









Auditory verbal hallucinations (AVH) as well as aberrant language functioning, i.e. deficits in verbal fluency (VF) are common symptoms in patients with schizophrenia. The relationship between deficits in VF and AVH is rarely studied. In previous imaging studies, activation patterns during VF differed between patients with schizophrenia and healthy controls, independent of AVH phenomenology. However, activation patterns were not investigated for patients with and without AVH in separate groups. We hypothesized that there would be a difference in activation patterns between healthy controls and patients with schizophrenia, but not between hallucinating and non-hallucinating patients.
MethodsIn the current study, we included 31 participants in three groups: patients with schizophrenia and AVH, patients with schizophrenia without AVH, and healthy controls without AVH. All subjects performed a VF task while functional activation was measured by blood-oxygen-level-dependent (BOLD) magnetic resonance imaging (MRI). Task-dependent activation was compared between the three groups.
Results and ConclusionsAll three groups showed activation in language areas of the brain, such as Broca’s area, located in the inferior frontal gyrus, or Wernicke’s area, located in the superior temporal gyrus. Patients had significant task-specific activation in the right hemisphere in addition to activation in language-related areas in the left hemisphere. Unexpectedly, we found significantly more activation in the bilateral fusiform gyrus of non-hallucinating compared to hallucinating patients. It seemed that fusiform gyrus activation serves as a strategy to compensate for performance deficits, significantly more in patients without AVH than in those with AVH.
Schizophrenia is a heterogeneous disorder with a variety of symptoms. Typical symptoms include delusions and auditory verbal hallucinations (AVH), as well as other emotional, cognitive, and motor deficits. A common finding in the cognitive domain, which comprises language function, is poor performance on tasks assessing verbal fluency (VF).1–3 Functional imaging during VF in healthy controls revealed activation in regions responsible for language processing, including the left inferior frontal gyrus (corresponding to Broca’s area), left inferior parietal regions (corresponding to Wernicke’s area), premotor cortex, anterior cingulate cortex, and cerebellum.4–9 Functional imaging during VF in schizophrenia patients revealed aberrant activation patterns, e.g. a reduction in left-lateralized processing of linguistic information.10–14 This decrease in language lateralization is hypothesized to be a consequence of increased right hemisphere engagement in individuals with schizophrenia.10 Decreased language lateralization has also been found in untreated first-episode psychosis patients and non-psychotic twins of schizophrenia patients. 13–15 Furthermore, some studies report a correlation between these aberrant functional activation patterns and AVH severity.10,13 However, results on this relation are inconsistent.14 When taking a closer look at the participants from these studies, we find that either hallucinating and non-hallucinating patients with schizophrenia were grouped together or solely hallucinating patients were included. In another investigation on lateralization, Diederen and colleagues included healthy individuals who presented with isolated AVH and compared their functional activation to that of patients with schizophrenia and AVH, and to functional activation of healthy subjects without AVH.16 The researchers found significant differences in functional activation between healthy controls with AVH and patients with schizophrenia. Further, the activation patterns between both healthy groups (with and without hallucinations) were not significantly different and thus were independent of AVH phenomenology. Diederen at al.16 concluded that activation patterns during VF differ between healthy and psychotic individuals independent of AVH manifestation. However, the findings from this study are not completely conclusive, as no study has included a population of non-hallucinating patients with schizophrenia.
By including a non-hallucinating schizophrenia group in the current study, we aim to better understand the relationship between aberrant functional activation, schizophrenia, and AVH symptoms. To our knowledge, we are the first to investigate differences in functional activation patterns during VF in patients with schizophrenia both with and without AVH. We therefore included participants with schizophrenia and AVH (AH), participants with schizophrenia without AVH (NH), and healthy controls without AVH (C).
Based on results from earlier studies, we expect to see a difference in activation patterns between healthy controls and patients with schizophrenia. Specifically, we expect to see right hemispheric and left hemispheric language-related activation during VF performance in patients with schizophrenia, but not controls. Following the rationale of Diederen et al.16 that psychosis, but not AVH, causes aberrant activation patterns, we do not expect to see different activation patterns in patients with schizophrenia with and without AVH.
