


The aim of this study was obtain a model that maximizes growth and production of inulinase and invertase by Aspergillus niger ATCC 20611, employing response surface methodology (RSM). The RSM with a five-variable and three-level central composite design (CCD) was employed to optimize the medium composition. Results showed that the experimental data could be appropriately fitted into a second-order polynomial model with a coefficient of determination (R2) more than 0.90 for all responses. This model adequately explained the data variation and represented the actual relationships between the parameters and responses. The pH and temperature value of the cultivation medium were the most significant variables and the effects of inoculum size and agitation speed were slightly lower. The intra-extracellular inulinase, invertase production and biomass content increased 10–32 fold in the optimized medium condition (pH 6.5, temperature 30°C, 6% (v/v), inoculum size and 150rpm agitation speed) by RSM compared with medium optimized through the one-factor-at-a-time method. The process development and intensification for simultaneous production of intra-extracellular inulinase (exo and endo inulinase) and invertase from A. niger could be used for industrial applications.
Microbial enzymes can be roughly classified into three major fields of application: (1) those which can be used to synthesize useful compounds; (2) that can stereo specifically carry out important bioconversion reactions; and (3) those which are able to hydrolyse polymers into interesting monomers.1 Many hydrolytic enzymes such as lipase, protease, and phytase have been commercially produced by yeasts. Inulinase (EC 3.2.1.7) and invertase (EC 3.2.1.26) both catalyze the hydrolysis of inulin and sucrose, but the inulinase enzyme has a higher specificity for inulin than invertase. These enzymes were initially isolated from plants, and thus it is difficult to extract high yields from such enzymes.2 Unlike plant enzymes, microbial enzymes show high inulin hydrolyzing activity.3 Microbial inulinase and invertase have great potential in a wide range of industrial and pharmaceutical applications. Study of structural functional properties of inulinases has large theoretical and applied significance in the conditions of various microenvirons, such as indicated growth and development of the organisms, play one of the key roles in controlling processes of cell differentiation, participate in carbohydrate metabolism of higher plants and microorganisms, present as most important components of signal pathways of the communication system, can be used in cycles of production of sugars with different degree of polymerization, in particular fructose and inulooligosaccharides – inalienable components of functional nutrition, lowering the risk of emergence of diabetes mellitus, caries and obesity.4 Invertase has a role in the metabolism, sucrose storage, as catalytic in the conversion of sucrose into glucose and fructose, produces invert syrup, which contains glucose and fructose at equimolar concentrations. The invert syrup is used in food and beverage industries as a humectant in the preparation of candies, noncrystallizing creams, jams and artificial honey.5,6 It is reported in the present study A. niger simultaneously produces intra-extracellular inulinase and invertase, which offers a great advantage to industry, particularly the food industry, for production of ultra-high fructose syrup, bio-ethanol, inulo-oligosaccharide, single-cell oil, single-cell protein, citric acid, butanediol, alcohols and lactic acid from inulin and sucrose by a single-step process.7–10 Since many enzymes of industrial significance are regulated by the medium composition and fermentation conditions, a study of such regulation is of prime importance for the commercial production of such enzymes.
Optimization of the fermentation conditions to achieve a suitable balance is very important to obtain the optimum microbial growth and high enzyme yield during fermentation. Traditional methods of optimization involve changing one independent variable, while fixing the others at given levels. This single-dimensional search technique is simple, but often fails to yield optimized conditions because it does not consider possible interactions among factors. Response surface method (RSM) is an efficient experimental strategy to run optimal conditions for multivariable systems. It is a collection of statistical techniques for design of experiments, building models, measured responses, evaluating the effects and relationship between clusters of controlled experimental factors and searching for the optimum conditions, has successfully been used in the optimization of bioprocesses.11 The benefit of employing RSM is to decrease the number of experimental trials that are needed to assess numerous parameters and their interactions.12 Also, the method of RSM makes it possible to study several factors simultaneously, to quantify the individual effect of each factor and to investigate their possible interactions.13,14 RSM can therefore be far less laborious and time-consuming compared to other methods for running an optimization process. Other advantages of using RSM is that no complex calculations are required to analyze the resulting data; in addition, searching for relativity between factors is possible, and the most suitable conditions can be found, and responses can be forecast.12
The objective of this study was to optimize fermentation conditions, using RSM to select an optimal range of physical parameters for maximum production of intra-extracellular inulinase and invertase as well as A. niger growth. First, the effect of medium composition on intra- and extracellular inulinase (exo- and endo), invertase production and biomass were evaluated by ANOVA. Then, based on the results of the optimization with one independent variable, RSM was applied in this study to optimize the factors of medium for enhancement of intra-extracellular inulinase, invertase production and growth. It is hypothesized that optimizing intra-extracellular inulinase and invertase production open new horizons for a wide range of industrial applications.
Materials and methodsMicroorganism and media compositionA. niger ATCC 20611, a high producer of intra-extracellular inulinase and invertase, was purchased from the American Type Culture Collection (ATCC; Rockville, Maryland, USA). The strain was maintained at 4°C on potato dextrose agar (PDA) after being incubated for 4 days at 30°C and sub-cultured every 3 weeks. The spores were harvested and suspended in sterile distilled water containing 0.01% (v/v) Tween 80 to obtain approximately 2.0×106spores/mL. Preliminary experiments were performed by using a production medium consisting of: sucrose 10% (w/v), yeast extract 2.5% (w/v), NaNO3 2% (w/v), Zn+2 1.5mM (v/v), Triton X-100 1% (v/v).12 The initial pH of the basal medium was adjusted to 6.0 with HCl (5M) and NaOH (5M), prior to sterilization at 121°C for 15min. Erlenmeyer flasks (250mL) containing 50mL of the production medium were inoculated with 6% (v/v) stock culture and incubated at 30°C with 150rpm shaking for 96h.12 All the fermentation procedures were carried out in triplicate.
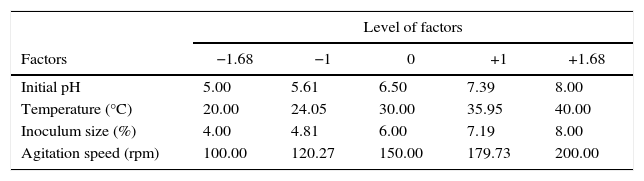
Experimental design and statistical analysisThe optimization of fermentation conditions is an important tool for the development of economically feasible bioprocesses. Combined interactions of medium physical parameters for the production of the desired product are large and the optimum process conditions may be developed using an effective experimental design procedure.12,15 RSM, which is a collection of statistical techniques for design of experiments, building models, measured responses, evaluating the effects and relationship between clusters of controlled experimental factors and searching for the optimum conditions, has successfully been used in the optimization of bioprocesses.15,16 A prior knowledge with understanding of the related bioprocesses is necessary for achieving a more realistic model. In the present work, four independent variables were studied at five different levels (Table 1) and screened in 21 experimental runs (Table 2), then insignificant ones were eliminated in order to obtain a smaller and more fitting set of factors. The minimum and maximum ranges of variables investigated in cultivation medium were pH (5–8), temperature (20–40°C); inoculum size (4–8%) and agitation speed (100–200rpm). The fractional factorial design comprised of 8 factorial points, 8 axial points and 5 center points. The center point is repeated five times to give a good estimate of the experimental error (pure error variance). This offered an adequate estimate of the variation of the response and provided the number of degrees of freedom needed for an adequate statistical test of the model.12
Composition of various experiments of the CCD for independent variables and responses.
