXrEOq5lCiG1bYbVRoz4b5rC/mtt5Z76dY4YaRNtQekK/QrNbPG6UsDyr4ZJ64xH79Pa0LNZ49YBb3E4vTKT/q/G8gNaEWafrN2gcQgUJMpSmzQR/S+VDYQeOXZiBdlYMrk+DESQxFCcCAU3L63HLMMXIWwsqgSnGzd7KylfPT7WP2H/0cJNWhxD0+tuqlh/JIhw715BOEieuMrCPbFKH4FkbuopqX9PPWfuo= "Effect of pH value on saccharification and glucose production [LSD (p≤0.05): saccharification 0.031768, glucose 0.050895].")
In order to evaluate the optimal temperature for enzymatic saccharification of ATDPL, the hydrolysis was performed at 40–70°C (Fig. 5). The results indicate that the maximum enzymatic hydrolysis occurred at 50°C, corresponding to a degree of saccharification of 71.23%. However, a minor decrease in saccharification to 66.48% was observed at 55°C. These results are consistent with those of Krishna and Chowdary,40 Akhtar et al.,35 Taniguchi et al.,48 Mahamud and Gomes,36 and Sharma et al.5 They observed that maximum adsorption of enzymes and the saccharification rate were achieved at the same temperature (50°C) and the active enzyme was bound more strongly than the non-active enzyme to cellulose. The results also indicated that increasing the hydrolysis temperature to 70°C decreased the saccharification percentage to 27.31%. This behavior was probably caused by an increase in enzyme deactivation at higher temperatures. Baig et al.13 reported 45°C as an optimal temperature for enzymatic saccharification of steam-treated leaves and pseudo-stem of banana, while Zhang et al.42 found 40°C as optimal temperature for saccharification of steam explosion pre-treated corn stover.
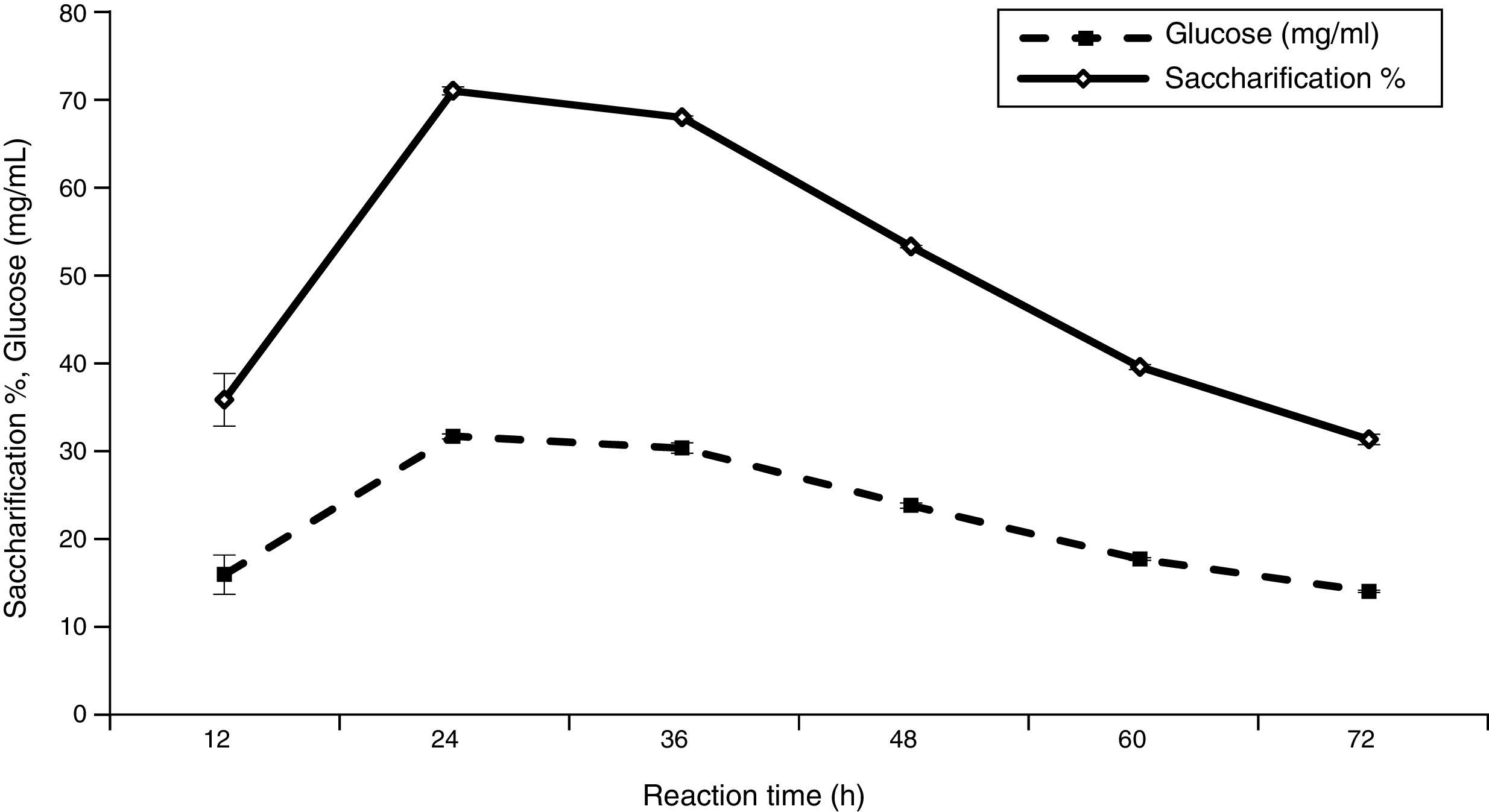
Effect of reaction time on saccharification and glucose production![Effect of temperature on saccharification and glucose production [LSD (p≤0.05): saccharification 0.036021, glucose 0.034173].](https://static.elsevier.es/multimedia/15178382/0000004700000001/v1_201602040959/S1517838215000167/v1_201602040959/en/main.assets/gr5.jpeg?xkr=ue/ImdikoIMrsJoerZ+w997EogCnBdOOD93cPFbanNerEcKXq/4gYMpNkXqSlVDY0lXCTapXrEOq5lCiG1bYbVRoz4b5rC/mtt5Z76dY4YaRNtQekK/QrNbPG6UsDyr4ZJ64xH79Pa0LNZ49YBb3E4vTKT/q/G8gNaEWafrN2gcQgUJMpSmzQR/S+VDYQeOXZiBdlYMrk+DESQxFCcCAU3L63HLMMXIWwsqgSnGzd7KylfPT7WP2H/0cJNWhxD0+tuqlh/JIhw715BOEieuMrCPbFKH4FkbuopqX9PPWfuo= "Effect of temperature on saccharification and glucose production [LSD (p≤0.05): saccharification 0.036021, glucose 0.034173].")
