




Actualmente el único tratamiento efectivo para las enfermedades hepáticas en estadio terminal es el trasplante de hígado. El número de pacientes en lista de espera aumenta considerablemente cada año, dando lugar a una mayor desproporción entre la oferta y la demanda de un hígado sano. El conocimiento y el posible uso de las células madre ha despertado un gran interés en el campo de la hepatología, haciendo de ellas una de las alternativas más prometedoras a corto plazo. La terapia celular hepática permitiría suplir al hígado de células sanas capaces de llevar a cabo las funciones que las células dañadas no son capaces de desarrollar. Observaciones recientes han puesto de manifiesto la capacidad de las células madre de diferenciarse hacia diferentes linajes celulares. La diferenciación hepática de células madre adultas de diversos orígenes ha dado resultados muy prometedores. El tejido adiposo contiene en el individuo adulto un reservorio de células madre capaces de ser inducidas y diferenciadas hacia diferentes estirpes celulares, presentando un elevado grado de plasticidad celular. La abundancia de este tejido y su fácil accesibilidad hacen de él una prometedora fuente de células madre adultas para su uso en terapia celular hepática. Se presenta una revisión de los avances obtenidos en la diferenciación de células madre procedentes del tejido adiposo hacia células de fenotipo hepático y sus posible aplicaciones como un método terapéutico con la finalidad de mantener la función hepática del paciente durante el período de espera hasta recibir el trasplante, o para facilitar la regeneración hepática en casos de fallo hepático fulminante, y para el tratamiento de pacientes con metabolopatías congénitas.
Currently, the only effective treatment for end-stage liver disease is liver transplantation. The number of patients on the waiting list increases considerably each year, giving rise to a wide imbalance between supply and demand for healthy livers. Knowledge of stem cells and their possible use have awakened great interest in the field of hepatology, these cells being one of the most promising short-term alternatives. He-patic stem cell therapy consists of the implantation of healthy cells capable of performing the functions that dama-ged cells are unable to carry out. Recent observations indicate that several stem cells can differentiate into distinct cell lineages. Hepatic differentiation of adult stem cells from several origins has yielded highly promising results. Adipose tissue in adults contains a reservoir of stem cells that can be induced and differentiated into different types of cells, sho-wing a high degree of plasticity. Because of its abundance and easy access, adipose tissue is a promising source of adult stem cells for hepatic stem cell therapy. The present article reviews the progress made in the differentiation of adult stem cells from adipose tissue into cells with hepatic phenotype. We also discuss the potential application of this technique as a therapy for temporary metabolic support in patients with end-stage liver failure awaiting whole organ transplantation, as a method to support liver function and facilitate regeneration of the native liver in cases of fulminant hepatic failure, and as a treatment in patients with genetic metabolic defects in vital liver functions.
El trasplante hepático es, actualmente, el único tratamiento efectivo existente para las enfermedades hepáticas en estadio terminal. A pesar del incremento constante del número de trasplantes hepáticos realizados, el número de pacientes en lista de espera aumenta cada año y, por tanto, también se incrementa el tiempo de espera y el número de pacientes que fallecen sin llegar a ser trasplantados. Para paliar estos problemas y aumentar el número de trasplantes, se han utilizado varias estrategias. Utilizar injertos procedentes de donantes que hace pocos años no se consideraban aceptables, como los donantes de más de 80 años de edad1, la extracción de órganos a donantes en paro cardíaco2, la división del hígado donante (split), que permite obtener injertos para dos receptores3, el trasplante a partir de un donante vivo4, y finalmente el xenotrasplante (el trasplante de un órgano de otra especie animal), que es teóricamente una solución prometedora ante la escasez de órganos. Sin embargo, aún no ha sido posible controlar la respuesta inmunitaria celular en el rechazo de los xenoinjertos y la respuesta inmunitaria humoral. Por tanto, teniendo en cuenta que las enfermedades hepáticas afectan aproximadamente a un 17,5% de la población mundial5, parece justificado plantearse nuevas alternativas al trasplante alogénico de hígado. Actualmente, la terapia celular se considera una prometedora estrategia complementaria al trasplante de órgano sólido. En principio, las células sanas utilizadas deberían suplir las funciones que las células o tejidos lesionados ya no son capaces de llevar a cabo.