Material and MethodsParticipantsWe recruited a total of 31 participants: 12 patients with schizophrenia prone to AVH (AH), 8 patients with schizophrenia not prone to AVH (NH), and 11 healthy controls (C). All patients fulfilled the ICD-10 criteria for schizophrenia or schizoaffective disorder for at least 5 years. General psychopathology was assessed in a clinical interview using the Positive and Negative Syndrome Scale (PANSS).17
Patients in the AH group could (but did not have to) perceive AVH at the time of MRI measurement. According to anamnesis (clinical interview and clinical history files), these patients perceived AVH in at least 90% of the acute phases of the illness. Patients in the NH group neither perceived AVH at the time of MRI measurement nor in the previous 12 months. According to anamnesis, they perceived AVH in less than 10% of the acute phases of the illness. Healthy controls did not have any history of neurological, psychiatric, or somatic medical disorders.
All participants were between 18-60 years of age (see Table 1) and right-handed. They did not use substances other than nicotine and did not have contraindications for magnetic resonance imaging (MRI). All subjects provided written consent after the nature and purpose of the study was fully explained. Further, the study was approved by the local ethics committee (Kantonale Ethikkommission Bern) and performed in accordance with the Declaration of Helsinki.
Subject characteristics of patients with schizophrenia and auditory verbal hallucinations (AH), patients with schizophrenia without auditory verbal hallucinations (NH), and healthy controls (C).
| AH (n = 12) | NH (n = 8) | C (n = 11) | Statistics | |
|---|---|---|---|---|
| M (SD) | M (SD) | M (SD) | ||
| Female / Male | 5/7 | 1/7 | 4/7 | F = 0.97, p = 0.39 |
| Age (years) | 37.8 (8.3) | 38.3 (13.9) | 38.4 (10.0) | F = 0.01, p = 0.99 |
| Age of onset (years) | 25.9 (7.8) | 26.3 (7.0) | Z = -0.47, p = 0.64 | |
| Duration of illness (months) | 161.0 (80.3) | 157.5 (127.2) | t(18) = 0.08, p = 0.94 | |
| Chlorpromazine equivalents (mg) | 648.19 (540.8) | 2389.2 (4110.7) | Z = -1.85, p = 0.07 | |
| PANSS Total score | 67.42 (19.0) | 53.38 (8.5) | Z = -2.01, p = 0.049* | |
| PANSS Positive score | 22.33 (6.0) | 13.50 (3.2) | t(18) = 3.83, p = 0.003* | |
| PANSS Negative score | 16.25 (8.0) | 16.75 (4.5) | Z = -0.93, p = 0.35 |
Results are reported for a level of p = 0.05. Abbreviations: * = significant result, mg = milligrams, PANSS = Positive and Negative Syndrome Scale.
MRI measurements were performed on a 3 T Siemens Trio Scanner using a 12-channel receive-only head coil. For fMRI, 35 axial blood-oxygen-level-dependent (BOLD)-slices parallel to the plane crossing anterior and posterior commissures were obtained (slice thickness 3 mm, inter slice distance 0.75 mm). A 2-D echo planar imaging sequence (repetition time (TR) 2.66 s, echo time (TE) 30 ms, flip angle (FA) 90°, field of view (FOV) 192 × 192 mm2, matrix 64 × 64 pixel resolution, voxel size 3 × 3x3 mm3) was used. Additionally, we acquired 176 sagittal slices by a 3-D gradient modified driven equilibrium Fourier transform sequence (MDEFT)18 with TR 7.92 ms, TE 2.48 ms, inversion time (TI) 910 ms, FA 16°, FOV 256 × 224 mm2, matrix 256 × 192 pixel resolution, and voxel size 1 × 1x1 mm3.
For functional imaging, a verbal fluency paradigm was used. Visual stimuli were presented on an LCD monitor eyeglass (Resonance Technology, Inc., Northridge, CA, USA). In the VF condition, a letter was presented and participants were asked to silently generate as many words as possible starting with the presented letter. In the rest condition (R), a fixation cross was presented and participants were asked to relax. Blocks of VF and R conditions were presented during 10 alternating scans, always starting with the R condition. Altogether, 80 scans were obtained.
Statistical AnalysisBehavioral data was assessed using SPSS software (version 24, IBM Corp., Armonk, NY, USA). PANSS Positive, Negative, and Total scores were tested for normal distribution and were then compared between patient groups by the appropriate parametric or non-parametric test. Statistical significance was set at α = 0.05 for all analyses.
