



The aim of this study was to evaluate the effect of a novel phytoestrogen, α-Zearalanol, on Alzheimer's disease-related memory impairment and neuronal oxidation in ovariectomized mice.
METHODSFemale C57/BL6 mice were ovariectomized or received sham operations and treatment with equivalent doses of 17β-estradiol or α-Zearalanol for 8 weeks. Their spatial learning and memory were analyzed using the Morris water maze test. The antioxidant enzyme activities and reactive oxygen species generation, neuronal DNA oxidation, and MutT homolog 1 expression in the hippocampus were measured.
RESULTSTreatment with 17β-estradiol or α-Zearalanol significantly improved spatial learning and memory performance in ovariectomized mice. In addition, 17β-estradiol and α-Zearalanol attenuated the decrease in antioxidant enzyme activities and increased reactive oxygen species production in ovariectomized mice. The findings indicated a significant elevation in hippocampi neuronal DNA oxidation and reduction in MutT homolog 1 expression in estrogen-deficient mice, but supplementation with 17β-estradiol or α-Zearalanol efficaciously ameliorated this situation.
CONCLUSIONThese results demonstrate that α-Zearalanol is potentially beneficial for improving memory impairments and neuronal oxidation damage in a manner similar to that of 17β-estradiol. Therefore, the compound may be a potential therapeutic agent that can ameliorate neurodegenerative disorders related to estrogen deficiency.
In addition to the regulation of reproductive functions, estrogens may exert several neuroprotective effects on the brain. Estrogens may facilitate axonal sprouting, neuronal repair, and neurogenesis induction (1), and they may also reduce reactive oxygen species (ROS)-related neuronal damage and protect neuronal nucleic acids against the detrimental consequences of oxidative stress (2). These beneficial effects have led to the use of estrogen replacement therapy as an effective method to reduce the risk and severity of Alzheimer's disease (AD) in postmenopausal women, who are burdened with a higher incidence of AD than men of the same age and premenopausal women (3–5). However, the fact that estrogen replacement therapy may predispose women to a much higher incidence of breast and endometrial cancers has undoubtedly compromised or jeopardized the clinical application of estrogen (6). Thus, the search for safe and effective estrogen substitutes has gained increased importance.
Recently, plant-derived phytoestrogens (which possess the physiological properties of animal-derived estrogen) have been demonstrated to act as potential replacements for estrogen. In our previous study, we confirmed the neuroprotective effect of the phytoestrogen α-Zearalanol (α-ZAL), and it was demonstrated to have low side-effects on the reproductive system compared with animal-derived estrogen (7). α-ZAL is a reductive product of Gibberella zeae metabolites that is abundant in plants and vegetables, including soybeans, wheat, grapes, radishes, celery, spinach, and apples (8), and can be safely metabolized in the body (9). Our previous reports implied that α-ZAL could provide a beneficial effect on the maintenance of the oxidant-antioxidant balance (10,11). This effect thereby allows α-ZAL to apparently ameliorate the neuron damage induced by oxidative stress. Recent results have suggested that the oxidative damage of DNA plays a meaningful role in the pathogenesis of AD (12,13), and estrogen deficiency supplementation appears to provide a powerful protective effect against DNA oxidation, which contributes to the improvement of AD-related cognitive deficits (14,15). However, whether α-ZAL exerts the same influence on cognition and neuronal oxidation remains unclear. In the present study, we investigated the effect of α-ZAL on memory impairment and neuronal DNA oxidation in estrogen-deficient mice and the underlying molecular mechanisms, with 17β-estradiol (E2) serving as an estrogen-positive control.
MATERIALS AND METHODSMaterialsα-ZAL was a gift from Prof. Shunling Dai at Perking Union Medical College. E2, 2‘,7‘-dichlorofluorescein diacetate (DCFH-DA), and RNase were purchased from Sigma-Aldrich (Saint Louis, MO, USA). The protease inhibitor mixture and BCA Protein Assay Kit were obtained from Pierce Biotechnology (Rockford, IL, USA). The reagent kits used for the measurement of superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px) were purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing, China). In addition, 8-oxo-7,8-dihydrodeoxyguanosine (8-oxoguanine) and MutT homolog 1 (MTH1) antibodies were purchased from Abcam (Abcam, England). β-actin antibody was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The avidin-biotin-peroxidase and DAB kits used for immunohistochemical detection were obtained from Zhongshang Goldenbridge Biotechnology (Beijing, China). All other chemicals used were of the highest commercial grade available.