The optimum reaction period for sugar production and the saccharification percentage of ATDPL was determined by carrying out the hydrolysis in a shaker at 50°C for various time intervals (up to 72h) using 4% substrate and 30 FPU/g substrate in acetate buffer (pH 5). The sugar production and enzymatic hydrolysis in relation to the reaction time varied and showed the maximum value after 24h (Fig. 6). Extending the reaction time to 36h had no effect on saccharification. The same trend was also observed by Krishna and Chowdary40 and Baig et al.13 They reported that the optimum incubation period was 24h and extending the reaction time did not show any significant effect on enzymatic saccharification of alkaline pre-treated Antigonun leptopus leaves and pseudo-stems of banana. In contrast, the maximum saccharification yields (90% and 88.4%, respectively) were obtained at a 48-hour reaction time for saccharification of corncob residue and steam explosion pre-treated corn stover.42,43 Additionally, maximum enzymatic saccharification of alkaline pre-treated wheat straw, rice straw, and bagasse occurred after 20h in the presence of Bacillus subtilis cellulases.35 In another study, the saccharification percentage increased in a uniform manner up to 24h, but the saccharification per unit of time decreased.49 This phenomenon might occur by cellulose recrystallization caused at pH and temperature conditions used for hydrolysis and selective enzyme attack on the amorphous regions of cellulose; both of these processes might occur simultaneously during enzymatic hydrolysis. Furthermore, Mahmud and Gomes36 reported that the rate and extent of saccharification depend on pre-treatment of the substrate, enzyme and substrate concentration, product inhibition, and enzyme stability, all of these make the rate of hydrolysis to decrease rapidly with the time.