Statistical analyses for MRI data were performed using Matlab software (Version 8.5, release 2015a; The MathWorks, Inc., Natwick, MA, USA), the toolbox SPM12 (Statistical Parametric Mapping; Wellcome Trust Centre for Imaging, London, England), and the toolbox SnPM13 (Statistical nonParametric Mapping; http://warwick.ac.uk/snpm).
Preprocessing of fMRI data included 3-D motion detection and spatial smoothing with a Gaussian Kernel at 8 mm full width at half maximum (FWHM). Two-D functional images were coregistered to 3-D structural images. Functional and structural images were transformed to Montreal Neurological Institute (MNI) space. The BOLD signal was modelled by a boxcar function convolved with a double-gamma hemodynamic response function to create a design matrix for a general linear model (GLM). The GLM included the 3-D motion parameters as covariates. Differences between the VF and R conditions were computed using one-sample t-tests.
For statistical analysis of functional magnetic resonance imaging (fMRI) data, we used a non-parametric approach, as suggested by Nichols and Holmes.19,20 In particular, t-statistic images were computed using the non-parametric statistical toolbox SnPM. These whole-brain contrasts were fed into analyses, performing a non-parametric one-way analysis of variances (ANOVA) with 5’000 permutations. Due to small sample size, variance smoothing with an 8 mm FWMH Gaussian Kernel was applied to obtain “pseudo t-images”. Post hoc tests included non-parametric two-sample t-tests to investigate group differences and non-parametric one-sample t-tests for group-specific activation. Statistical significance was set at non-parametric p = 0.001 and only clusters larger than 4 voxels were reported. Visualizations were performed using the program MRIcroGL (http://www.mccauslandcenter.sc.edu/mricrogl).
ResultsBehavioral dataDemographic characteristics of all subjects are displayed in Table 1. PANSS Negative and Total scores were not normally distributed, thus we used a Mann-Whitney-U-test to investigate group differences. PANSS Positive scores were distributed normally and thus tested for group differences using a two-sample t-test. PANSS Total and Positive scores were significantly higher in the AH than the NH group. There was no significant difference in PANSS Negative scores between patients.
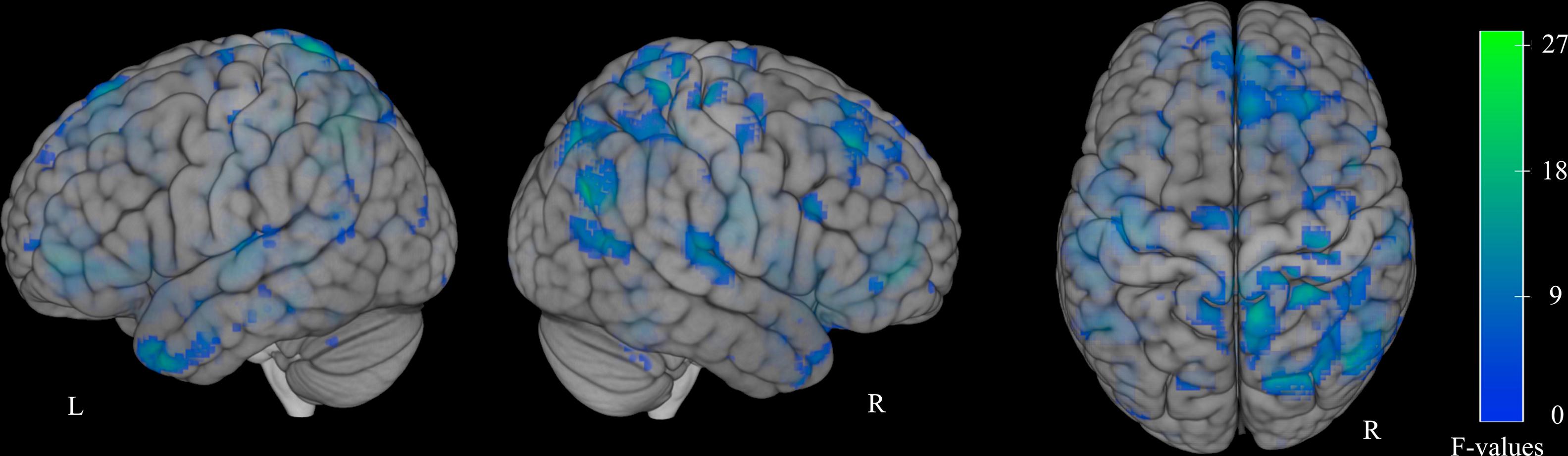
Functional MRI dataUsing a one-way ANOVA with factor “group” (AH, NH, C), we found activation differences in various brain regions, including the bilateral inferior, middle, and superior frontal gyri; anterior cingulate; middle and superior temporal gyri; right inferior parietal lobule; precentral gyrus; insula; and inferior operculum (Supplementary Fig. 1, Supplementary Table 1).