Animals and treatmentFemale C57/BL6 mice (8 weeks old) were purchased from Weitong Lihua Experimental Animal Center (Beijing, China). All of the mice were housed in groups of 5 on a 12-hour light/dark schedule. The mice were allowed free access to mouse lab chow and water. Animal treatment and maintenance were conducted in accordance with the guidelines established by the National Institutes of Health for the care and use of laboratory animals and were approved by the Animal Care Committee of the Peking Union Medical College and the Chinese Academy of Medical Science.
After 5 days of habituation, the animals were anesthetized with ketamine (100 mg/kg) and xylazine (20 mg/kg), and a bilateral ovariectomy (OVX) or sham-operation was performed. Following surgery, the mice were allowed to recover for 14 days and then assigned to one of the following four groups (n = 10 in each group): control group, Sham-operation+ olive oil; OVX group, OVX+olive oil; E2 group, OVX+E2; and α-ZAL group, OVX+α-ZAL. Mice in the E2 and α-ZAL groups received intraperitoneal (i.p.) injections of E2 (0.5 mg/kg) and α-ZAL (0.5 mg/kg), respectively. The E2 and α-ZAL chemicals were dissolved in olive oil. Mice in the control group and OVX group received an equal volume of olive oil. The intraperitoneal injections were administered every three days. Treatment continued for 8 weeks. The above animal treatment procedures were conducted as described previously in a paper by our group (7).
Morris water mazeThe method used for training and testing mice in the Morris water maze was similar to that previously described (16). The animals were trained in a water tank (100 cm in diameter) located in a lit room with visual cues. A black escape platform (9.5 cm in diameter) was submerged 1 cm below the water surface in the middle of the southwest quadrant of the tank. The water was maintained at 21±1°C and mixed with milk powder to obscure the platform. The test began on the 52nd day after injection. The Morris water maze protocol consisted of a 4-day learning and memory training period and a probe test on day 5. For each training trial, each mouse was randomly placed in the water, facing the tank wall, at one of the four starting points (the midpoints of the four quadrants of the tank) and was allowed a maximum time of 60 s to find the platform. Mice that found the platform were allowed to remain on the platform for 10 s and subsequently placed into an empty and warm cage for 15 min before the next trial began. If a mouse did not reach the platform within 60 s, it was gently guided to the platform by the experimenter, where it remained for 10 s. Four training trials were conducted each day for 4 consecutive days. After 4 days of trials, the probe test was conducted by removing the platform and allowing each mouse to swim freely for 60 s inside the tank. The time spent in the southwest quadrant (explore time; location where the platform was originally placed), the escape latency (time to reach the original platform position), and the swimming speed were recorded by a video camera and computer system and subsequently analyzed by a computer (the hardware and software were provided by HVS IMAGE P.O. Box 100, Hampton TW12 2YD, UK).
Sample preparationAfter the behavioral tests, six animals in each group were deeply anesthetized by isoflurane inhalation and sacrificed by decapitation. The brain was removed and dissected on ice. The hippocampus was removed and stored at -80°C for Western blotting and biochemical analysis. The remaining 4 animals were perfused with phosphate-buffered saline (PBS) and 4% paraformaldehyde under deeply anaesthetized conditions. Their brains were removed and post-fixed in 10% sucrose/4% paraformaldehyde for 8 h at 4°C and then placed in 20% sucrose/PBS at 4°C overnight. Using a cryostat microtome, 25-μm frozen sections were cut for the 8-oxoguanine experiment.