La función normal de los tejidos requiere que el índice de la pérdida celular (apoptosis6,7) sea correspondido por el índice de su renovación. Dado el agotamiento celular continuo, que ocurre con el tiempo, cuando la pérdida excede la reparación, lo que sobreviene conduce eventualmente a una disminución en la función y, en última instancia, al fallo del órgano. La finalidad de cualquier tratamiento ante una pérdida de función o fallo orgánico es sustituir o reparar la función de células o tejidos lesionados.
Dentro del área de la hepatología, el trasplante celular debería permitir la corrección de enfermedades hepáticas (metabolopatías hereditarias, hepatopatías virales, hepatopatías tóxicas), así como el mantenimiento de la función hepática en pacientes que se encuentran en lista de espera8–10.
FUENTES CELULARES PARA TERAPIA CELULAR HEPÁTICAEn los últimos años se ha dedicado un gran esfuerzo al desarrollo y la caracterización de modelos celulares que puedan constituir una alternativa a los hepatocitos humanos. Entre las diferentes aproximaciones experimenta-les, cabe destacar el uso de células derivadas de hepatoblastomas naturales, como las HepG2. Sin embargo, estas células carecen de la expresión de funciones hepáticas, articularmente del citocromo P450, sistema enzimático encargado del metabolismo de fármacos en el hígado y, por tanto, no parecen ser una alternativa al uso de hepatocitos11,12. La inmortalización de los hepatocitos humanos es otra opción; sin embargo, a pesar de los avances, las células resultantes no manifiestan los rasgos fenotípicos deseables13–16. El uso de células hepáticas procedentes de otras especies para xenotrasplante es otra posibilidad; en cambio, esta alternativa no está exenta de dificultades, y también arrastra el problema del mantenimiento de un tratamiento inmunosupresor, así como las complicaciones relacionadas con el rechazo17,18.
El trasplante celular a partir de hepatocitos adultos aislados de órganos enteros descartados para implante se ha aplicado con éxito en el caso de determinadas metabolopatías congénitas. Así, se ha descrito la recuperación de las funciones hepáticas, específicamente la función deficitaria por el trastorno metabólico, en casos puntuales (déficit de alfa-1-antitripsina, déficit de UDP-glucuronosiltransferasa o síndrome de Criggler-Najjar tipo I, trastorno del almacenamiento de glucógeno por déficit de glucosa-6-fosfatasa o enfermedad del almacenamiento del glucógeno tipo Ia y en el caso de la enfermedad de Refsum infantil o error del metabolismo de los peroxisomas) 10,19–23. Son muchas las ventajas del trasplante celular hepático frente al trasplante alogénico. Sin embargo, hay limitaciones en la utilización de hepatocitos humanos adultos, ya que se constata un déficit de hígados descartados para trasplante alogénico pero útiles para el aislamiento de hepatocitos viables. Así, los órganos ideales para la obtención de hepatocitos humanos viables son los que se utilizan para el implante de órgano sólido24. En estos momentos, las únicas fuentes de hepatocitos para trasplante celular son los órganos descartados para el implante, el tejido hepático redundante resultante de los procedimientos de reducción hepática para trasplante infantil25, y el tejido sobrante tras los procedimientos de partición hepática (split)26. Dada esta limitación, los problemas a los que nos enfrentamos son los derivados de la calidad y la cantidad del tejido hepático disponible. La esteatosis, principal motivo de exclusión de los injertos para implante, supone un descenso drástico en la capacidad del método de digestión enzimática para la obtención de hepatocitos humanos viables. Hay evidencias recientes de que los hepatocitos esteatósicos no son suficientemente activos desde el punto de vista de su repertorio metabólico para suplir otras funciones deficitarias27,28. Son necesarios nuevos métodos que permitan recuperar los hepatocitos humanos viables a partir de los hígados grasos25. El desfase de tiempo entre el momento de la obtención de hepatocitos (momento de la donación) y su utilización (injerto en un receptor adecuado) hace que se pierda gran cantidad de recursos, debido a la falta de la optimización de los métodos de crioconservación de los hepatocitos29.
La terapia celular basada en la utilización de células madre es una nueva y prometedora alternativa. Estudios recientes realizados en animales indican la utilización de las llamadas células ovales del hígado y células hepáticas troncales, que parecen tener la capacidad de proliferar y diferenciarse a hepatocitos30. Se han utilizado también hepatocitos fetales y células troncales fetales sin demostrar que sean capaces de restaurar la función hepática normal31. Finalmente, despiertan gran interés los precursores pluripotentes como fuente potencial para el trasplante celular a distintos órganos. Un ejemplo de ello es el uso de células mesenquimales adultas derivadas de la médula ósea para la repoblación hepática mostrada en modelos animales32,33.