Post hoc two-sample t-tests revealed significant activation differences in clusters in the inferior temporal and fusiform gyrus (FG) bilaterally that were less activated in the AH than NH group (Fig. 1, Table 2).

Voxel statistics and location of significant clusters’ maxima for two-sample t-test with regions more activated for NH compared to AH.
| Cluster size | Statistics | MNI coordinates | Brain regions | |||
|---|---|---|---|---|---|---|
| t-value | p-value | x | y | z | ||
| 64 | 4.02 | < 0.001 | 52 | -56 | -24 | Right Cerebellum / Right ITG / FG |
| 3.44 | < 0.001 | 58 | -48 | -16 | Right Cerebellum / Right ITG / FG | |
| 3.12 | < 0.001 | 54 | -56 | -32 | Tuber | |
| 8 | 3.73 | < 0.001 | -50 | -58 | -22 | Left ITG / FG |
Significant clusters larger than 4 voxels are reported for uncorrected non-parametric level of p = 0.001. FG = Fusiform Gyrus, ITG = inferior temporal gyrus.
Further, group comparisons yielded a pattern of widely distributed clusters that were more activated in the AH compared to the C group in the bilateral middle and inferior frontal gyri, middle and superior temporal gyri, insulae, and anterior cingulum as well as Broca’s and Wernicke’s areas in the left hemisphere (Supplementary Fig. 2A, Supplementary Table 2). The activation pattern in the right hemisphere extended over the angular and supramarginal gyri, including Wernicke’s homologue, as well as to the temporal pole.
A comparison between the NH and C groups revealed a wide pattern of higher activation for NH bilaterally in the middle and superior frontal gyri, anterior cingulate, and middle and superior temporal gyri, including Wernicke’s area (Supplementary Fig. 2B). Activation patterns were also evident in language areas’ homologues (Broca’s and Wernicke’s areas) in the right hemisphere. Additional activation in the right hemisphere ranged over the right FG, parahippocampal, and hippocampal regions. There was only one single cluster in the left cuneus significantly less activated in the NH group (not shown).