Measurement of antioxidant enzyme activities and ROS generationThe SOD and GSH-Px activities were assessed using commercially available kits following the manufacturer's instructions. The levels of ROS production in the hippocampal tissues were measured fluorimetrically using DCFH-DA as previously described (17). Briefly, 20 mg of tissue was pooled in 500 μL of cold lysis buffer (50 mM Tris–HCl, pH 7.5; 50 mM NaCl; 1 mM EGTA; 10 mM EDTA; 1 mM NaVO4; 1 mM PMSF, supplemented with 0.1% v/v Triton X-100 and protease inhibitor cocktail). The samples were homogenized for 15 s with a sonic dismembrator and centrifuged at 10,000 rpm at 4°C for 20 min. The supernatants were filtered through 0.45-μm hydrophilic filters. The protein concentration of the lysates was estimated using a BCA Protein Assay Kit. The homogenate samples from different groups were incubated with 10 μM DCF-DA at 37°C for 1 h, and the fluorescence intensity was measured at an excitation wavelength of 485 nm and emission wavelength of 525 nm. The ROS level was presented as the percentage of increase with respect to the control group.
Western blottingHippocampal tissue was lysed using ice-cold RIPA buffer (150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS, 5 mM ethylenediamine tetraacetic acid, 50 mM Tris-HCl, pH 8.0) supplemented with protease inhibitor cocktail and then centrifuged at 13,000 rpm for 10 min at 4°C. The supernatant was collected, and the protein concentration was estimated using a BCA Protein Assay Kit with bovine serum albumin as the standard. In addition, 30 μg of protein was loaded into SDS PAGE gels after adding the sample buffer and boiling. The proteins were then transferred to nitrocellulose membranes. After transfer, the membranes were blocked with Tris-buffered saline (TBS) containing 0.1% Tween-20 (TBST) and 5% non-fat milk for 2 h at room temperature and then overnight at 4°C with the following primary antibodies: anti-MTH1 (1:500) and anti-β-actin (1:2000). After washing, the membranes were incubated with goat radish peroxidase-conjugated secondary antibody (1:5000), followed by detection using enhanced chemiluminescence. The scanned images of the developed blots were quantified using densitometry functions in Image-Pro Express 4.0 software (Media Cybernetics, Inc., Bethesda, MD, USA).
Immunohistochemical detection of 8-oxoguanineFor the detection of oxidized nuclear DNA in neurons, the sections were first pretreated in 10 mM Tris–HCl (pH 7.4) containing 15 mM NaCl and 5 mg/ml DNase-free RNase for 60 min at 37°C to eliminate cellular RNA. Then, the RNase-treated sections were incubated in 3 N HCl for 30 min at room temperature to denature the DNA, and the rinsed sections were immersed in a solution of 3% H2O2 in methanol/PBS (1:1) for 5 min at room temperature and then incubated with blocking buffer for 2 h at room temperature, followed by the primary monoclonal antibody, anti-8-oxoguanine (1:100), overnight at 4°C. After washing with PBS, the sections were incubated with a goat anti-rat biotin-conjugated secondary antibody for 1 h, followed by peroxidase-conjugated streptavidin. Visualization was performed using 3-3'-diaminobenzidine (DAB). The final immunoreaction product of DAB was brown-yellow in color. The sections were washed with PBS, dehydrated in graded ethanol solutions, cleared in xylene, and coverslipped.
Quantitative 8-oxoguanine analysisAll acquired digital images were processed uniformly at a threshold in gray-scale mode to subtract any background corresponding to the area without tissue using a Leica microscope (Leica, Germany). The optical density (OD) in an area comprising the cytoplasm and nucleus was determined using the Leica Q550CW Image Analysis System linked to a Leica microscope. Six adjacent fields of the CA1 and DG subregions of the hippocampi from each mouse were selected. The OD value was corrected for background by subtracting the OD of the white matter on the same section and averaged. All measurements were performed under the same optical and light conditions, and electronic shading correction was performed to compensate for any unevenness that may be present in the illumination.
Statistical analysisData are expressed as the mean ± SEM. Differences between mean values were evaluated by analysis of variance (ANOVA) followed by post-hoc tests. Statistical significance was set at p≤0.05.