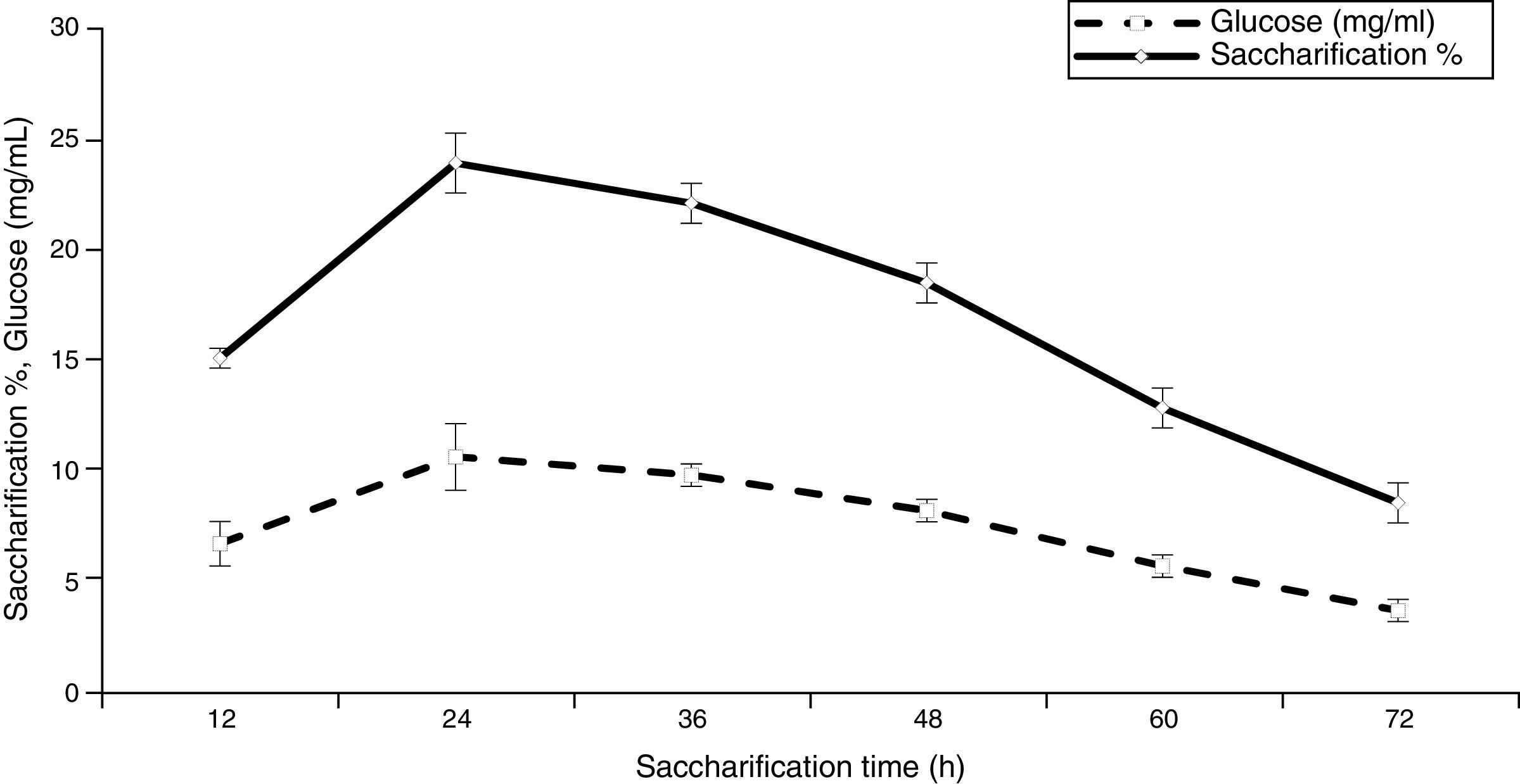
Two-step enzymatic saccharification process of ATDPL![Effect of reaction time on saccharification and glucose production [LSD (p≤0.05): saccharification 0.147473, glucose 0.112541].](https://static.elsevier.es/multimedia/15178382/0000004700000001/v1_201602040959/S1517838215000167/v1_201602040959/en/main.assets/gr6.jpeg?xkr=ue/ImdikoIMrsJoerZ+w997EogCnBdOOD93cPFbanNerEcKXq/4gYMpNkXqSlVDY0lXCTapXrEOq5lCiG1bYbVRoz4b5rC/mtt5Z76dY4YaRNtQekK/QrNbPG6UsDyr4ZJ64xH79Pa0LNZ49YBb3E4vTKT/q/G8gNaEWafrN2gcQgUJMpSmzQR/S+VDYQeOXZiBdlYMrk+DESQxFCcCAU3L63HLMMXIWwsqgSnGzd7KylfPT7WP2H/0cJNWhxD0+tuqlh/JIhw715BOEieuMrCPbFKH4FkbuopqX9PPWfuo= "Effect of reaction time on saccharification and glucose production [LSD (p≤0.05): saccharification 0.147473, glucose 0.112541].")
It is essential to reuse the enzymes absorbed on the residuals in subsequent saccharification cycles to achieve an economically viable process. The enzyme molecules bound to the residuals were recovered after the first step of hydrolysis following the methods of Vallander and Eriksson50 and Ouyang et al.43 The earlier reports indicate that 71.03% saccharification was obtained in a reference experiment (step 1), where the hydrolysis was continued only for 24h without interruption. As illustrated in Fig. 7, the saccharification was increased significantly to 94.88% by removing the hydrolysate of 24-hour hydrolysis and then suspending the residual in a fresh buffer to continue the hydrolysis for another 24h (step 2). This means that a multi-step alternative hydrolysis was better than the continuous hydrolysis for 48h; and the second batch of hydrolysis resulted in a total saccharification (step 2) of about 23.85%. The high saccharification rate obtained from the two-step process compared with the continuous saccharification for 48h may be explained on the basis of avoidance of the feedback inhibition of the enzymes by removing the oligosaccharides that accumulate in the first step process.43,51,52 Furthermore, Yang et al.53 investigated the feasibility of three-step hydrolysis of steam-exploded corn stover at high-substrate concentrations. They reported that, compared with the one-step hydrolysis for 72h, an increase in hydrolysis yield of 34–37% could be achieved. They also observed that the removal of end products improved the adsorption of cellulase on the substrate and enhanced the productivity during the enzymatic hydrolysis.