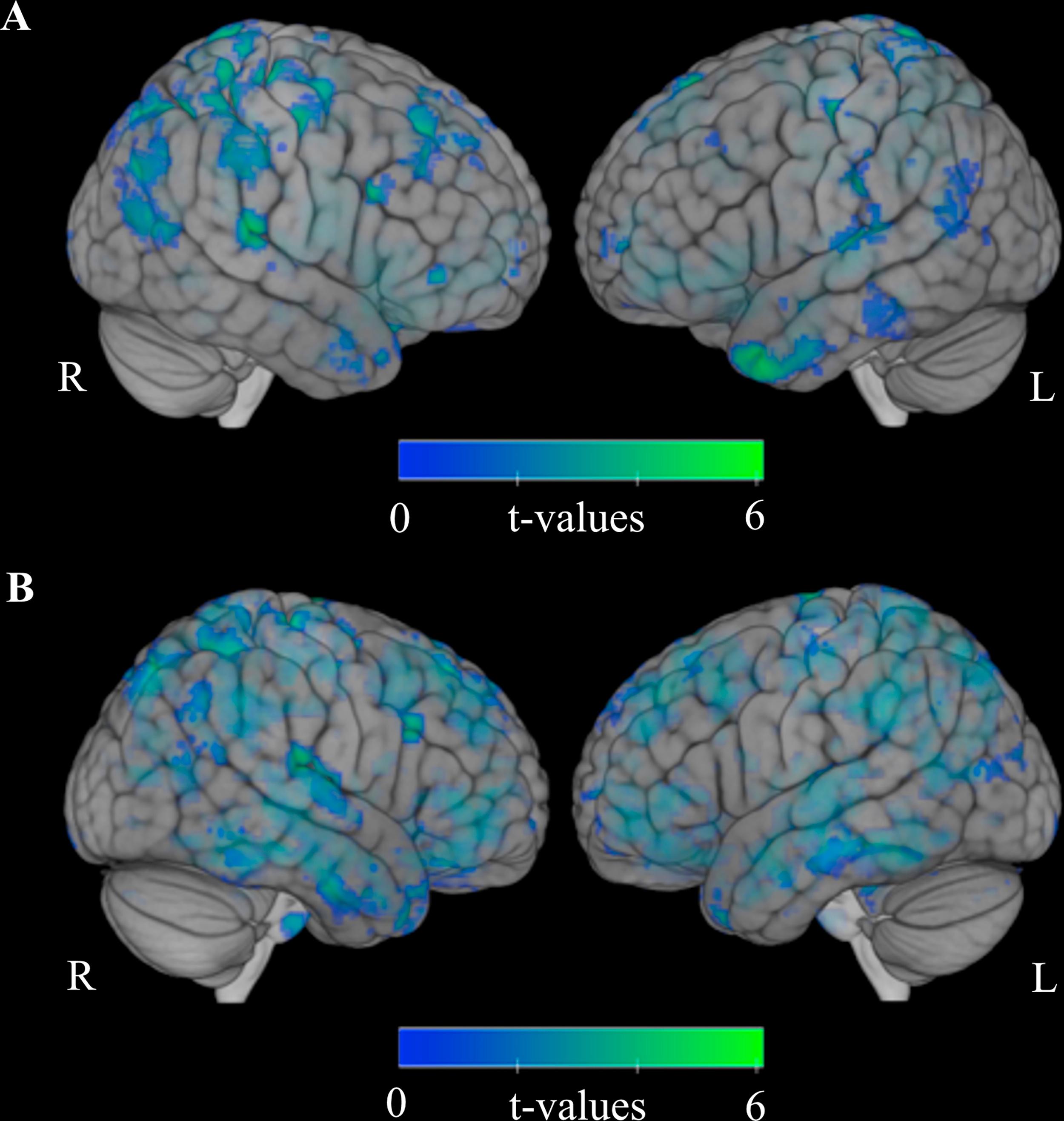
One-sample t-tests were performed to investigate group-specific task-related activation patterns in the VF and R conditions. We found that patients in the AH group had more task-related activation in the bilateral inferior frontal gyrus, insula, and superior temporal gyrus (Broca’s area and its homologue in the right hemisphere), left inferior parietal lobule, supramarginal area (Wernicke’s area), lenticular nucleus as well as activation in the right hemisphere in clusters including the FG, lingual gyrus, and cerebellum, including the declive (Fig. 2A).
 Activation for AH (hallucinating patients with schizophrenia) and (B) activation for C (healthy controls). AH show activation in left-sided Broca’s and Wernicke’s areas whereas C only show activation in right-sided homologue of Broca’s area. Thus, no left-sided activation in language-related areas was found for C. Results for NH (non-hallucinating patients with schizophrenia) not shown due to lack of significant results at threshold of uncorrected non-parametric p = 0.001. Faded coloring resembles activation in deeper lying regions. L = left hemisphere, R= right hemisphere.")
Group-specific task-dependent functional activation compared to rest. (A) Activation for AH (hallucinating patients with schizophrenia) and (B) activation for C (healthy controls). AH show activation in left-sided Broca’s and Wernicke’s areas whereas C only show activation in right-sided homologue of Broca’s area. Thus, no left-sided activation in language-related areas was found for C. Results for NH (non-hallucinating patients with schizophrenia) not shown due to lack of significant results at threshold of uncorrected non-parametric p = 0.001. Faded coloring resembles activation in deeper lying regions. L = left hemisphere, R= right hemisphere.