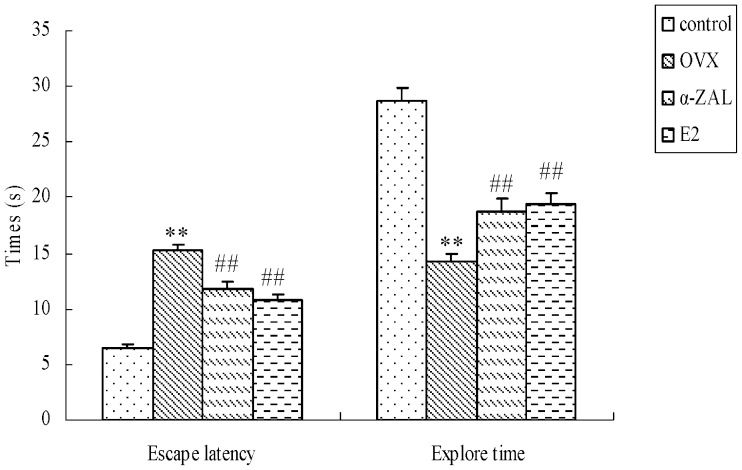
RESULTSMorris water maze testIn the Morris water maze test, the swimming speeds of the mice in the control and OVX groups were 20.9 and 22.2 cm/s, respectively, and there were no significant differences in the speeds between the groups (data not shown). The time spent in the target quadrant (explore time) and the escape latency in the probe test are shown in Figure 1). The data revealed that OVX-treated animals exhibited significantly longer escape latency (F(1,19) = 95.31, p<0.001) and less time spent in the target quadrant (F(1,19) = 76.59, p<0.001) in probe tests compared with the animals in the control group. α-ZAL administration significantly shortened the escape latency (F(1,19) = 19.79, p<0.001) and lengthened the explore time (F(1,19) = 21.51, p<0.001) compared with the OVX group, as did the administration of E2.
. ∗∗p<0.01 vs. control group; ##p<0.01 vs. OVX group (n = 10 in each group).")
The effects of α-ZAL treatment on escape latency and explore time during the Morris water maze in OVX-induced, memory-deficient mice. The training began on the 52nd day after α-ZAL or E2 administration. Four training trials each day were conducted for four consecutive days. The escape latency and explore time from the Morris water maze are shown for the probe test session (the results from the trial at the 56th). ∗∗p<0.01 vs. control group; ##p<0.01 vs. OVX group (n = 10 in each group).
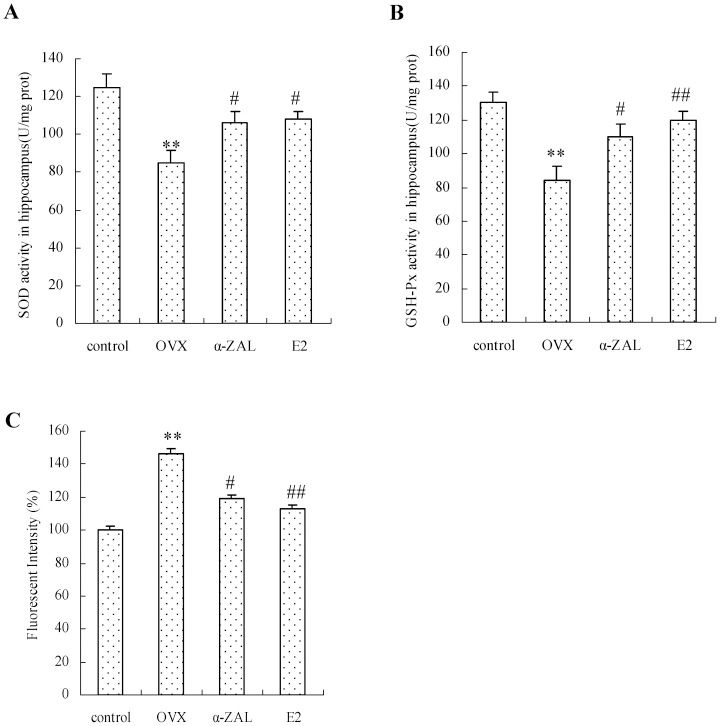
Our data revealed that the activities of the anti-oxidant enzymes, including SOD and GSH-Px, were significantly decreased by 32 and 35%, respectively, in OVX animals compared with the control group (Figure 2A and B), and OVX administration resulted in a 1.5-fold increase in the ROS level relative to the control group (Figure 2C). However, α-ZAL administration effectively attenuated the decrease in SOD (α-ZAL versus OVX: F(1,13) = 5.37, p = 0.039) and GSH-Px (α-ZAL versus OVX: F(1,13) = 5.49, p = 0.037) and the increase in ROS (α-ZAL versus OVX: F(1,13) = 6.08, p = 0.031) in OVX mice, as did E2 administration. Although the effects of α-ZAL on ROS levels, SOD activity, and GSH-Px activity were less than those of E2, no significant difference was observed in the enzyme activities and ROS levels between the E2 and α-ZAL groups.