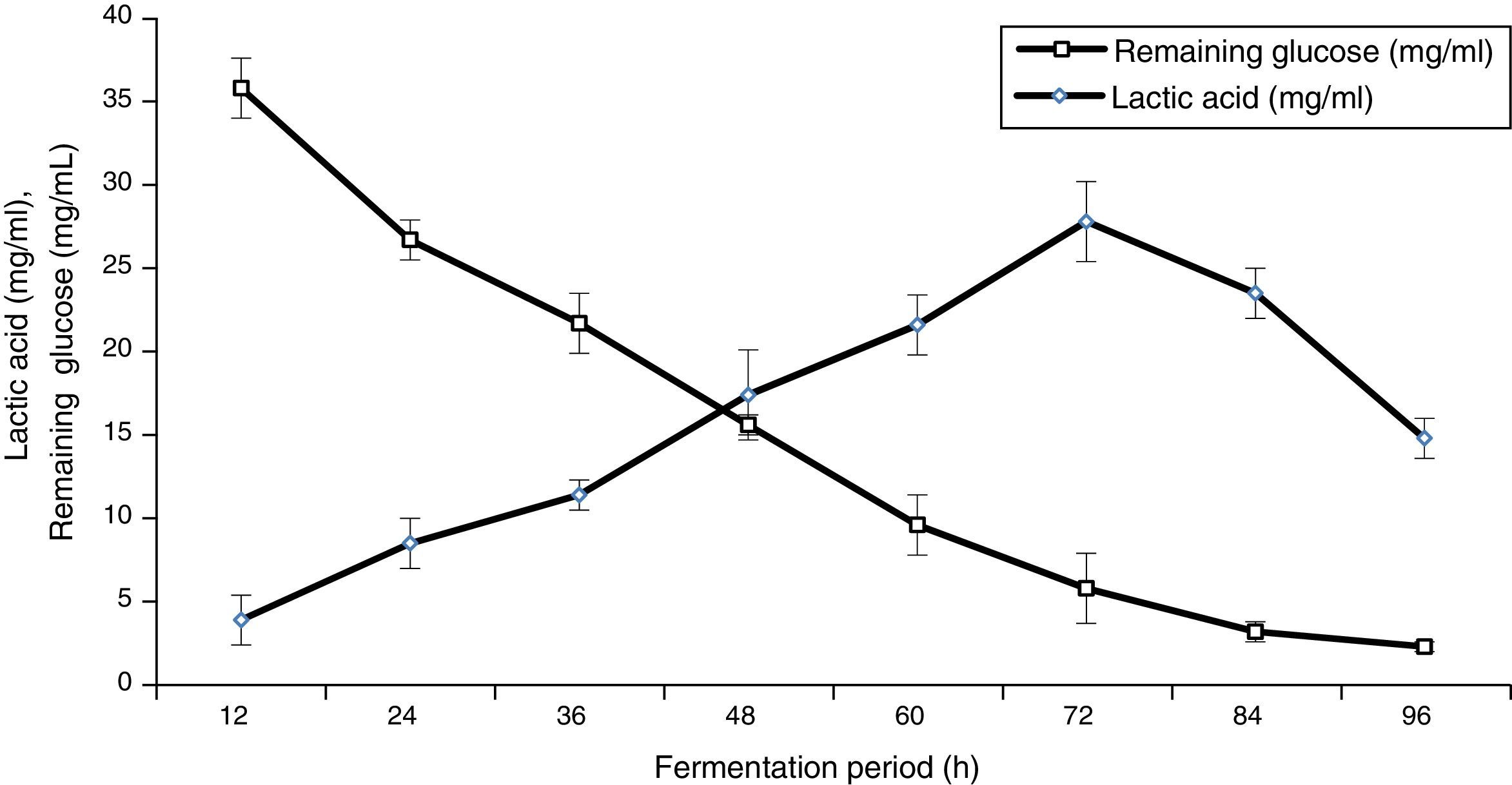
Lactic acid production by separate saccharification and fermentation technique![Enzymatic saccharification in the second hydrolysis process (two-step) [LSD (p≤0.05): saccharification 0.036221, glucose 0.058577].](https://static.elsevier.es/multimedia/15178382/0000004700000001/v1_201602040959/S1517838215000167/v1_201602040959/en/main.assets/gr7.jpeg?xkr=ue/ImdikoIMrsJoerZ+w997EogCnBdOOD93cPFbanNerEcKXq/4gYMpNkXqSlVDY0lXCTapXrEOq5lCiG1bYbVRoz4b5rC/mtt5Z76dY4YaRNtQekK/QrNbPG6UsDyr4ZJ64xH79Pa0LNZ49YBb3E4vTKT/q/G8gNaEWafrN2gcQgUJMpSmzQR/S+VDYQeOXZiBdlYMrk+DESQxFCcCAU3L63HLMMXIWwsqgSnGzd7KylfPT7WP2H/0cJNWhxD0+tuqlh/JIhw715BOEieuMrCPbFKH4FkbuopqX9PPWfuo= "Enzymatic saccharification in the second hydrolysis process (two-step) [LSD (p≤0.05): saccharification 0.036221, glucose 0.058577].")