| Run no. | Initial pH | Temperature (°C) | Inoculum size (%) | Agitation speed (rpm) | Extracellular inulinase (U/mL) | Intracellular inulinase (U/mL) | Extracellular invertase (U/mL) | Intracellular invertase (U/mL) | Biomass (mg/mL) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Actual | Predicted | Actual | Predicted | Actual | Predicted | Actual | Predicted | Actual | Predicted | |||||
| 1 | 7.39 | 35.95 | 7.19 | 120.27 | 2365.54 | 2028.32 | 1374.64 | 1051.09 | 2645.90 | 2355.74 | 1338.66 | 1115.07 | 34.47 | 30.32 |
| 2 | 7.39 | 35.95 | 4.81 | 120.27 | 2157.87 | 1840.58 | 1252.54 | 1001.40 | 2047.95 | 1729.27 | 1173.22 | 1071.55 | 32.66 | 29.04 |
| 3 | 7.39 | 24.05 | 7.19 | 179.73 | 1948.70 | 1908.24 | 1009.34 | 872.56 | 1075.54 | 623.35 | 1095.64 | 908.85 | 30.76 | 26.95 |
| 4 | 5.61 | 35.95 | 4.81 | 179.73 | 139.87 | −177.99 | 57.54 | −237.29 | 1584.50 | 1417.55 | 185.32 | 7.66 | 25.65 | 22.00 |
| 5 | 7.39 | 24.05 | 4.81 | 179.73 | 2076.80 | 1881.29 | 927.91 | 840.99 | 153.54 | −3.12 | 1003.81 | 865.33 | 29.65 | 26.24 |
| 6 | 5.61 | 24.05 | 7.19 | 120.27 | 136.50 | 96.62 | 36.70 | −56.39 | 1089.53 | 789.08 | 180.26 | 37.62 | 23.30 | 19.39 |
| 7 | 5.61 | 35.95 | 7.19 | 179.73 | 185.60 | −151.04 | 74.14 | −205.72 | 1584.54 | 1301.46 | 198.77 | 51.18 | 26.84 | 23.27 |
| 8 | 5.61 | 24.05 | 4.81 | 120.27 | 104.96 | −91.13 | 24.54 | −106.08 | 1054.75 | 905.17 | 176.71 | −5.91 | 22.56 | 18.69 |
| 9 | 5.00 | 30.00 | 6.00 | 150.00 | 76.50 | 391.33 | 20.05 | 302.33 | 100.65 | 380.58 | 52.42 | 282.41 | 21.25 | 26.55 |
| 10 | 8.00 | 30.00 | 6.00 | 150.00 | 41.56 | 356.39 | 13.03 | 295.30 | 34.54 | 503.34 | 14.18 | 244.18 | 10.56 | 15.86 |
| 11 | 6.50 | 20.00 | 6.00 | 150.00 | 1025.98 | 1091.94 | 493.54 | 567.12 | 1023.40 | 1397.77 | 513.80 | 743.79 | 24.62 | 29.92 |
| 12 | 6.50 | 40.00 | 6.00 | 150.00 | 420.97 | 984.66 | 100.70 | 591.67 | 543.54 | 917.91 | 260.65 | 490.64 | 20.36 | 25.66 |
| 13 | 6.50 | 30.00 | 4.00 | 150.00 | 1765.30 | 2161.16 | 1093.54 | 1355.08 | 2842.85 | 3058.45 | 861.10 | 1061.31 | 40.12 | 45.16 |
| 14 | 6.50 | 30.00 | 8.00 | 150.00 | 2107.90 | 2341.70 | 1120.40 | 1423.42 | 2954.50 | 3487.63 | 874.73 | 1134.51 | 41.25 | 46.82 |
| 15 | 6.50 | 30.00 | 6.00 | 100.00 | 2398.70 | 2713.53 | 1287.73 | 1570.01 | 3043.90 | 3418.27 | 952.21 | 1182.20 | 50.50 | 56.13 |
| 16 | 6.50 | 30.00 | 6.00 | 200.00 | 2485.80 | 2800.63 | 1357.38 | 1639.66 | 3059.43 | 3433.80 | 1672.38 | 1902.37 | 51.58 | 56.55 |
| 17 | 6.50 | 30.00 | 6.00 | 150.00 | 3949.54 | 3266.89 | 2801.80 | 2351.42 | 4954.89 | 4331.77 | 2193.03 | 1782.51 | 77.72 | 70.05 |
| 18 | 6.50 | 30.00 | 6.00 | 150.00 | 3476.50 | 3266.89 | 2284.30 | 2351.42 | 4534.54 | 4331.77 | 1995.65 | 1782.51 | 67.91 | 70.05 |
| 19 | 6.50 | 30.00 | 6.00 | 150.00 | 3346.87 | 3266.89 | 2484.40 | 2351.42 | 4416.54 | 4331.77 | 1886.42 | 1782.51 | 74.68 | 70.05 |
| 20 | 6.50 | 30.00 | 6.00 | 150.00 | 3275.56 | 3266.89 | 2688.63 | 2351.42 | 4395.54 | 4331.77 | 1752.25 | 1782.51 | 69.40 | 70.05 |
| 21 | 6.50 | 30.00 | 6.00 | 150.00 | 3023.65 | 3266.89 | 2159.40 | 2351.42 | 4234.54 | 4331.77 | 1624.14 | 1782.51 | 72.95 | 70.05 |
Once the critical factors was identified through the screening, the central composite design (CCD) was used to obtain a quadratic model, consisting of factorial trials and star points to estimate quadratic effects and central points to estimate the pure process variability with biomass, intra-extracellular inulinase and invertase production as the responses (Y). The central composite design experimental data were employed using Design Expert version 6.06 (Stat-Ease Inc., Minneapolis, USA) and then interpreted. Behavior of the system is explained by the following second-order polynomial equation:
where Y is the dependent variable, xi and xj are the independent variables, β0, βi, βii and βij are the regression coefficients of model and e is the error of model. In order to test the estimation capabilities of the technique, the predicted responses obtained from RSM were compared with the actual responses. An analysis of variance (ANOVA) and R2 statistic were performed to evaluate significant differences between factors and check the adequacy of the developed model and must be close to 1. Lack-of-fit is a special diagnostic test for adequacy of a model that compares the pure error, based on the replicate measurements to the other lack of fit, based on the model performance.17F-value, calculated as the ratio between the lack-of-fit mean square and the pure error mean square, is the statistic parameter used to determine whether the lack of-fit is significant or not, at a significance level. The statistical model was validated with respect to intra-extracellular inulinase, invertase production and biomasses under the conditions predicted by the model in erlenmeyer flasks level. Samples were drawn at the desired intervals and intra-extracellular inulinase, invertase production and biomasses were determined as described above (data not shown).Analytical method for intra-extracellular inulinase, invertase activities and biomass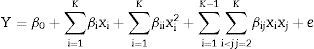
The supernatants of the cultures at 96h of fermentation were harvested by centrifugation at 4°C (20min, 10,000×g) in triplicate to determinate extracellular inulinase activity (I) and invertase activity (S). Then, the cell pellet was used as a source of intracellular inulinase and invertase activities enzymes. For the extraction of intracellular enzymes, the pellet was resuspended in 50mL of sodium acetate buffer (200mM, pH 5.0) with vigorous vortexing followed by incubation at 30°C and 150rpm for 30min. Then, it was sonicated on ice in glass tubes using a Branson Sonic Power Sonicator (48 Bransonic Power, 40W, 30s with 30s cooling periods) for 5min followed by centrifugation at 4°C (20min, 10,000 ×g).12 Cell debris was removed by centrifugation at 4°C (20min, 10,000×g). The supernatant was used as the crude intracellular inulinase and invertase enzymes. The inulinase and invertase activities were assayed by measuring the reducing sugar released from inulin and sucrose, respectively, with DNS method as describe by Dinarvand et al., 12. The same reaction mixture without the enzyme extract was used as the control. One unit (U) of inulinase and invertase activity was defined as the amount of enzyme (in 1mL), which liberates 1μmol of fructose or glucose equivalent from inulin (inulinase) or sucrose (invertase) per min, respectively. The pH was measured by pH-meter (620 lab pH meter). Biomass concentration was determined according to Dinarvand et al. 12, while the dry mass was expressed as milligram per milliliter of the fungal culture. Experiments were carried out in triplicate runs, and all standard deviations were lower than 10%.
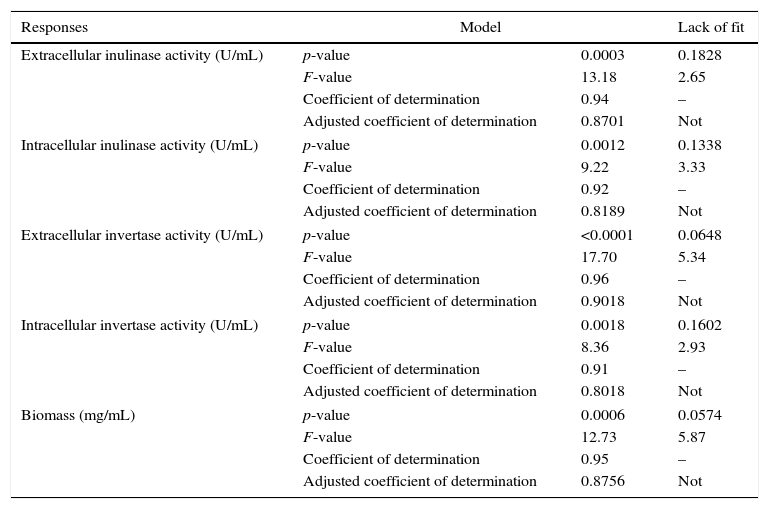
ResultsModel fitting and analysis of variance (ANOVA)Analysis of variance for response surface quadratic polynomial models of intra-extracellular inulinase, invertase production and biomass are shown the models are significant (0.5 <) but the lack of fits are not significant (0.5 >). The computed model F-value of intracellular inulinase (9.22), extracellular inulinase (13.18), intracellular invertase (8.36), extracellular invertase (17.70) production and biomass (12.73) implies that the model is significant and there is only a 0.01% chance that a “model F-value” this large could occur due to noise. In addition, with a suitable coefficient of determination and adjusted coefficient of determination (Table 3), the analysis of ANOVA shown the lowest predicted residual sum of squares for “model Prob> F” (<0.0001) and P-values (<0.0018).
