





Recuperamos 22 aislados de estafilococos coagulasa negativos resistentes al linezolid de nuestro hospital para su identificación, sensibilidad, perfil epidemiológico, mecanismos de la resistencia al linezolid y posibles combinaciones antibióticas sinérgicas.
MétodosLa identificación de los aislados fue realizada mediante espectrometría de masas (Vitek-MS, BioMérieux). La sensibilidad se realizó con el sistema Vitek-2 (bioMérieux) y el método de microdilución en caldo según las recomendaciones del CLSI. Se realizó electroforesis en gel de campo pulsado (PFGE) para estudiar la relación genética de los aislados. Los mecanismos de la resistencia al linezolid fueron evaluados por PCR/secuenciación: presencia del gen cfr, mutaciones puntuales en el dominio V del ARN ribosomal 23S y mutaciones ribosomales adicionales (en los genes rplC, rplD y rplV).
La actividad in vitro del linezolid fue estudiada por separado y en combinación con otros 3 antibióticos utilizando tiras de E-test.
ResultadosVeinte aislados fueron identificados como Staphylococcus epidermidis y 2 como Staphylococcus hominis. La PFGE demostró que los aislados pertenecían a diferentes clones: 21 de los aislados presentaron mutaciones en la región del dominio V del ARNr 23S y en el 54,5% fue encontrado el gen cfr.
La administración previa de linezolid fue documentada en la mayor parte de casos.
El linezolid en combinación con gentamicina mostró una actividad sinérgica en el 45,5% de los aislados.
ConclusionesStaphylococcus epidermidis fue la especie de estafilococos coagulasa negativos resistentes al linezolid más frecuentes. Los valores de CMI fueron además elevados para otros antiestafilocócicos. Todas las cepas presentaron varios mecanismos de resistencia al linezolid. Nuestros datos sugieren que linezolid más gentamicina podría ser una combinación sinérgica frente a los estafilococos coagulasa negativos resistentes al linezolid.
We recovered 22 coagulase-negative staphylococci isolates in our hospital to study their identity, susceptibility, epidemiological profile, linezolid resistance mechanisms, and the possibilities of different antibiotic combinations.
MethodsIsolate identification was performed using mass spectrometry (Vitek-MS, bioMérieux). Susceptibility testing was carried out with the Vitek-2 system and the broth microdilution method according to CLSI guidelines. Pulsed-field gel electrophoresis (PFGE) was performed to analyze the genetic relationship between isolates. Linezolid resistance mechanisms were evaluated by PCR/sequencing: presence of cfr gene, point mutations in domain V of 23S ribosomal RNA and additional ribosomal mutations (in the rplC, rplD and rplV genes).
The in vitro activity of linezolid was investigated alone and in combination with another three antibiotics acting on different cellular targets, using E-test strips.
ResultsTwenty isolates were identified as Staphylococcus epidermidis, and 2 as Staphylococcus hominis. PFGE showed that isolates belonged to diverse clones, 21 of them presented mutations in the domain V region of 23S rRNA and the cfr gene was found in 54.5%.
Prior administration of linezolid was documented in most of cases.
Linezolid in combination with gentamicin showed a synergistic activity in 45.5% of isolates.
ConclusionsStaphylococcus epidermidis was the most prevalent linezolid-resistant coagulase-negative staphylococci. All isolates showed increased MIC values compared to other anti-staphylococcal drugs and several linezolid resistance mechanisms. Our data suggest that linezolid plus gentamicin could be a synergistic combination against linezolid-resistant coagulase-negative staphylococci.
El linezolid es un antimicrobiano perteneciente al grupo de las oxazolidinonas, activo frente a un gran número de bacterias grampositivas, incluyendo Staphylococcus aureus resistentes a la meticilina, Enterococcus resistentes a los glucopéptidos y una gran variedad de estreptococos. Su mecanismo de acción se efectúa mediante unión al dominio V del 23S ARN ribosomal correspondiente a la subunidad 50S, inhibiendo la síntesis de proteínas bacterianas1.
Diferentes especies de Staphylococcus coagulasa negativos (SCN) residentes habituales en piel y mucosas de sujetos sanos pueden, en ocasiones, ser la causa de infecciones sistémicas, especialmente entre portadores de prótesis, catéteres u otros dispositivos, convirtiéndose en una de las principales causas de bacteriemia2.