 and the increase in ROS generation (C) in hippocampal tissues in OVX mice. The data are expressed as the mean ± SEM (n = 6 in each group). ∗∗ p<0.01 vs. control group; # p<0.05 and ## p<0.01 vs. OVX group.")
E2 and α-ZAL effectively reversed the decrease in anti-oxidative enzyme activities (A and B) and the increase in ROS generation (C) in hippocampal tissues in OVX mice. The data are expressed as the mean ± SEM (n = 6 in each group). ∗∗ p<0.01 vs. control group; # p<0.05 and ## p<0.01 vs. OVX group.
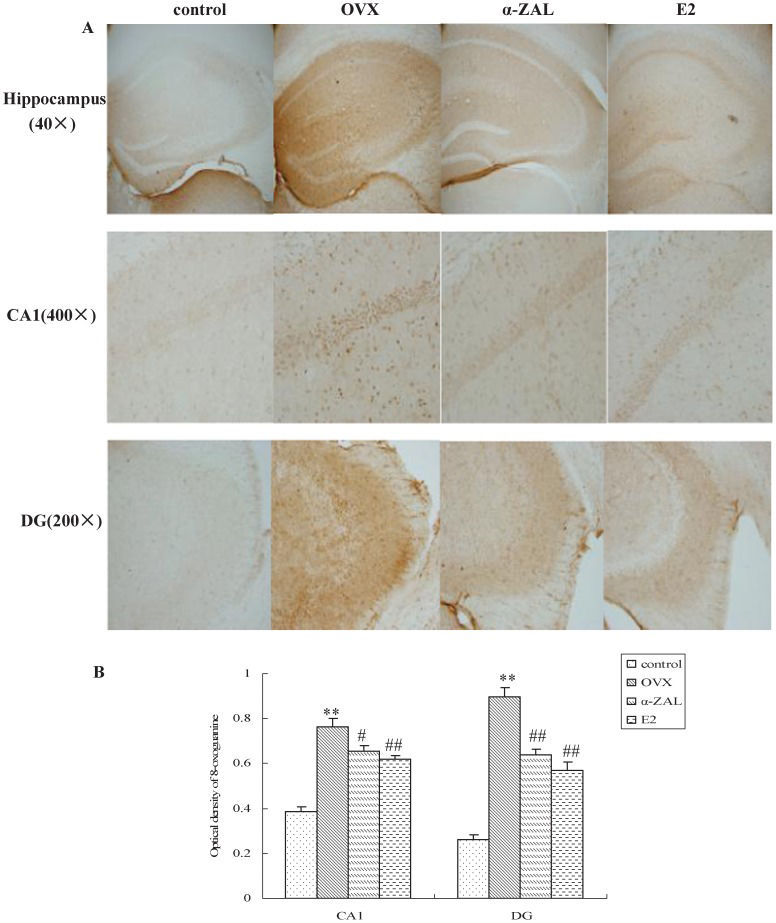
To detect oxidized nuclear DNA in the hippocampi, immunostaining with antibodies against 8-oxoguanine was performed. As shown in Figure 3A, low immunoreactivity was observed in the nuclei of the control animals. When antibodies against 8-oxoguanine were applied to the samples from OVX mice, specific staining of the nuclei was observed in the CA1 and DG regions of the mice hippocampi. The average optical density was measured for the subregions of the hippocampus in the mice, and the digitized data are shown in Figure 3B. A decrease in 8-oxoguanine in the CA1 and DG regions of the hippocampus was observed in mice that received E2 compared with the mice treated with OVX alone (F(1,23) = 12.55, p = 0.002 for the CA1 region, and F(1,23) = 35.56, p<0.001 for the DG region). The immunohistochemistry assay data also indicated that the levels of 8-oxoguanine were significantly decreased in the presence of α-ZAL compared with OVX alone (F(1,23) = 5.77, p = 0.025 for the CA1 region, and F(1,23) = 29.35, p<0.001 for the DG region), indicating a low level of oxidative DNA damage in E2- or α-ZAL-treated mice.