Analysis of variance (ANOVA).
| Responses | Model | Lack of fit | |
|---|---|---|---|
| Extracellular inulinase activity (U/mL) | p-value | 0.0003 | 0.1828 |
| F-value | 13.18 | 2.65 | |
| Coefficient of determination | 0.94 | – | |
| Adjusted coefficient of determination | 0.8701 | Not | |
| Intracellular inulinase activity (U/mL) | p-value | 0.0012 | 0.1338 |
| F-value | 9.22 | 3.33 | |
| Coefficient of determination | 0.92 | – | |
| Adjusted coefficient of determination | 0.8189 | Not | |
| Extracellular invertase activity (U/mL) | p-value | <0.0001 | 0.0648 |
| F-value | 17.70 | 5.34 | |
| Coefficient of determination | 0.96 | – | |
| Adjusted coefficient of determination | 0.9018 | Not | |
| Intracellular invertase activity (U/mL) | p-value | 0.0018 | 0.1602 |
| F-value | 8.36 | 2.93 | |
| Coefficient of determination | 0.91 | – | |
| Adjusted coefficient of determination | 0.8018 | Not | |
| Biomass (mg/mL) | p-value | 0.0006 | 0.0574 |
| F-value | 12.73 | 5.87 | |
| Coefficient of determination | 0.95 | – | |
| Adjusted coefficient of determination | 0.8756 | Not | |
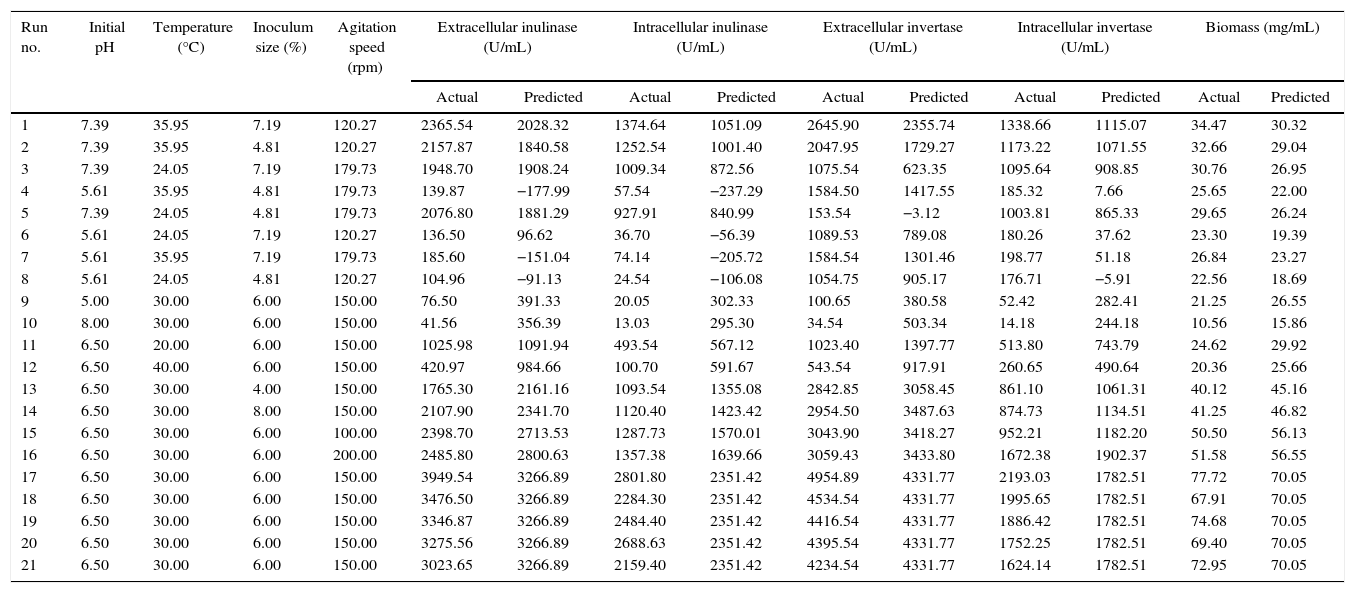
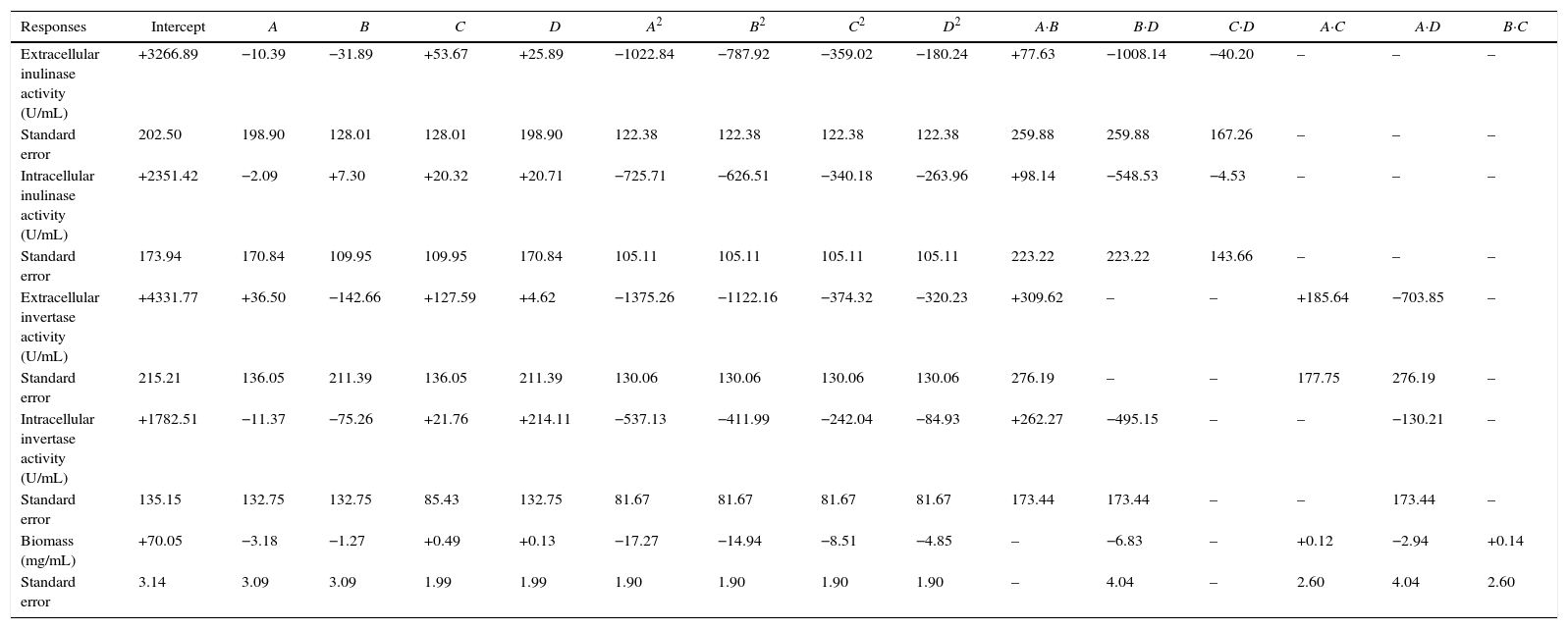
ANOVA analysis of intra-extracellular inulinase, invertase production and biomass (mg/mL) revealed that the first order of each parameter has not significant effect on responses (p>0.05). The second order of pH, temperature and inoculum size showed significant effect on intra-extracellular inulinase, invertase production and biomass (p<0.05) while agitation speed was not significant for all responses (data not shown). Among the different interactions, only interactions between temperature–agitation speed and initial pH–agitation speed significantly (p<0.05) affected the responses (intra-extracellular inulinase, invertase productions and biomass). The interaction between initial pH-temperature did not show significant effects on responses (p>0.05); however, their interaction was also kept in the final model to have the model less modified. The predicted and observed responses along with design matrix are presented in Table 2. According to ANOVA, it can be deduced that the quadratic polynomial model was significant and adequate to represent the actual relationship between the response variables (intra-extracellular inulinase, invertase production and biomass) and the significant variables. The second-order regression equation provided the levels of intra-extracellular inulinase, invertase production and biomass as the function of initial pH, temperature, inoculum size and agitation speed which can be presented in terms of coded factors as in the following equation as shown in Table 4.