Aunque, hasta el momento, la mayoría de los aislados clínicos de SCN se muestran sensibles al linezolid, se describen cada vez con mayor frecuencia cepas que presentan resistencia a este antimicrobiano3. Teniendo en cuenta la naturaleza sintética de esta molécula, originalmente no deberían existir reservorios naturales que la explicaran, por lo que los mecanismos de resistencia primarios deberían ser de naturaleza mutacional con baja frecuencia de bacterias resistentes. Así, el mecanismo más frecuentemente observado es el que conlleva mutaciones en el dominio V del 23S ribosomal, de una o más copias del gen codificante, especialmente la denominada G2576T4. Otras mutaciones o sustituciones en distintas proteínas ribosomales también han sido descritas últimamente5-7. Sin embargo, otro mecanismo no mutacional, la adquisición del gen cfr, puede estar implicado como causa de la resistencia y ser el origen de brotes transmisibles a través del plásmido adquirido y, por tanto, con un poder de diseminación mucho más elevado8.
En nuestro hospital hemos ido coleccionando en este último año diferentes aislamientos clínicos de SCN que mostraban resistencia al linezolid cuando se realizaba una determinación rutinaria de su sensibilidad.
El objeto de este estudio ha sido la caracterización de estas cepas resistentes al linezolid; conocer su patrón de resistencia a otros antiestafilocócicos, como la vancomicina, la gentamicina, la daptomicina y rifampicina; la determinación de su mecanismo de resistencia al linezolid, y la exploración de diferentes combinaciones de linezolid con otros antimicrobianos ante la posibilidad de establecer asociaciones sinérgicas con ellos.
Material y métodosMicroorganismosSe estudiaron 22 cepas de Staphylococcus coagulasa negativos resistentes al linezolid (SCN-RL) aisladas a partir de octubre de 2010 en nuestro hospital, procedentes de muestras clínicas de diferentes pacientes. La localización de los pacientes fue la siguiente: 10 de ellos estaban ingresados en la unidad de cuidados intensivos, 5 procedían de la unidad de reanimación, otros 4 estaban ingresados en el servicio de medicina interna y los 3 restantes en otras unidades. Catorce cepas se aislaron de hemocultivos, 3 de cultivos de catéteres y 5 de diferentes localizaciones.
Identificación bacterianaLa identificación se realizó inicialmente usando tarjetas GP pertenecientes al sistema Vitek-2 (bioMérieux, Francia). La identificación definitiva de cada aislado se llevó a cabo mediante espectrometría de masas utilizando el sistema Vitek-MS (bioMérieux, Francia).
Electroforesis en campo pulsadoLa electroforesis en campo pulsado se realizó mediante una modificación del protocolo de Murchan et al.9 digiriendo el ADN incluido en bloques de agarosa con SmaI y cargándolos en geles de agarosa al 1% peso/volumen. La electroforesis se realizó en un tampón Tris-Borato 0,5X en el sistema CHEF-DRII (Bio-Rad Lab., Reino Unido) bajo las siguientes condiciones: pulsos de 0,1 a 30s, 6V/cm durante 24h a una temperatura de 12-14°C. Los geles fueron teñidos con bromuro de etidio, visualizados con luz ultravioleta y fotografiados. Los patrones de los fragmentos fueron interpretados según la descripción de Tenover et al.10.
Estudio de sensibilidadEl estudio de sensibilidad inicial se llevó a cabo mediante tarjetas AST-P588 del sistema Vitek-2 (bioMérieux, Francia), confirmándose posteriormente por microdilución frente a los siguientes antimicrobianos: linezolid, vancomicina, gentamicina, rifampicina y daptomicina, siguiendo los criterios de actuación e interpretación del CLSI11.
Mecanismos de resistencia al linezolidDetección del gen cfrLa presencia del gen cfr se comprobó mediante PCR, según las condiciones descritas por Kehrenberg y Schwarz12. Como control positivo se usó una cepa de S. epidermidis resistente a linezolid de nuestra colección.
Detección de la mutación G2576T en dominioVSe amplificó la región del dominio Schwarz V que incluye el nucleótido en posición 2576 según la numeración empleada para Escherichia coli utilizando los cebadores descritos por Pillai et al.13. Las secuencias de nucleótidos se compararon con las de la cepa patrón S. aureus ATCC12600.