In NH patients, we did not find any significant activation differences between the VF and R conditions at the uncorrected p-level of non-parametric p = 0.001. We wanted to assure a good quality of data and rule out the possibility that the results are due to non-compliant participants who did not pursue task instructions. Therefore, we applied a more liberal non-parametric p-level. This level was determined as follows: in the non-parametric approach that we used for data analysis, a p-level was calculated for the permutation test in each voxel. The distribution of these p-levels was then employed for significance testing. The lowest uncorrected p-value in this dataset was p < 0.003, thus we applied this threshold to further analyses. At this more liberal significance level, NH activation patterns were significantly different for VF and R conditions with task-related activation in the bilateral inferior and middle frontal gyri (Broca’s area and its homologue in the right hemisphere), left superior parietal lobule and precuneus, and fusiform and middle occipital gyri (not shown). However, the power of this analysis seemed to be too low, either due to the effect or group size, and thus was unable to reach significance at the uncorrected non-parametric value of p = 0.001.
In group C, a one-sample t-test revealed activation differences between the VF and R conditions with task-related activation bilaterally in the insulae; superior, middle and inferior frontal gyri; cingulate gyri; supplementary motor areas; a large cluster extending over the fusiform gyri, lingual gyri, middle occipital gyri and cuneus; and right cerebellum, including the declive (Fig. 2B). Neither Wernicke’s area nor its homologue in the right hemisphere were activated. There was no activation in Broca’s area in the left hemisphere, however, there was some activation in its homologue in the right hemisphere.
DiscussionIn the current study, the brain’s functional modulation during a verbal fluency task was compared between three groups: participants with schizophrenia and AVH (AH), participants with schizophrenia without AVH (NH), and healthy controls without AVH (C). Subjects of all three groups displayed expected activation in brain regions associated with language processing, i.e. Broca’s or Wernicke’s areas. However, we unexpectedly found more bilateral fusiform cortex activation in the NH than the AH group. Our finding of left fusiform activation when performing a VF task is not surprising for patients with psychosis and for those with AVH.11,21,22 Activation in the right FG is usually associated with processing and recognition of faces in an area called the fusiform face area, or FFA.23 In patients with schizophrenia, however, this area does not seem to be highly specialized. Patients utilize the right fusiform cortex during object recognition;24,25 those with AVH during VF.11 Remarkably, we found differences in activation patterns between patients with schizophrenia with and without hallucinations. No other study has compared activation between non-hallucinating and hallucinating patients performing a VF task. McGuire and colleagues observed differences in fusiform activation in patients with and without AVH when performing two covert language tasks.26 Their participants had to either read a word silently (baseline), complete a sentence with a presented adjective and recite the complete sentence silently (inner speech), or imagine an alien voice citing the sentence (auditory verbal imagery). In the inner speech condition, McGuire et al.26 observed a deactivation of the right fusiform area in hallucinating patients when compared to baseline. Furthermore, non-hallucinating patients displayed higher activation than hallucinating patients in the bilateral fusiform cortex during both the inner speech and auditory verbal imagery conditions compared to baseline. Hence, the fusiform cortex seems to play a role in language-related task execution, as patients with and without hallucinations display different activation patterns.
Hallucinating patients engage fusiform areas during task performance less than non-hallucinating patients. One possible interpretation of this phenomenon is that the AH group might be more efficient in task operation than the NH group because they do not require fusiform area engagement to complete a task. Alternatively, patients with schizophrenia may utilize the fusiform cortex to regulate or monitor AVH. Further, AH would regulate AVH to some extent, whereas the NH group would be able to repress AVH completely (with the fusiform cortex as one of several active regions). Following this logic, however, one would expect fusiform cortex activation during AVH perception regardless of any additional task performance. Sommer et al.11 oppose this interpretation, as they observed fusiform cortex activation in patients with AVH during VF performance, but not during AVH perception alone. Nevertheless, the utilization of fusiform regions seems to be crucial to compensate for performance deficits in patients with schizophrenia.
In children, the left FG was demonstrated to be important in auditory semantic processing.27 Balsamo and colleagues showed that FG activation was correlated with higher accuracy in semantic categorization and a tendency towards better performance in verbal fluency. 27 While shown in children, this FG activation is not a usual finding in adults. 27 Higher activation of FG in patients with schizophrenia but not in healthy controls might thus be a sign of an ‘immature’ or less developed language processing.
The NH group utilizes fusiform areas more than the AH group. It would be interesting to examine whether this discrepancy is advantageous for VF performance. In the current study, there is no performance data available because we employed a covert VF task. Further, we are not aware of any other study that has assessed covert VF performance separately in patients with and without AVH. Consequently, the lack of available data makes it difficult to draw a conclusion. Therefore, future investigations of VF performance and their neuronal correlates which compare patients with schizophrenia with and without hallucinations are crucial. Given that patients without hallucinations manage to enhance their performance by a fusiform strategy, these findings necessitate further research to develop treatments which enhance language-related functioning in schizophrenia, e.g. neurofeedback training.