The equation of the parameters (intra-extracellular inulinase, invertase production and biomass) as the function of initial pH, temperature, inoculum size and agitation speed which can be presented in terms of coded factors.
| Responses | Intercept | A | B | C | D | A2 | B2 | C2 | D2 | A·B | B·D | C·D | A·C | A·D | B·C |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Extracellular inulinase activity (U/mL) | +3266.89 | −10.39 | −31.89 | +53.67 | +25.89 | −1022.84 | −787.92 | −359.02 | −180.24 | +77.63 | −1008.14 | −40.20 | – | – | – |
| Standard error | 202.50 | 198.90 | 128.01 | 128.01 | 198.90 | 122.38 | 122.38 | 122.38 | 122.38 | 259.88 | 259.88 | 167.26 | – | – | – |
| Intracellular inulinase activity (U/mL) | +2351.42 | −2.09 | +7.30 | +20.32 | +20.71 | −725.71 | −626.51 | −340.18 | −263.96 | +98.14 | −548.53 | −4.53 | – | – | – |
| Standard error | 173.94 | 170.84 | 109.95 | 109.95 | 170.84 | 105.11 | 105.11 | 105.11 | 105.11 | 223.22 | 223.22 | 143.66 | – | – | – |
| Extracellular invertase activity (U/mL) | +4331.77 | +36.50 | −142.66 | +127.59 | +4.62 | −1375.26 | −1122.16 | −374.32 | −320.23 | +309.62 | – | – | +185.64 | −703.85 | – |
| Standard error | 215.21 | 136.05 | 211.39 | 136.05 | 211.39 | 130.06 | 130.06 | 130.06 | 130.06 | 276.19 | – | – | 177.75 | 276.19 | – |
| Intracellular invertase activity (U/mL) | +1782.51 | −11.37 | −75.26 | +21.76 | +214.11 | −537.13 | −411.99 | −242.04 | −84.93 | +262.27 | −495.15 | – | – | −130.21 | – |
| Standard error | 135.15 | 132.75 | 132.75 | 85.43 | 132.75 | 81.67 | 81.67 | 81.67 | 81.67 | 173.44 | 173.44 | – | – | 173.44 | – |
| Biomass (mg/mL) | +70.05 | −3.18 | −1.27 | +0.49 | +0.13 | −17.27 | −14.94 | −8.51 | −4.85 | – | −6.83 | – | +0.12 | −2.94 | +0.14 |
| Standard error | 3.14 | 3.09 | 3.09 | 1.99 | 1.99 | 1.90 | 1.90 | 1.90 | 1.90 | – | 4.04 | – | 2.60 | 4.04 | 2.60 |
A, initial pH; B, temperature; C, inoculum size; D, agitation speed.
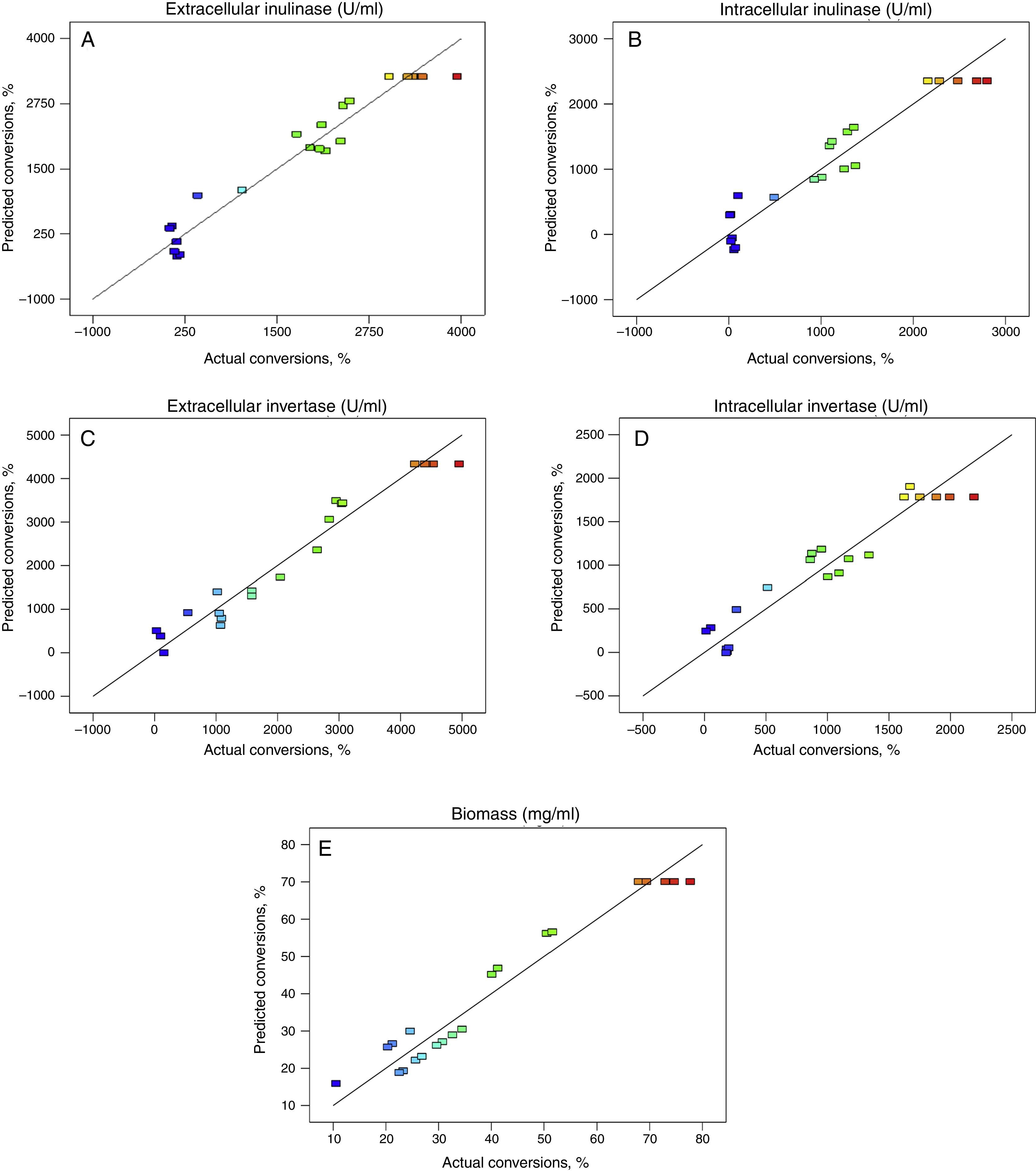
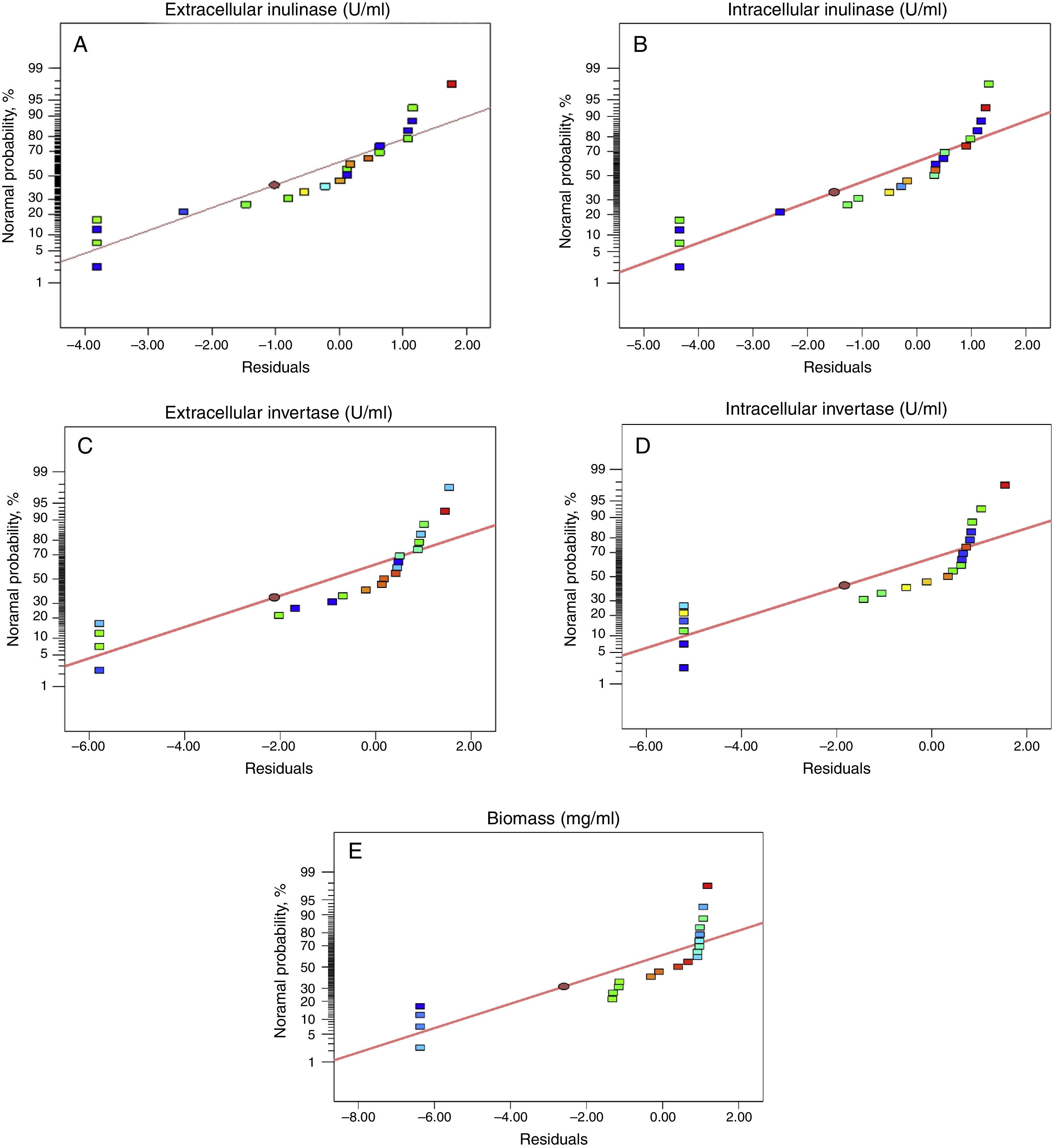
Fig. 1 shows that the actual response values agree well with the predicted response values. A linear distribution is observed which is indicative of a well-fitting model. Also, straight line in Fig. 2 approved the satisfaction of normality assumption. The computed ratio of 8.5, 9.7, 8.0, 11.5 and 9.4 for intra-extracellular inulinase, invertase production and biomass, respectively, in this study indicates an adequate signal.