TERAPIA CELULAR BASADA EN LA UTILIZACIÓN DE CÉLULAS MADRELa investigación con células madre constituye actualmente uno de los campos más fascinantes de la biología moderna, no sólo en términos del entendimiento de los factores que determinan el desarrollo embrionario, sino también en términos de la búsqueda de mecanismos que provocan el reemplazamiento de células dañadas por células sanas en un organismo adulto. Una de las principales características de las células madre es su capacidad de dar lugar a cada uno de los tipos celulares especializados, proceso denominado «diferenciación». En este proceso de diferenciación intervienen tanto las señales internas controladas por genes intracelulares como externas, que incluyen sustancias químicas secretadas por otras células, el contacto físico de unas células con otras y ciertas moléculas del microentorno.
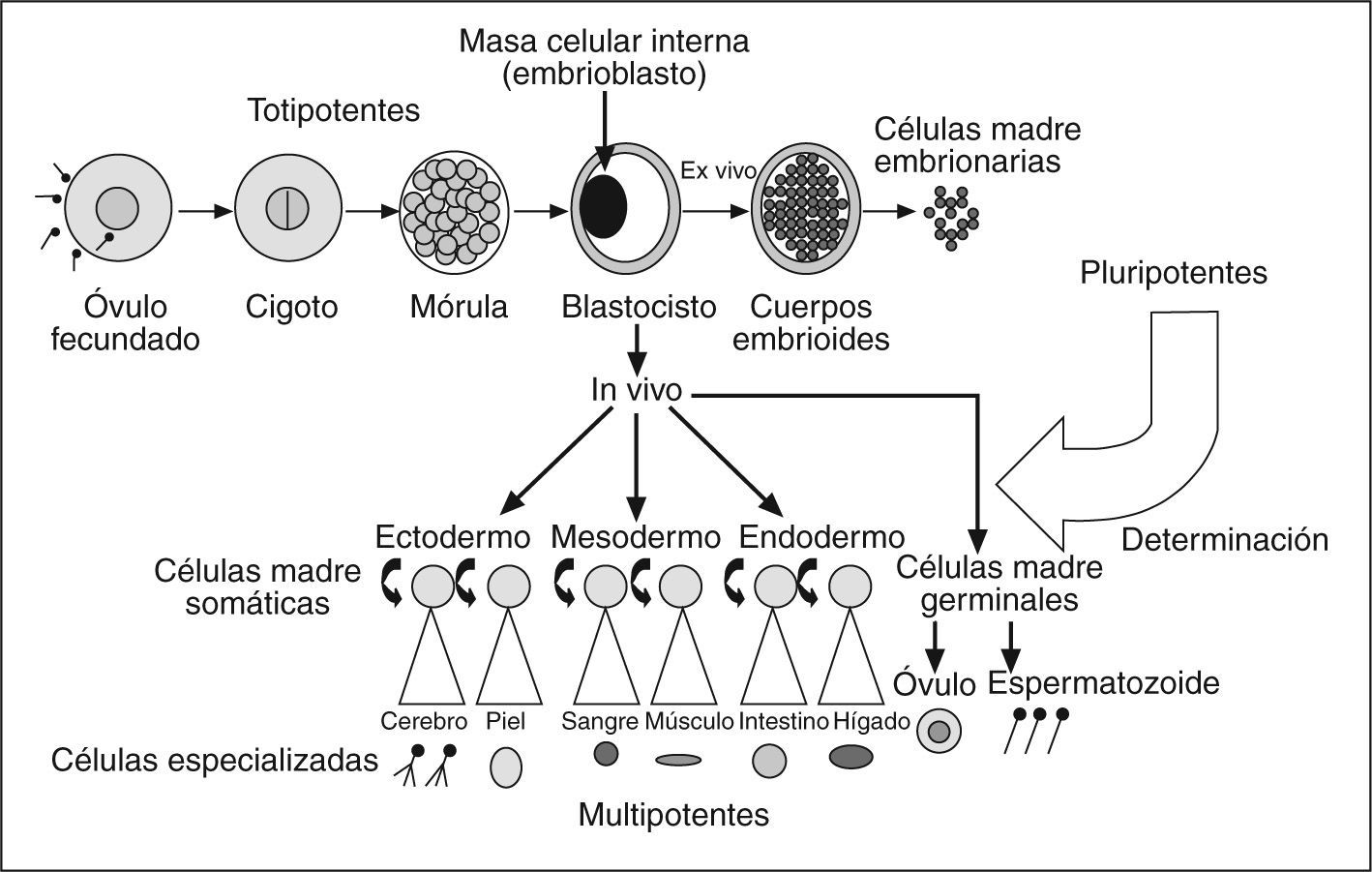
Por célula madre, o troncales, se entiende cualquier célula que tiene la doble capacidad de dividirse ilimitadamente y de dar lugar a diferentes tipos de células especializadas. De acuerdo con esta segunda capacidad, las células troncales pueden ser totipotentes, pluripotentes y multipotentes en relación con su mayor o menor grado o potencial para diferenciarse, tal como se definen a continuación. La totipotencia es la capacidad funcional de una célula para dar lugar a un individuo completo tras un proceso de desarrollo normal. La pluripotencia es la capacidad funcional de una célula para dar lugar a cualquier estirpe celular, pero no a un embrión completo. La multipotencia se define como la capacidad de generar células, capacidad funcional de una célula para dar lugar a alguno, pero no a todos, los linajes celulares. Las células troncales multipotentes están presentes en los tejidos u órganos adultos y tienen una capacidad limitada de reactivar su programa genético en respuesta a determinados estímulos exógenos34. Atendiendo a su origen, hay varias clases de células troncales (embrionarias, germinales embrionarias, adultas). Las células troncales adultas se obtienen de tejidos adultos. Se trata de células indiferenciadas capaces de autorrenovarse para lograr el mantenimiento de una reserva funcional y de diferenciarse hacia tipos celulares, no sólo de su tejido originario sino también hacia otras estirpes celulares35–41. De este modo, las células madre adultas dejan de considerarse multipotentes para ser consideradas células pluripotentes. Este mayor potencial de diferenciación permite hablar en términos de plasticidad celular, capacidad de una célula madre adulta de un tejido específico para dar lugar a un tipo celular especializado diferente del de su origen embrionario41,42 (fig. 1).