LimitationsAs expected, patients displayed activation in language-related areas in the left hemisphere and in the homologues of these areas. Furthermore, activation in the right hemisphere was observed in a wider range of areas in patients with schizophrenia than in healthy controls. Hence, our results are consistent with previous literature that suggests that patients with schizophrenia have a deficit in left-lateralized language processing.10–14,28,29 It should be noted, however, that activation patterns for healthy controls were not left-lateralized in our study. In particular, group C did not have any activation in Broca’s or Wernicke’s areas in the left hemisphere; only the homologue of Broca’s area was activated. This finding is rather unexpected, as over 90% of the healthy right-handed population is thought to have lateralized language processing, also in covert VF tasks, in the left hemisphere.30,31 One can only speculate the cause of this unexpected finding. The task may have been too easy for healthy subjects and, consequently, activation in the left hemisphere remained low due to highly efficient processing. Another reason for the finding could be a lack in subject compliance resulting in the absence of typical activation in the left hemisphere. Though no conclusion about the reason behind our unexpected finding can be drawn, when interpreting our results, it is imperative to remember that we did not observe the typical left-lateralization in the healthy control group.
To our knowledge, we are the first to report a difference in brain activation during verbal fluency in patients with and without AVH. One could argue that our unexpected finding is a result of liberal thresholding during our statistical analysis. We are aware of our seemingly liberal threshold without correction. However, unlike most other studies, we used a non-parametric approach to analyze the data. The advantage of this method is that it requires few a priori assumptions compared to random field theory based parametric approaches, which are widely used in other studies.20 In the non-parametric approach, statistical values (i.e. t-, F-, and p-values) are determined by the current sample characteristics. Further, statistical values cannot be mapped like in a parametric approach. So, before beginning the analysis, we decided to threshold using p-values. The non-parametric method is exact and relies less on assumptions and approximations than a parametric approach.19 Furthermore, our hypotheses were informed by findings from earlier studies which used a similar threshold, but utilized parametric approaches with no evidence of meeting the underlying assumptions. Finally, further limitations are the small sample size and a difference in medication dose for the patient groups, although not significant, which warrant replication of our study.
Ethical disclosuresProtection of human and animal subjects. The authors declare that the procedures followed were in accordance with the regulations of the relevant clinical research ethics committee and with those of the Code of Ethics of the World Medical Association (Declaration of Helsinki).
Confidentiality of data. The authors declare that they have followed the protocols of their work center on the publication of patient data.
Right to privacy and informed consent. The authors have obtained the written informed consent of the patients or subjects mentioned in the article. The corresponding author is in possession of this document.
Role of the funding sourceThe funding sources had no role in the study design; collection, analysis, and interpretation of data; writing of the report; or decision to submit the paper for publication.
Declarations of interestNone.
The following are Supplementary data to this article:
Supplementary Figure 1: Task-specific activation differences between all groups. Results for one-way ANOVA with factor group, thresholded at non-parametric p = 0.001. Faded coloring resembles activation in deeper lying regions. R= right hemisphere.
Supplementary Figure 2: Task-specific activation patterns in group comparisons. (A) Higher activation for AH (hallucinating patients with schizophrenia) compared to C (healthy controls) and (B) higher activation for NH (non-hallucinating patients with schizophrenia) compared to C (healthy controls). The comparison of AH to NH did not yield any significant effect. For comparison of NH and AH, see Fig. 1. The comparison of C to AH did not yield any significant effect. Only one cluster in the left cuneus was more activated in C compared to NH (not shown). All results are thresholded at uncorrected non-parametric p = 0.001. Faded coloring resembles activation in deeper lying regions. L = left hemisphere, R = right hemisphere.
Address: Arud, Zentren für Suchtmedizin, Schützengasse 31, CH-8001 Zürich, Switzerland.
Address: Center for Psychiatric Neuroscience, The Feinstein Institute for Medical Research, Zucker School of Medicine at Northwell Hofstra University, 350 Community Drive, Manhasset, NY 11030, USA.
{kind=link}
{kind=link}