, invertase (C and D) productions and biomass (E), respectively.")
, invertase (C and D) productions and biomass (E), respectively.")
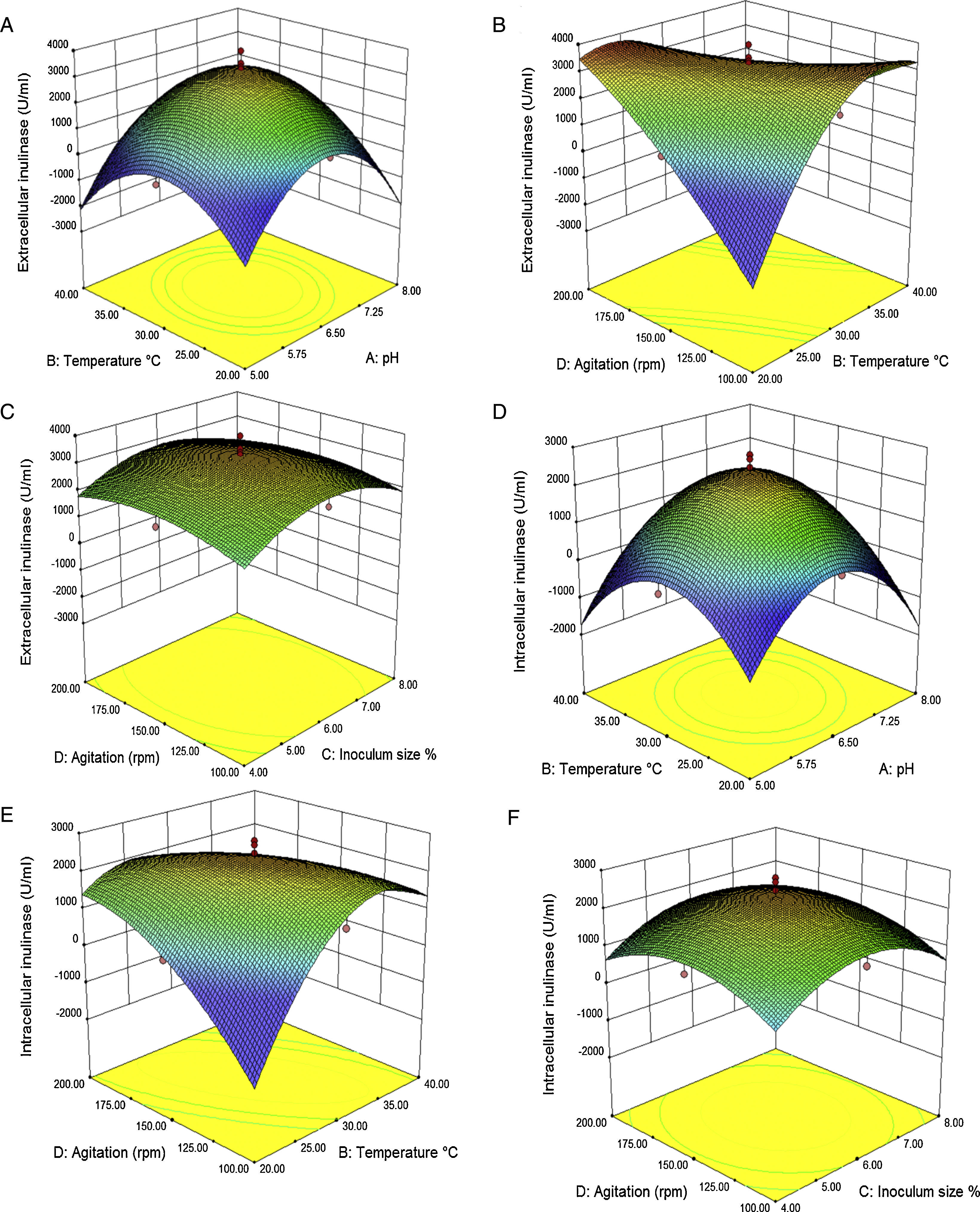
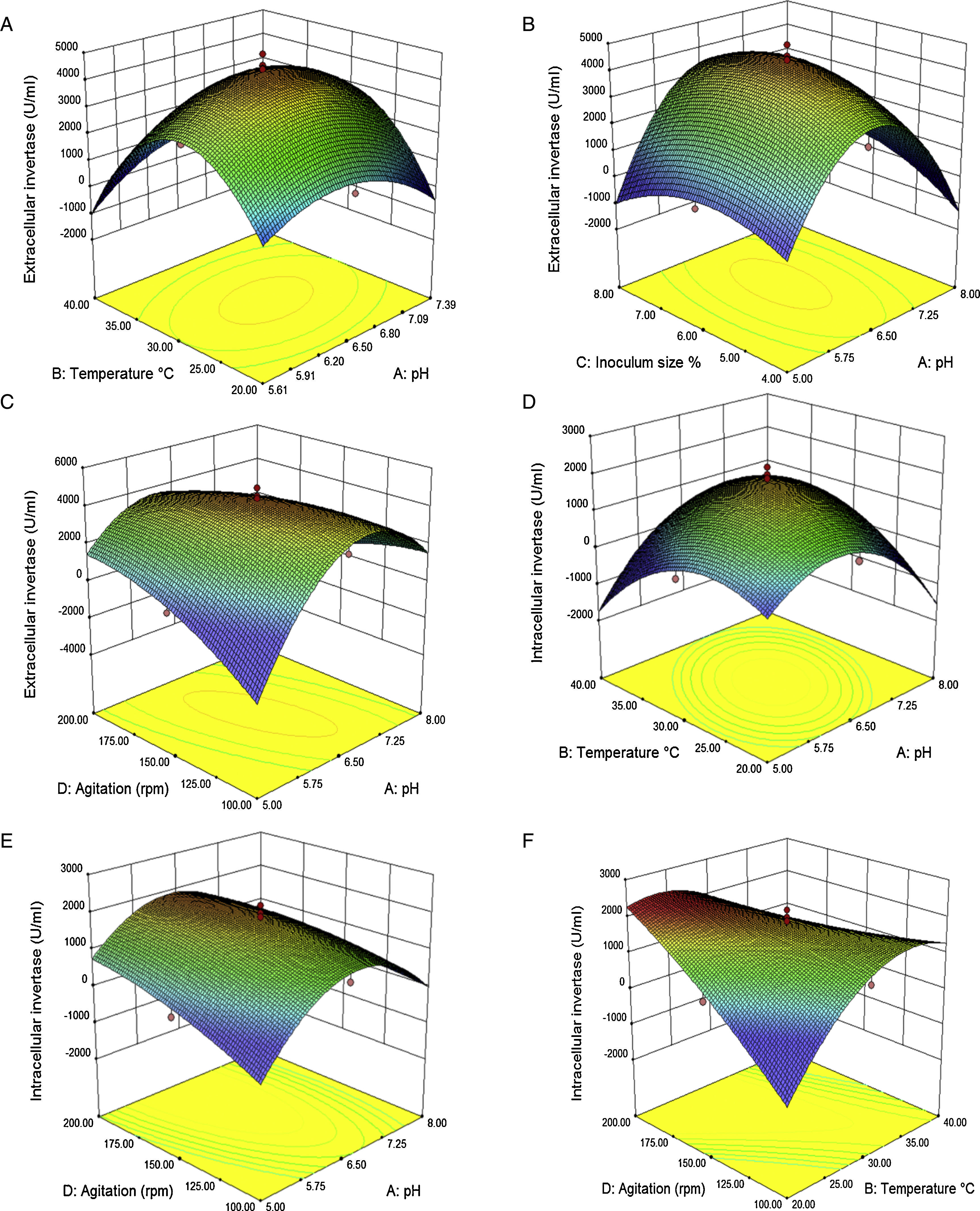
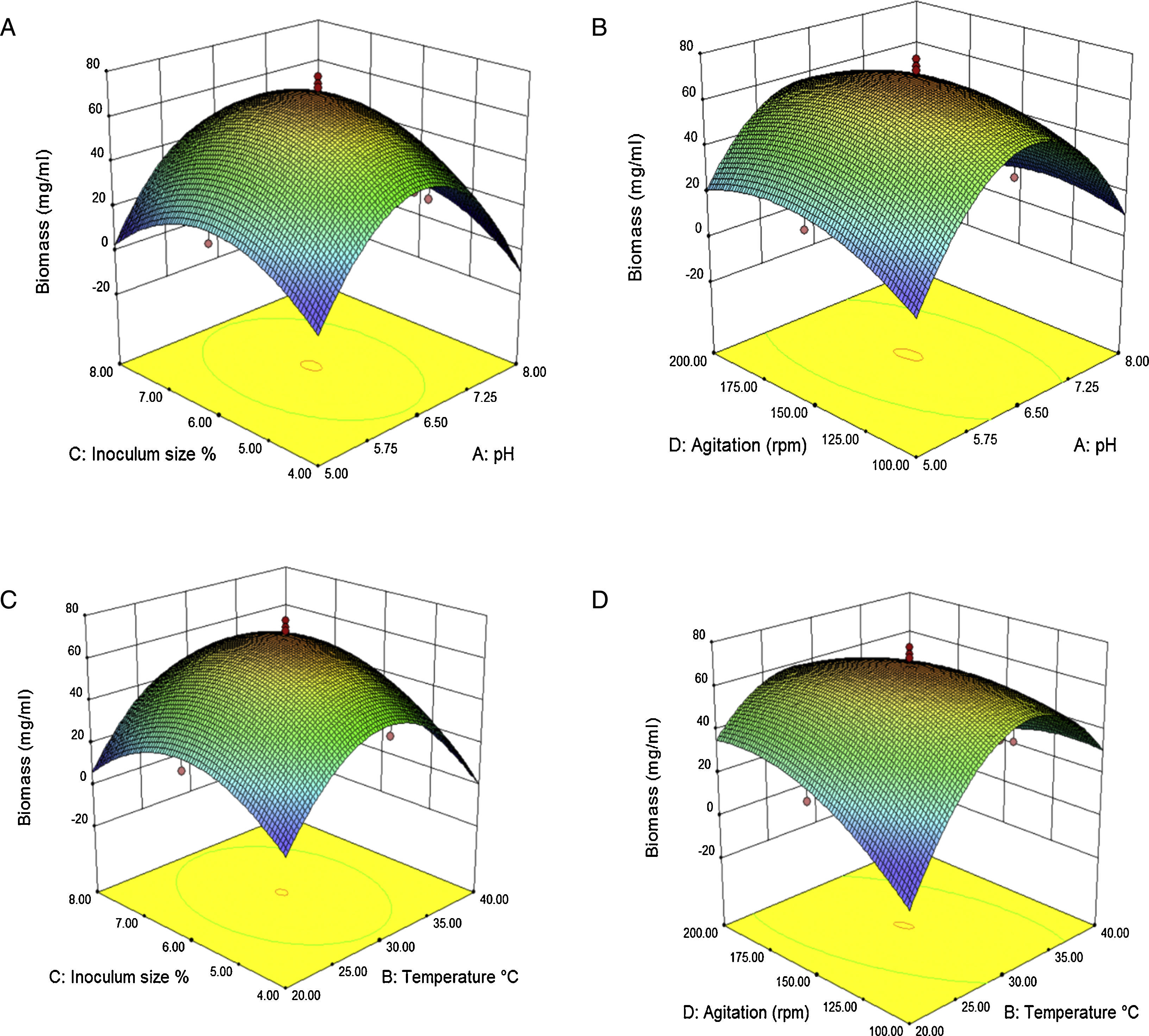
The interaction effects of variables on the calculated responses (intra-extracellular inulinase, invertase production and biomass) were studied by plotting 3D surface curves against any two independent variables, while keeping another variable at its central (0) level as shown in Figs. 3–5. The results from this study show that the actual response values agree well with the predicted response values in Table 2.
 productions from A. niger TCC 20611 in batch fermentation as a function of temperature (°C) and pH (A and D), temperature (°C) and agitation speed (rpm) (B and E), agitation speed (rpm) and inoculum size (C and F). Other variables are constant at their center points. The numbers inside the contour plots indicate conversion yield (U/mL) of the intra-extracellular inulinase productions.")
Response surface for intra-extracellular inulinase (U/mL) productions from A. niger TCC 20611 in batch fermentation as a function of temperature (°C) and pH (A and D), temperature (°C) and agitation speed (rpm) (B and E), agitation speed (rpm) and inoculum size (C and F). Other variables are constant at their center points. The numbers inside the contour plots indicate conversion yield (U/mL) of the intra-extracellular inulinase productions.
 productions. (A and D) temperature (°C) and pH, (B) inoculum size and pH, (C and E) agitation speed (rpm) and pH, (F) agitation speed (rpm) and temperature (°C). Other variables are constant at their center points. The numbers inside the contour plots indicate conversion yield (U/mL) of the intra-extracellular invertase productions.")
Response surface showing the interaction between four parameters and intra-extracellular invertase (U/mL) productions. (A and D) temperature (°C) and pH, (B) inoculum size and pH, (C and E) agitation speed (rpm) and pH, (F) agitation speed (rpm) and temperature (°C). Other variables are constant at their center points. The numbers inside the contour plots indicate conversion yield (U/mL) of the intra-extracellular invertase productions.