RiboproteínasPara la amplificación de los genes rplC, rplD y rplV que codifican las proteínas ribosomales L3, L4 y L22 utilizamos los cebadores descritos por Mendes et al.14. Los amplicones fueron secuenciados y las secuencias correspondientes se analizaron usando el sistema DNASTAR (Madison, WI, EE.UU.). Las secuencias para L3 se compararon con las de S. epidermidis ATCC 12228 y S. hominis SK119 y para L4 y L22 con S. epidermidis RP62A y S. hominis C80.
Combinaciones de antimicrobianosSe evaluó la actividad de las siguientes combinaciones antibióticas: linezolid más vancomicina (LZ+VA), linezolid más rifampicina (LZ+RIF) y linezolid más gentamicina (LZ+CN) mediante tiras E-test (bioMérieux, Francia) siguiendo las recomendaciones del fabricante.
El índice fraccional de concentración inhibitoria (FICI) se calculó a partir de la suma de las fracciones de las concentraciones inhibitorias de acuerdo con la fórmula:

Los resultados obtenidos se categorizaron según el siguiente criterio: FICI≤0,5, sinérgico; 0,5<FICI<1, parcialmente sinérgico; FICI=1, aditivo; 1<FICI≤4 indiferente y FICI>4 antagónico.
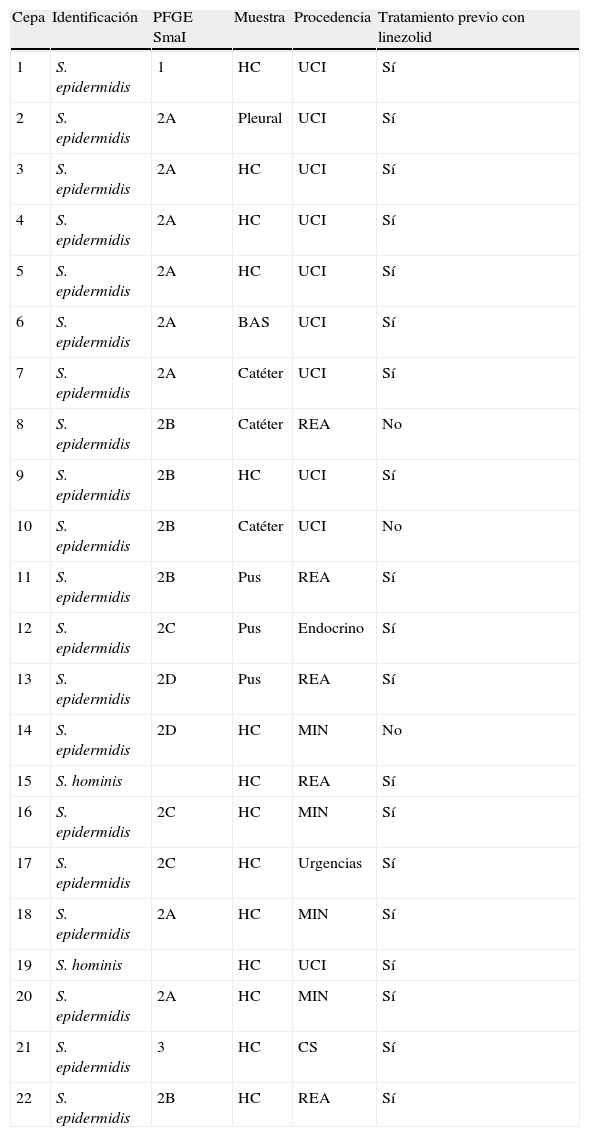
ResultadosDe las 22 cepas seleccionadas para el estudio, 20 fueron identificadas mediante espectrometría como S. epidermidis y 2 como S. hominis. En 15 casos los aislamientos procedían de unidades de cuidados críticos. Catorce cepas se obtuvieron de hemocultivos, 3 aislamientos a partir del cultivo de catéteres y los 5 restantes se recogieron de exudados purulentos. El 86,3% de los pacientes en los que se aisló SCN-RL habían recibido previamente tratamiento con linezolid (tabla 1).