. The first row shows a lower magnification of the hippocampus (40×), whereas the magnified views of the CA1 (400×) and DG (200×) subregions are shown in the middle and last rows, respectively. B. Quantification of 8-oxoguanine immunolabeling in the CA1 and DG regions of the hippocampus. n = 4 in each group. The results represent at least three independent experiments in each animal. ∗∗p<0.01 vs. control group; # p<0.05 and ## p<0.01 vs. OVX group.")
The effect of E2 and α-ZAL treatment on the levels of 8-oxoguanine in the hippocampi of mice. A. The immunohistochemical analyses of free-floating sections (25 μm in thickness). The first row shows a lower magnification of the hippocampus (40×), whereas the magnified views of the CA1 (400×) and DG (200×) subregions are shown in the middle and last rows, respectively. B. Quantification of 8-oxoguanine immunolabeling in the CA1 and DG regions of the hippocampus. n = 4 in each group. The results represent at least three independent experiments in each animal. ∗∗p<0.01 vs. control group; # p<0.05 and ## p<0.01 vs. OVX group.
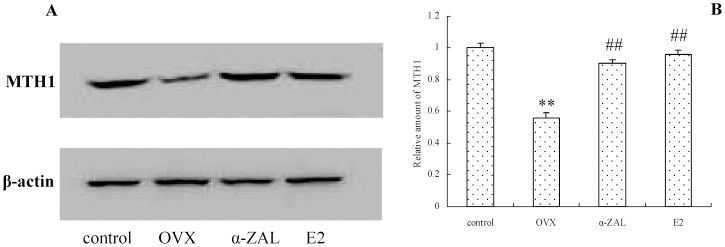
We also examined the expression of MTH1, which possesses the ability to hydrolyze the 8-oxoguanine-containing deoxyribonucleoside triphosphate, which thereby facilitates DNA repair and ameliorates oxidized DNA accumulation. Western blot (Figure 4) analyses indicated that the protein expression of MTH1 was decreased in the brains from OVX mice (control versus OVX: F(1,13) = 105.37, p<0.001); however, after E2 or α-ZAL treatment, these changes were reversed, and they were significantly different compared with those in the OVX group (α-ZAL versus OVX: F(1,13) = 81.97, p<0.001, and E2 versus OVX: F(1,13) = 84.59, p<0.001).

The effect of α-ZAL treatment on the protein expression levels of MTH1 in the hippocampi of mice. A. MTH1 protein expression determined by Western blot analysis. B. Quantitative analysis of MTH1 protein levels by densitometry. The Western blot data were normalized by setting the control group value to 1. ∗∗p<0.05 vs. control group; ##p<0.01 vs. OVX group.
As a neuromodulatory and neuroprotective hormone, estrogen exerts useful effects associated with the prevention of AD. However, the use of estrogen is imperiled due to the uncertainty of the increased risk of developing breast as well as endometrial cancers in women taking estrogen. Our published data have shown that treatment with the phytoestrogen α-ZAL can provide protection against neuronal injury both in vivo and in vitro, but it yields fewer side effects on the breasts and uterus than estrogen (7,10). In this study, we demonstrated that α-ZAL exerted an in vivo improvement in estrogen deficiency-induced spatial learning and memory impairment. The neuroprotection afforded by α-ZAL may be mediated through the regulation of antioxidase activities to improve oxidative stress and expression of MTH1 to decrease neuronal oxidative damage, in a manner similar to that of E2.