 pH, (B) temperature, (C) inoculum size and (D) agitation speed (rpm) on biomass.")
The combined effect of initial temperature–pH and inoculum size–pH on intra-extracellular inulinase, invertase production and biomass is shown in Figs. 3, 4(A and D) and 5(A), respectively. The maximum intra-extracellular inulinase productions occurred at pH (6.5) (Fig. 3A and D), further decrease or increase in the pH (6.5) led to a decrease in intra-extracellular inulinase production. According to the plots (Figs. 4A and D and 5A), intra-extracellular invertase productions and growth rate of A. niger occurs at moderately acidic initial pH of 6.5. The initial pH value lower or higher than 6.5 decreases the invertase production and growth rate of A. niger.
Effect of temperature on intra-extracellular inulinase, invertase production and biomassAs shown in Fig. 3(A and D) the combined effect of temperature–pH on intra-extracellular inulinase productions indicated that intracellular and extracellular inulinase productions were effectively increased from 30 to 35°C and decreased thereafter up to 40°C. The combined effect of temperature–pH and inoculum size-temperature on intra-extracellular invertase productions and biomass is also depicted in Fig. 4(A and D) and Fig. 5(C), respectively. Intracellular and extracellular invertase production and biomass were effectively increased when the temperature increased up to 30°C and decreased thereafter.
Effect of inoculum size on intra-extracellular inulinase, invertase production and biomassThe combined effect of agitation speed-inoculum size on intra-extracellular inulinase production (Fig. 3C and F) and inoculum size–pH on extracellular invertase production (Fig. 4B) and biomass (Fig. 5A) show that the highest production of enzymes and biomass were achieved at 6% (v/v) inoculum size. With higher or lower values, the intra-extracellular inulinase, extracellular invertase production and biomass were comparatively reduced. The same trend was observed for intracellular invertase production (data not shown).