Identificación, procedencia y clonalidad de las cepas estudiadas
| Cepa | Identificación | PFGE SmaI | Muestra | Procedencia | Tratamiento previo con linezolid |
| 1 | S. epidermidis | 1 | HC | UCI | Sí |
| 2 | S. epidermidis | 2A | Pleural | UCI | Sí |
| 3 | S. epidermidis | 2A | HC | UCI | Sí |
| 4 | S. epidermidis | 2A | HC | UCI | Sí |
| 5 | S. epidermidis | 2A | HC | UCI | Sí |
| 6 | S. epidermidis | 2A | BAS | UCI | Sí |
| 7 | S. epidermidis | 2A | Catéter | UCI | Sí |
| 8 | S. epidermidis | 2B | Catéter | REA | No |
| 9 | S. epidermidis | 2B | HC | UCI | Sí |
| 10 | S. epidermidis | 2B | Catéter | UCI | No |
| 11 | S. epidermidis | 2B | Pus | REA | Sí |
| 12 | S. epidermidis | 2C | Pus | Endocrino | Sí |
| 13 | S. epidermidis | 2D | Pus | REA | Sí |
| 14 | S. epidermidis | 2D | HC | MIN | No |
| 15 | S. hominis | HC | REA | Sí | |
| 16 | S. epidermidis | 2C | HC | MIN | Sí |
| 17 | S. epidermidis | 2C | HC | Urgencias | Sí |
| 18 | S. epidermidis | 2A | HC | MIN | Sí |
| 19 | S. hominis | HC | UCI | Sí | |
| 20 | S. epidermidis | 2A | HC | MIN | Sí |
| 21 | S. epidermidis | 3 | HC | CS | Sí |
| 22 | S. epidermidis | 2B | HC | REA | Sí |
BAS: broncoaspirado; CS: centro de salud; HC: hemocultivo; MIN: medicina interna; REA: unidad de reanimación; UCI: unidad de cuidados intensivos.