As mentioned above, alkaline date palm leaves hydrolysate contained a high concentration of glucose (42.16mg/mL); therefore, this hydrolysate offers a great opportunity as feedstock for lactic acid production. Lactobacillus delbrueckii was cultivated at pH 5 and 50°C on the hydrolysate of alkaline date palm leaves as a substrate and then it was supplemented with other nutrients. The remaining glucose and lactic acid production throughout 96h of the incubation are illustrated in Fig. 8. Lactic acid production was obtained (27.8mg/mL) after 72h of incubation. The yield of lactic acid from glucose was 76.45% with 0.386g/L/h productivity rate. The remaining glucose was decreased when the increasing lactic acid production reached 5.8mg/mL after 3 days of incubation. This yield was higher compared to that reported by Parajo et al.54 They reported that 25.29mg/mL lactic acid was produced using processed wood hydrolysate; however, this yield was lower compared with the results reported by Shen and Xia.55 They reported a yield of 65mg/mL of lactic acid produced from municipal waste hydrolysate and 48.7g/L lactic acid from corn cob residue hydrolysate. Furthermore, Akao et al.17 and Zhou et al.19 observed increased lactic acid production when used separate saccharification and fermentation techniques. On the other hand, Shen and Xia55 reported that the accumulation of cellobiose and glucose in separate saccharification and fermentation processes would cause severe feedback inhibition of the cellulase reaction. Thus, the concentration of sugar in the hydrolysate was limited, and the yield of lactic acid was accordingly restricted.
Conclusions![Lactic acid production by separate saccharification and fermentation technique [LSD (p≤0.05): lactic acid 2.062469, remaining glucose 1.659854].](https://static.elsevier.es/multimedia/15178382/0000004700000001/v1_201602040959/S1517838215000167/v1_201602040959/en/main.assets/gr8.jpeg?xkr=ue/ImdikoIMrsJoerZ+w997EogCnBdOOD93cPFbanNerEcKXq/4gYMpNkXqSlVDY0lXCTapXrEOq5lCiG1bYbVRoz4b5rC/mtt5Z76dY4YaRNtQekK/QrNbPG6UsDyr4ZJ64xH79Pa0LNZ49YBb3E4vTKT/q/G8gNaEWafrN2gcQgUJMpSmzQR/S+VDYQeOXZiBdlYMrk+DESQxFCcCAU3L63HLMMXIWwsqgSnGzd7KylfPT7WP2H/0cJNWhxD0+tuqlh/JIhw715BOEieuMrCPbFKH4FkbuopqX9PPWfuo= "Lactic acid production by separate saccharification and fermentation technique [LSD (p≤0.05): lactic acid 2.062469, remaining glucose 1.659854].")
The results revealed that alkaline pre-treatment was the most effective digestion method to decrease the lignin content in all of the investigated wastes. The maximum sugar production and enzymatic saccharification levels were obtained using alkaline-treated date palm leaves using a multi-step hydrolysis by removing the hydrolysate after 24h. This study demonstrates that date palm wastes can be almost completely converted into fermentable sugar that can ultimately be converted into lactic acid by employing alkaline pre-treatment and separate saccharification and fermentation.
Conflict of interestThe author declares no conflicts of interest.
The author would like to acknowledge and thank the King Khalid University for supporting this study.