Effect of agitation on intra-extracellular inulinase, invertase production and biomassFigs. 3(B and E), 4(C and E) and 5(B and D) represent the three dimensional plots as a function of combined effect of agitation speed–temperature and agitation speed–pH, on intra-extracellular inulinase, invertase productions and biomass, respectively, when other parameters were kept at an optimum point. Maximum extracellular inulinase and invertase production was obtained at the 150rpm that was technically similar to the condition for maximum intracellular inulinase, invertase production and biomass.
OptimizationThe optimal fermentation conditions for maximum production of intra and extracellular inulinase (2801U/mL and 3949U/mL), intra and extracellular invertase (2193U/mL, 4954U/m) and biomass (77.7mg/mg) was achieved by the combined level of 6% (v/v) of spore's suspension, 150rpm agitation speed, pH 6.5 and temperature 30°C.
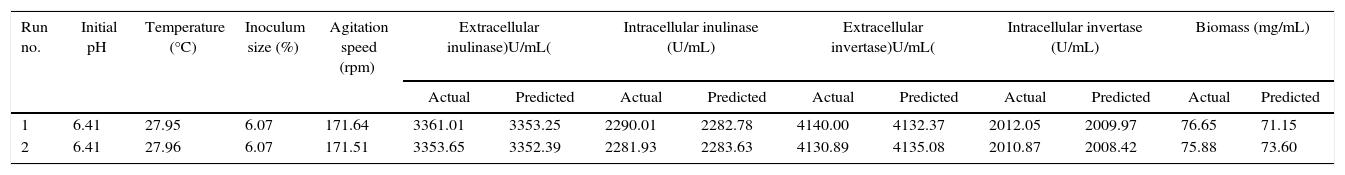
Model validationTo test the competence of the response surface equations, the experimental and predicted values were compared for measurement the intra-extracellular inulinase, invertase production and biomass from A. niger under optimal operation conditions. Repeated experiments were performed and the results were compared. The actual and predicted intra-extracellular inulinase, invertase production and biomass rate can be seen from values listed in Table 5. The theoretical values for levels of intra-extracellular inulinase, invertase production and biomass were 2282, 3353, 2009, 4132U/mL and 71.2mg/mL respectively, while the actual values were 2290.01, 3361.01, 2012.05, 4140U/mL and 76.7mg/mL, respectively after 4 days of culturing.
Effect of optimum conditions of parameters on responses.
| Run no. | Initial pH | Temperature (°C) | Inoculum size (%) | Agitation speed (rpm) | Extracellular inulinase)U/mL( | Intracellular inulinase (U/mL) | Extracellular invertase)U/mL( | Intracellular invertase (U/mL) | Biomass (mg/mL) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Actual | Predicted | Actual | Predicted | Actual | Predicted | Actual | Predicted | Actual | Predicted | |||||
| 1 | 6.41 | 27.95 | 6.07 | 171.64 | 3361.01 | 3353.25 | 2290.01 | 2282.78 | 4140.00 | 4132.37 | 2012.05 | 2009.97 | 76.65 | 71.15 |
| 2 | 6.41 | 27.96 | 6.07 | 171.51 | 3353.65 | 3352.39 | 2281.93 | 2283.63 | 4130.89 | 4135.08 | 2010.87 | 2008.42 | 75.88 | 73.60 |
X2=∑(ya−yp)2yp=0.56.
The predicted values from the regression equation closely agreed with that obtained from experimental values. The intra-extracellular inulinase, invertase and biomass obtained from experiments was very close to the actual response predicted by the regression model, which proved the validity of the model. In addition, some physical factors like pH and temperature can act as inducers, for biomass and production of intra-extracellular hydrolytic enzymes. The optimum conditions can be used for future upscale productions of the intra-extracellular inulinase, invertase and biomass. The maximum enzymes production and growth rate were obtained in a short time (96h).12 From an economic stand-point, it would be favorable to use the minimum time and maximum enzyme production.
The X2 goodness-of-fit test was used to examine the validity of the model by summing the squared difference between the actual (A) and the predicted (P) values, divided by the expected values in all possible categories as shown in the following equation.18,19
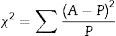
The test shows that there is not a significant difference between the predicted and actual values since the X2 value for production of extracellular inulinase (0.003), intracellular inulinase (0.009), extracellular invertase (0.008), intracellular invertase (1.72) and biomass (0.81) are much smaller than the cut-off value of X0.52 for 1 degrees of freedom (3.84). This indicates that the generated model is valid at 95% confidence level. Prediction of intra-extracellular inulinase, invertase production and biomass at any tested parameter within the range of experimental design is achieved by employing second-order polynomial regression equation obtained from experimental data. Point prediction capability of the software with predicted intra-extracellular inulinase, invertase production and biomass rate lead to the idea of confirmation of the conditions located within the levels defined previously.
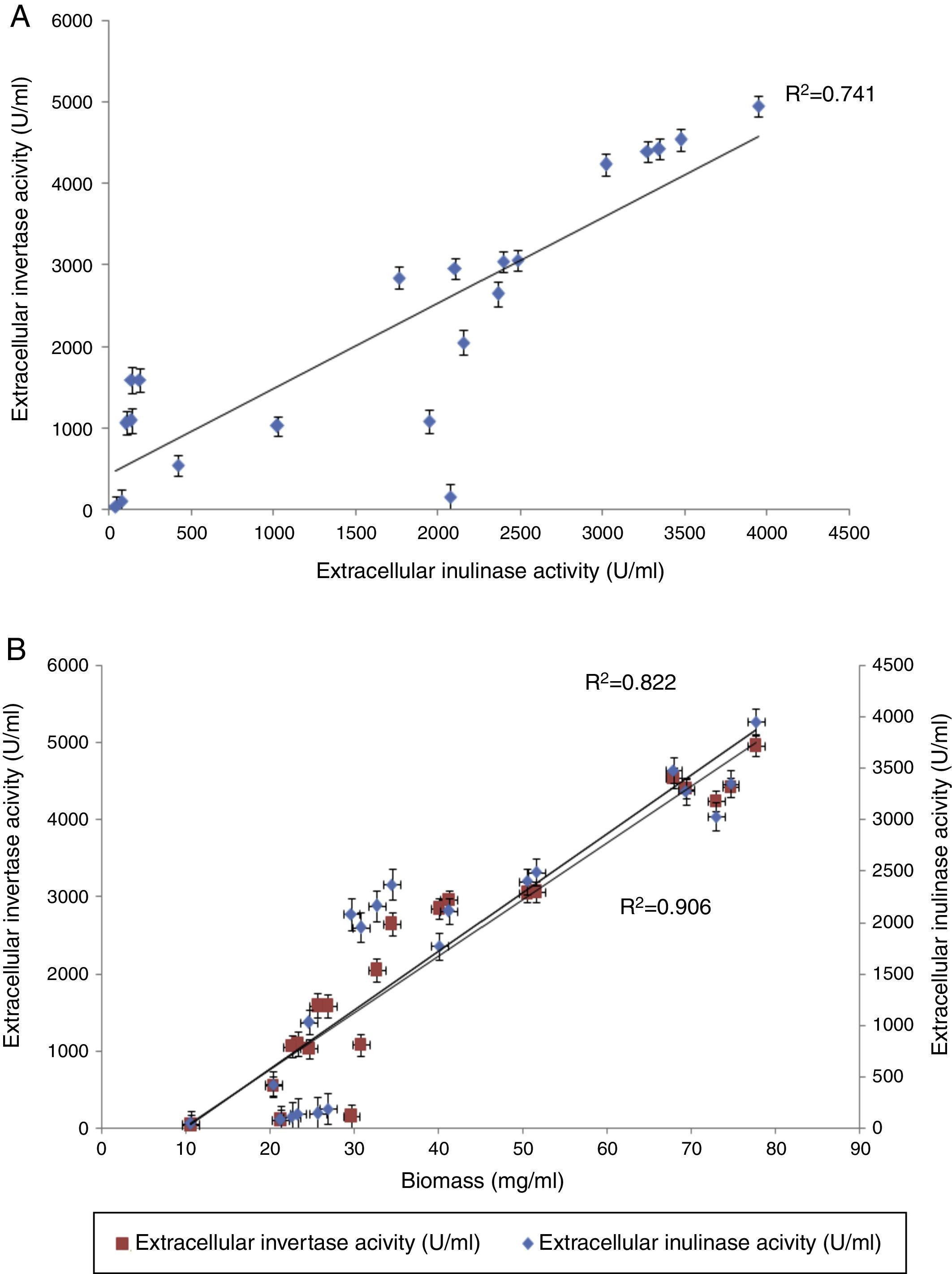
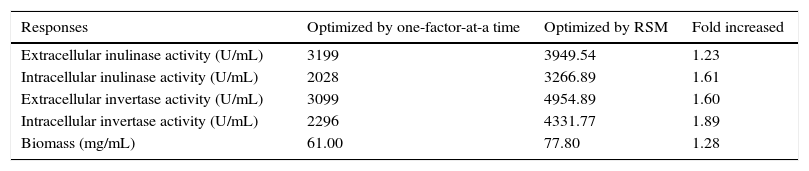
Following optimization, the levels of intra-extracellular inulinase, invertase productions and biomass were significantly increased over 10 times for all responses, in comparison with the control (optimized by one-factor-at-a-time) (data not shown). Table 6 compares the content of intra-extracellular inulinase, invertase production and biomass before and after RSM optimization.12Fig. 6(A) depicts the production profile of inulinase versus invertase. The correlation between the two compounds shows that inulinase is produced during invertase biosynthesis phase. Inulinase serves as a precursor of invertase, hence, increasing inulinase content leads to an elevation of invertase production.12 On the one side, there was a significant positive correlation (r) coefficient at p≤0.01 between the production of inulinase (r=0.98) and invertase (r=0.99) with fungi growth (Fig. 6B).