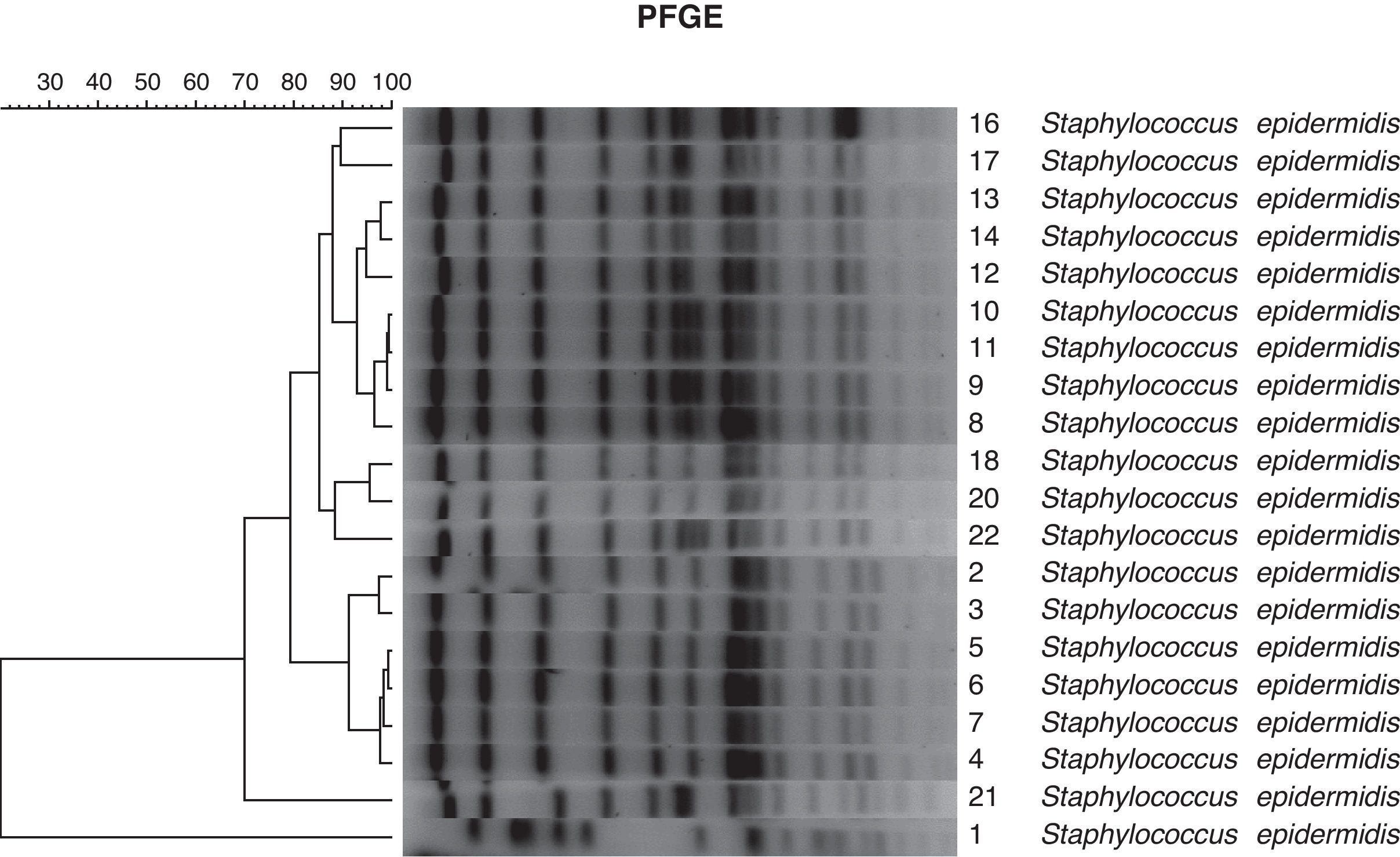
Según el dendrograma (fig. 1) obtenido al analizar el gel de electroforesis en campo pulsado de las 20 cepas de S. epidermidis, se obtuvieron los siguientes resultados:
- -
El perfil de la cepa n.° 1 es completamente diferente del resto de las cepas estudiadas.
- -
El perfil de la cepa n.° 21 también es diferente, pero más cercano (70% de similitud).
- -
El cluster formado por las cepas 2, 3, 4, 5, 6 y 7 presenta un alto grado de similitud (92%).
.")
Asimismo, también tienen alto grado de similitud (90%) las cepas 8, 9, 10, 11, 12, 13, 14, 16 y 17, cercanamente relacionadas (85%) al grupo formado por las cepas números 18, 20 y 22.
Los 2 últimos grupos están posiblemente relacionados (80% de similitud) con el primer cluster (tabla 1).
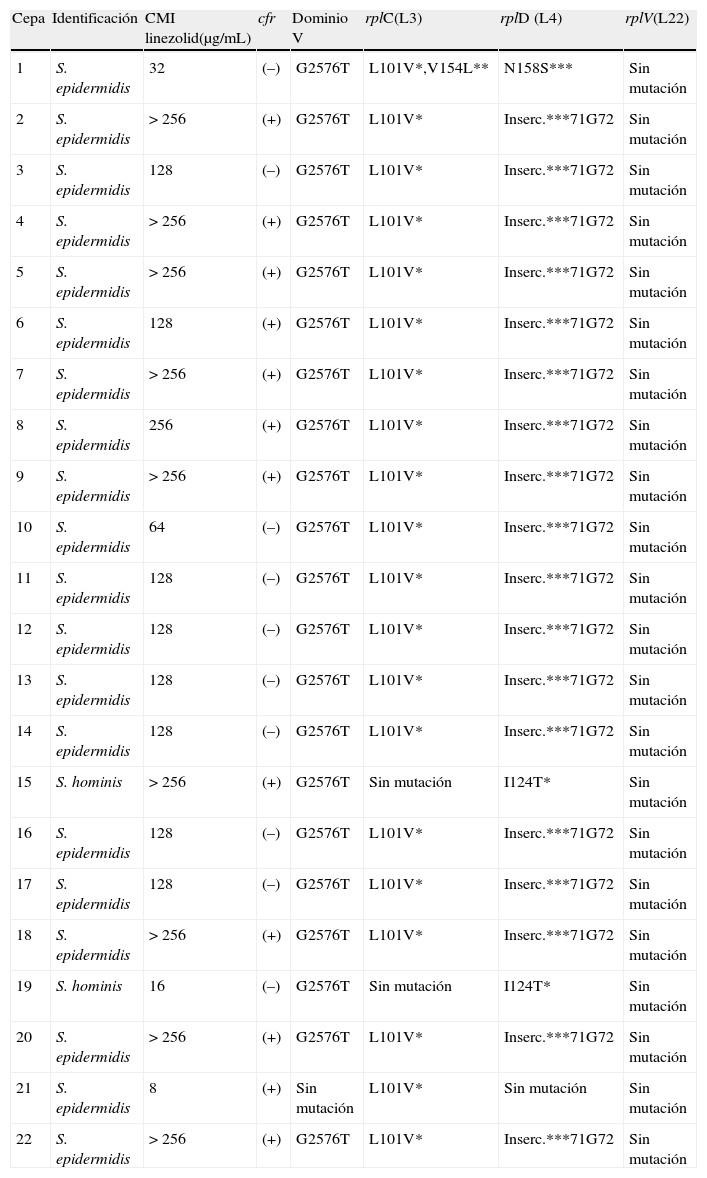
En el 54,5% de los aislados se detectó la presencia del gen cfr, correspondiéndose en el 95,5% de los casos con las cepas que presentaron un valor más alto de concentración mínima inhibitoria (CMI) a linezolid. Veintiuno de los 22 aislados presentaron la mutación G2576T en el dominio V del gen ARNr 23S. En ninguna de las cepas se objetivó la mutación en el gen rplV (L22) (tabla 2).