The reduction of estrogen levels is an important factor associated with memory impairment in postmenopausal women (3–5). OVX animals have been frequently used as a model of hormone deprivation to study post-menopausal changes in adult females. Therefore, we used this model to evaluate the effects of E2 and α-ZAL on the learning and memory abilities of OVX mice. Results from our study indicated significant decreases in explore time but increases in escape latency in OVX animals compared with control animals. These results are in agreement with those of Azizi-Malekabadi et al., who demonstrated that ovarian hormone loss due to ovariectomy in animals impairs water maze performance (18). However, treatment with E2 significantly improved spatial learning and memory performance in OVX mice, as did α-ZAL treatment. The results suggested that the use of α-ZAL may be invaluable in producing neuroprotective effects similar to E2. The cognitive deficits correlate well with the increased oxidative stress in ovariectomized animals (19,20); thus, the observation of the high oxidative stress condition that was indicated by low SOD and GSH-Px activities and increased ROS levels in OVX animals in the present study was not surprising. However, increases in ROS generation and decreases in antioxidase activities induced by OVX were significantly attenuated when the animals received supplementation with 17β-E2 or α-ZAL, which thereby decreased oxidative stress by affecting ROS formation and anti-oxidant enzyme activities in a beneficial manner to limit neuronal damage.
In response to oxidative stress, neurons are attacked by ROS, resulting in widespread oxidative damage. Protein oxidation may affect neuron function by damaging enzymes, and lipid oxidation can cause structural membrane damage that may impair cellular stabilization (21,22). Nucleic acids exposed to ROS also generate oxidative damage that may lead to neuron dysfunction or even death (23). Several studies have demonstrated that oxidative DNA damage is significantly increased in neuronal samples from the brains of AD patients (24,25,12). Moreover, the existing studies also revealed that the accumulation of oxidative DNA damage in hippocampal neurons is a trigger for cognitive dysfunction in AD (26). Therefore, we assayed the level of 8-oxoguanine, the most commonly analyzed biomarker of DNA oxidative damage (27). The results of the 8-oxoguanine immunohistochemical analysis performed in the present study revealed that the optical density of 8-oxoguanine in the hippocampus increased in OVX mice, which was partially reversed by E2 or α-ZAL administration. These results demonstrated the ability of estrogen to dampen the DNA oxidation process. MTH1 possesses the specific enzymatic activity necessary to convert the deleterious 8-oxoguanine-containing deoxyribonucleoside triphosphate into monophosphates, thus preventing its misincorporation into DNA and thereby minimizing the accumulation of oxidized DNA in neurons. Additionally, MTH1 activity is significantly decreased in AD subjects (28,29). Therefore, we examined the effect of MTH1 on estrogen-mediated DNA protection. The Western blot results revealed a sharp decrease in MTH1 protein expression levels in OVX mice, whereas E2 and α-ZAL significantly inhibited these changes. Obviously, the effect of E2 and α-ZAL on MTH1 expression is beneficial for attenuating DNA oxidation, which, overall, contributes to improvements in the neuronal dysfunction and cognitive impairments induced by ovarian hormone loss. In addition, we observed that E2 or α-ZAL administration cannot completely reverse DNA oxidation. This result suggests that there are more factors involved in oxidative DNA damage in OVX animals, and further studies are in progress to address these possibilities.
In summary, in the current study, we compared the ability of E2 and α-ZAL to ameliorate memory deficits and neuronal oxidation due to estrogen deficiency and assessed whether α-ZAL is associated with the same mechanism for the amelioration of memory impairment and neuronal DNA oxidative damage as E2. This study had a few limitations that should be noted. For example, (1) a wider range of α-ZAL concentrations should be chosen to investigate the changes in neuronal oxidative damage, and (2) a greater number of animals should be used to analyze the clinical efficacy and potential toxicity of α-ZAL. It appears that α-ZAL may be a potential agent that can be used to prevent the progression of neurodegenerative disorders related to estrogen deficiency.
AUTHOR CONTRIBUTIONSDong Y and Wang Y contributed equally to this study. Dong Y and Zuo P designed the study. Dong Y, Wang Y, Liu Y, and Yang N performed the research. Dong Y and Wang Y analyzed the data. Dong Y and Wang Y drafted the manuscript. Zuo P reviewed and approved the final version of the manuscript.
This work was supported by a grant from the National 973 Fundamental Project of China (2010CB934002), the Science and Technology project of Yunnan Provincial Science and Technology Department (2011FZ005), and the scientific research foundation of the Yunnan Provincial Education Department (2012Z084).
No potential conflict of interest was reported.