Results of maximum growth and inulinase and invertase production by A. niger ATCC 20611 before and after RSM optimization.
| Responses | Optimized by one-factor-at-a time | Optimized by RSM | Fold increased |
|---|---|---|---|
| Extracellular inulinase activity (U/mL) | 3199 | 3949.54 | 1.23 |
| Intracellular inulinase activity (U/mL) | 2028 | 3266.89 | 1.61 |
| Extracellular invertase activity (U/mL) | 3099 | 4954.89 | 1.60 |
| Intracellular invertase activity (U/mL) | 2296 | 4331.77 | 1.89 |
| Biomass (mg/mL) | 61.00 | 77.80 | 1.28 |
 Production profile of extracellular inulinase and extracellular invertase production during the fermentation process. (B) Correlation between extracellular inulinase and invertase productions and growth rates. Error bars indicate the mean±standard deviation of three independent experiments.")
Extracellular inulinase and invertase productions by A. niger ATCC 20611. (A) Production profile of extracellular inulinase and extracellular invertase production during the fermentation process. (B) Correlation between extracellular inulinase and invertase productions and growth rates. Error bars indicate the mean±standard deviation of three independent experiments.
The application of RSM allowed the simultaneous determination of the main and interaction effects of physical factors on the productions of intra-extracellular inulinase, invertase and biomass. The optimum values of the process variables were obtained from the quadratic regressions. The adequacies of the models were justified through analysis of variance (ANOVA).
Prob>F-value less than 0.05 indicates that the model terms are significant. Values greater than 0.10 indicate the model terms are not significant. Therefore, it can be assumed that the developed statistical model is reasonably accurate. In other words, fitting the data to various models (linear, two factorial, quadratic and cubic) and their subsequent ANOVA revealed that intra-extracellular inulinase, invertase production and biomass were most suitably defined with the quadratic polynomial model. The model adequacy can be judged from residual's least square which is important to ensure for providing maximum approximation on relationship between factors and response when normal probability is checked by normal probability plot of residuals.
Many attempts have been made by different investigators for identifying suitability of different independent variables as well as process conditions for enhanced intra-extracellular inulinase, invertase production and biomass. The central composite design of response surface can be accurately used for prediction and optimization of intra-extracellular inulinase, invertase production and biomass from A. niger under the experimental conditions employed in this study.
The plot indicates that the residuals (difference between actual and predicted values) follow a normal distribution and form an approximately straight line. “Adeqate Precision” is an index that measures the signal to noise ratio. A ratio greater than 4 is desirable. The computed ratio of 8.5, 9.7, 8.0, 11.5 and 9.4 for intra-extracellular inulinase, invertase production and biomass, respectively, in this study indicates an adequate signal. This means that this model can be used to navigate the design space. The pure error computed, on the other hand, was very low. These results are indicative of maximum predictive responses with constant variance and quadratic model accuracy and demonstrated a good reproducibility of the data obtained in this study. Therefore, the above model can be used to predict the intra-extracellular inulinase, invertase production and biomass within the limits of the experimental factors. Systematic optimization of media showed a wide variation in inulinase, invertase production and biomass.20,21 This variation reflected the importance of optimization to attain higher productivity. From the previous studies optimizing the medium by one factor at a time method, the variables, namely, initial pH, temperature, inoculum size and agitation speed, were selected for further optimization.15,17 In this research, we tried to analyze, model, and interpret the experimental data using RSM as a mathematical modeling system. The optimum conditions were determined using RSM with a four-factor-five-level central composite design (CCD) and also estimated by regression equation to find the best range of parameters and achieve maximum production of intra-extracellular inulinase, invertase and biomass.
It is concluded from the result of this study that acidic conditions can induce A. niger for intra-extracellular inulinase production. Previous studies evaluating the effect of initial pH on inulinase yield reported the highest inulinase production mostly at a near to neutral pH range;2,20,22 nevertheless, acidic fermentation medium in our study induced such elevation in the production of intra-extracellular inulinase. Such similar acidic pH values (6.0–6.5) were demonstrated21 to maximize invertase production in Saccharomyces cerevisiae and maximum biomass from A. niger.20,21 Since the above mentioned enzymes have an almost neutral pH range optimum,2,20–23 therefore, acidic (pH 4.0) and alkaline (pH 12) conditions can block the growth and enzymes that cause intra-extracellular inulinase and invertase to decompose and result in a very low growth rate and enzyme production in the medium. High cell density, in general, is also required for efficient inulinase and invertase biosynthesis.20,21
According to our results, the enzymes productions and growth was increased in the range of temperature (30–35°C).20,21 Elevation in temperature was previously shown to significantly increase inulinase production in other microorganisms.20,21,24,25 In the case of Cryptococcus aureus G7 and A. niger AUP19 the temperature (28°C) was previously reported as the optimum point for inulinase production,2,26 which is near to the temperature value (30°C) found in our study. Such phenomena can be explained by the effect of temperature on intra-extracellular invertase production. It is proven that temperature can influence the efflux of invertase production from fungi cells in which the yield and the specific invertase production rate can be increased by a temperature shift-up to range 30–35°C.17 Such higher temperature for invertase production in our study (30°C) can be explained by the higher temperature necessary for invertase activity than the optimum condition for the cell growth of fungi.17 Generally, the temperature of the cultivation medium was shown to be one of the most important variables. A significant decrease in inulinase, invertase production and biomass was observed at a temperature above or below 30–35°C, which is in agreement with those reported in the literature.27–29 Low enzyme production at higher temperatures, could be due to decreasing oxygen solubility in the medium, or enzyme denaturation.29 In the case of extracellular enzymes, temperatures may influence their secretion, possibly by changing the physical properties of the cell membrane.12
Inoculum size was shown to influence intra-extracellular inulinase, invertase productions and biomass similar to other variables. The results shown 6% of 10 days old culture was found to be the most suitable inoculum size for the enzymes and biomass production by A. niger used in this study; less enzymes production was recorded above and below of this inoculum size. At low inoculum size, the cells present in the culture might not be enough to utilize an essential amount of substrate to produce enzyme.31 However, at high inoculum size, viscosity of fermentation medium might increase due to high growth of fungi resulting in nutritional imbalance in the medium or excessive uptake of nutrients before the cells in the culture were physiologically ready to start enzyme production.31 Compared to other variables, the effects of agitation speed on intra-extracellular inulinase, invertase production and biomass were slightly lower. Generally, suitable agitation leads to better dispersion of the substrate, nutrients and oxygen in the medium and its corresponding availability to the cell.31 It also promotes a reduction in nutrient particle size, favoring the nutrient homogenization in the culture medium, providing a rise in mass transfer rates and nutrient uptake by fungi, which favors fungi growth.23 The results showed a progressive increase in the enzymes productions and growth, when agitation speed was increased to 150rpm. Higher agitation rate; reduce fungal growth because during respiration hydrogen atoms may combine with oxygen to form hydrogen peroxide, which is lethal to the cell.30 Enhancement of growth levels were also reported to increase inulinase and invertase synthesis in A. niger.20,21 Results from this study have demonstrated a significant increase in intra-extracellular inulinase, invertase production and biomass by A. niger through response surface methodology compared to non-optimization condition. Intra-extracellular inulinase, invertase production and biomass were the result of a synergistic combination of effective parameter interactions and these parameters were in equilibrium content. The optimized conditions for the production of intra-extracellular inulinase, invertase and biomass, as reported in this study, can be of several advantages to a wide range of industries particularly food manufacturers.
Conflicts of interestThe authors report no conflicts of interest in this work.