CMI de linezolid y mecanismos de resistencia asociados en las cepas estudiadas
| Cepa | Identificación | CMI linezolid(μg/mL) | cfr | Dominio V | rplC(L3) | rplD (L4) | rplV(L22) |
| 1 | S. epidermidis | 32 | (–) | G2576T | L101V*,V154L** | N158S*** | Sin mutación |
| 2 | S. epidermidis | > 256 | (+) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 3 | S. epidermidis | 128 | (–) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 4 | S. epidermidis | > 256 | (+) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 5 | S. epidermidis | > 256 | (+) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 6 | S. epidermidis | 128 | (+) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 7 | S. epidermidis | > 256 | (+) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 8 | S. epidermidis | 256 | (+) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 9 | S. epidermidis | > 256 | (+) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 10 | S. epidermidis | 64 | (–) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 11 | S. epidermidis | 128 | (–) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 12 | S. epidermidis | 128 | (–) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 13 | S. epidermidis | 128 | (–) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 14 | S. epidermidis | 128 | (–) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 15 | S. hominis | > 256 | (+) | G2576T | Sin mutación | I124T* | Sin mutación |
| 16 | S. epidermidis | 128 | (–) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 17 | S. epidermidis | 128 | (–) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 18 | S. epidermidis | > 256 | (+) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 19 | S. hominis | 16 | (–) | G2576T | Sin mutación | I124T* | Sin mutación |
| 20 | S. epidermidis | > 256 | (+) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
| 21 | S. epidermidis | 8 | (+) | Sin mutación | L101V* | Sin mutación | Sin mutación |
| 22 | S. epidermidis | > 256 | (+) | G2576T | L101V* | Inserc.***71G72 | Sin mutación |
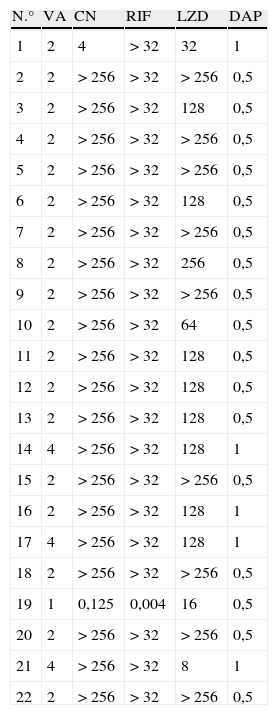
Mediante microdilución, todos los aislados fueron resistentes a la oxacilina, el levofloxacino y el cotrimoxazol. El 90,9% (20 cepas) presentaron resistencia a la gentamicina y el 95,5% (21 cepas) a la rifampicina. Veintiuna de las 22 cepas presentaron CMI≥2μg/ml a la vancomicina, y en 3 de ellas la CMI de este antimicrobiano fue de 4μg/ml. Todos los aislados fueron sensibles a la daptomicina, con valores de CMI entre 0,5 y 1μg/ml. El rango de CMI para el linezolid fue de 8 a >256μg/ml (tabla 3).
CMI del linezolid, la vancomicina, la gentamicina, la rifampicina y la daptomicina mediante microdilución
| N.° | VA | CN | RIF | LZD | DAP |
| 1 | 2 | 4 | > 32 | 32 | 1 |
| 2 | 2 | > 256 | > 32 | > 256 | 0,5 |
| 3 | 2 | > 256 | > 32 | 128 | 0,5 |
| 4 | 2 | > 256 | > 32 | > 256 | 0,5 |
| 5 | 2 | > 256 | > 32 | > 256 | 0,5 |
| 6 | 2 | > 256 | > 32 | 128 | 0,5 |
| 7 | 2 | > 256 | > 32 | > 256 | 0,5 |
| 8 | 2 | > 256 | > 32 | 256 | 0,5 |
| 9 | 2 | > 256 | > 32 | > 256 | 0,5 |
| 10 | 2 | > 256 | > 32 | 64 | 0,5 |
| 11 | 2 | > 256 | > 32 | 128 | 0,5 |
| 12 | 2 | > 256 | > 32 | 128 | 0,5 |
| 13 | 2 | > 256 | > 32 | 128 | 0,5 |
| 14 | 4 | > 256 | > 32 | 128 | 1 |
| 15 | 2 | > 256 | > 32 | > 256 | 0,5 |
| 16 | 2 | > 256 | > 32 | 128 | 1 |
| 17 | 4 | > 256 | > 32 | 128 | 1 |
| 18 | 2 | > 256 | > 32 | > 256 | 0,5 |
| 19 | 1 | 0,125 | 0,004 | 16 | 0,5 |
| 20 | 2 | > 256 | > 32 | > 256 | 0,5 |
| 21 | 4 | > 256 | > 32 | 8 | 1 |
| 22 | 2 | > 256 | > 32 | > 256 | 0,5 |
CN: gentamicina; DAP: daptomicina; LZD: linezolid; RIF; rifampicina; VA: vancomicina.
Entre las combinaciones antibióticas ensayadas, la más efectiva fue linezolid más gentamicina (LZ+CN), que presentó efecto sinérgico en 10 de los 22 aislados (45,5%) y aditivo en 3 aislados (13,6%).
Otras combinaciones, linezolid más rifampicina (LZ+RIF) y linezolid más vancomicina (LZ+VA), no presentaron efecto sinérgico en ningún caso.
De los aislados en los que se observó efecto sinérgico, solamente se objetivó la presencia del gen cfr en 2 casos (tabla 4).
Combinaciones de linezolid y gentamicina y efectos sinérgicos observados en las 22 cepas estudiadas
| Cepa | Identificación | cfr | CMI linezolid | Efecto |
| 1 | S. epidermidis | (–) | 32 | Aditivo |
| 2 | S. epidermidis | (+) | > 256 | Indiferente |
| 3 | S. epidermidis | (–) | 128 | Sinérgico |
| 4 | S. epidermidis | (+) | > 256 | Indiferente |
| 5 | S. epidermidis | (+) | > 256 | Aditivo |
| 6 | S. epidermidis | (+) | 128 | Sinérgico |
| 7 | S. epidermidis | (+) | > 256 | Indiferente |
| 8 | S. epidermidis | (+) | 256 | Indiferente |
| 9 | S. epidermidis | (+) | > 256 | Indiferente |
| 10 | S. epidermidis | (–) | 64 | Aditivo |
| 11 | S. epidermidis | (–) | 128 | Sinérgico |
| 12 | S. epidermidis | (–) | 128 | Sinérgico |
| 13 | S. epidermidis | (–) | 128 | Sinérgico |
| 14 | S. epidermidis | (–) | 128 | Sinérgico |
| 15 | S. hominis | (+) | > 256 | Indiferente |
| 16 | S. epidermidis | (–) | 128 | Sinérgico |
| 17 | S. epidermidis | (–) | 128 | Sinérgico |
| 18 | S. epidermidis | (+) | > 256 | Indiferente |
| 19 | S. hominis | (–) | 16 | Sinérgico |
| 20 | S. epidermidis | (+) | > 256 | Indiferente |
| 21 | S. epidermidis | (+) | 8 | Sinérgico |
| 22 | S. epidermidis | (+) | > 256 | Indiferente |
La resistencia al linezolid entre los aislados clínicos de estafilococos, que en un principio aparecía puntualmente y de forma anecdótica, ha ido aumentando tanto a nivel nacional como internacional. Aunque la mayor atención se ha dirigido fundamentalmente a las cepas de S. aureus, la presencia de un número cada vez mayor de SCN con estas características nos obliga a tener en cuenta la implicación de estos últimos microorganismos, capaces de ocasionar infecciones en pacientes inmunocomprometidos o con prótesis u otros dispositivos.
El programa LEADER de vigilancia de resistencia al linezolid en EE.UU. informa de un incremento en el número de aislamientos de estafilococos coagulasa negativos desde el 0,2% en 2004 al 1,63% en 200815. En nuestro país, la emergencia de este tipo de cepas resistentes en los centros sanitarios se está convirtiendo en un problema importante, asociado, en algún caso, a brotes epidémicos16.
Entre nuestras cepas, S. epidermidis fue la especie de SCN más frecuentemente encontrada. De las 20 cepas estudiadas, 12 presentaban un alto grado de similitud, cercano al 90%, en su perfil electroforético.
La mayoría de los aislamientos procedían de hemocultivos y cultivos de catéteres obtenidos de pacientes ingresados en unidades de cuidados intensivos. En algunos casos estos aislamientos se asociaron a bacteriemias e infecciones del catéter y en casi todos ellos constaba el antecedente de tratamiento previo con linezolid.
El mecanismo de resistencia más común en las cepas estudiadas fue la mutación G2576T en el dominio V. Por el contrario, en ninguna cepa se observó la mutación en el gen rplV (L22). En 12 cepas se detectó el gen cfr, asociado en la gran mayoría de los casos a los valores más altos de CMI de linezolid.
Las cepas de SCN-RL incluidas en este estudio presentan, en el 95% de los casos, una CMI de vancomicina ≥2μg/ml. La presencia de estos aislamientos con CMI elevadas, aunque dentro del intervalo de sensibilidad, podría asociarse a una peor respuesta al tratamiento con este antimicrobiano17. La daptomicina se ha mostrado muy efectiva, y su actividad no se ha visto afectada por la resistencia a otros antimicrobianos.
La valoración de las combinaciones de linezolid con otros agentes antiestafilocócicos es escasa y los resultados contradictorios, dependiendo del tipo de antibiótico asociado y del método utilizado para su determinación. Pero aun así, empleando métodos iguales y con la misma combinación, los resultados no son ni mucho menos uniformes18. Además estos resultados in vitro no se corresponden, en ocasiones, con los obtenidos a partir de modelos animales19. Todo ello produce una falta de criterio a la hora de valorar las combinaciones de linezolid con otras moléculas, generando numerosas incógnitas al respecto.
Por otra parte, todos los estudios referidos anteriormente parten de microorganismos sensibles al linezolid en un intento de optimizar resultados con la asociación con otros antimicrobianos. Sin embargo, otro aspecto a explorar con las combinaciones antibióticas consiste en el rescate de cepas resistentes mediante la acción conjunta de la asociación.
En nuestra serie y partiendo de cepas de Staphylococcus coagulasa negativos resistentes al linezolid, observamos un efecto sinérgico en buena parte de ellas con la combinación de linezolid+gentamicina, hecho que no se observaba con otras combinaciones como linezolid+rifampicina o linezolid+vancomicina. Este efecto sinérgico se observó fundamentalmente en las cepas que no presentaron el gen cfr.
Aunque el linezolid y la gentamicina tienen al ribosoma como diana común, cada uno de ellos actúa en diferentes niveles del ciclo ribosomal. Concretamente, mientras el linezolid se fija a la fracción 23S de la unidad 50S del ribosoma impidiendo la iniciación del complejo 70S, la acción del aminoglucósido es mucho más rápida para ligarse de forma irreversible al ribosoma e impide desde sus inicios la síntesis proteica. El diferente estadio temporal en la actividad de ambos compuestos podría explicar el antagonismo descrito por otros autores19 en la combinación frente a cepas sensibles al linezolid, pero tal vez permitiría una acción secuencial en cepas resistentes a la oxazolidinona, optimizando los resultados. Estas valoraciones provisionales necesitarían confirmarse empleando otras técnicas in vitro distintas al E-test, como la utilización de curvas de muerte o la valoración de concentraciones tipo tablero de ajedrez, así como el estudio de simulaciones farmacocinéticas y modelos experimentales.
Finalmente, podemos concluir que, aunque el número de cepas estudiadas es pequeño, la mayoría de ellas se corresponden a S. epidermidis con un alto grado de proximidad, aparecen en unidades de pacientes críticos que han sido sometidos a tratamientos prolongados con linezolid y que suelen albergar un doble mecanismo de resistencia, mutacional y enzimático. En cuanto a su sensibilidad, suelen presentar valores elevados de CMI de otros antiestafilocócicos como la vancomicina, la rifampicina o la gentamicina. Por último, la asociación de linezolid y gentamicina podría actuar sinérgicamente en algunas de estas cepas.
Conflicto de interesesLos autores declaran que no existe ningún conflicto de intereses.