")
Embriogénesis humana: potencial y grado de diferenciación. (Modificado de Hernández Ramírez y Dorticós Balea83.)
La médula ósea se considera la fuente de células troncales adultas más prometedora. Se han descrito diferentes tipos de células madre en la médula ósea: hematopoyéticas (HSC)34, mesenquimales (MSC)43, side population cells (SPC)44 y células progenitoras adultas multipotenciales (MAPC)45.
Las MSC, también llamadas células madre estromales, constituyen un modelo muy útil en aplicaciones clínicas para un número de enfermedades, tanto en terapia regenerativa como en terapia génica. En los últimos años se han descrito distintos marcadores de superficie que han permitido identificar y aislar células MSC, como SH2, SH3, CD29, CD44, CD71, CD90, CD105, CD106 y CD120a43,46. Las MSC no expresan antígenos de superficie típicos de las HSC, como CD34, CD45 o CD1443,47. Algunos experimentos recientes han demostrado que las MSC son capaces de diferenciarse in vitro a tejidos mesodérmicos funcionales, como osteoblastos, condroblastos, adipocitos y mioblastos esqueléticos45, y a otros tipos celulares derivados de otras hojas embrionarias, como hepatocitos43,45,48–51.
De forma casi continua aparecen nuevos estudios en los que se aíslan células mesenquimales a partir de tejidos adultos con capacidad multipotencial.
El tejido adiposo es altamente complejo, formado por adipocitos maduros, preadipocitos, fibroblastos, células del músculo liso vascular, células endoteliales, monocitos, macrófagos y linfocitos. La fracción celular de la estroma vascular del tejido adiposo ha despertado recientemente un gran interés en la investigación con células madre52,53. Estudios recientes han identificado una población de células madre en la estroma adiposa (adipose derived stem cells [ADSC])52,53. Se ha demostrado que las células madre derivadas del tejido adiposo tienen características similares a las de la médula ósea in vitro e in vivo, de forma que el tejido adiposo podría ser una fuente ideal de células madre autólogas para el trasplante celular39–41. El tejido adiposo, como la médula ósea, deriva del mesénquima embrionario, pero, a diferencia de ésta, su estroma se puede aislar en gran cantidad con facilidad y con la mínima molestia para el paciente. Estas células poseen mayor capacidad proliferativa que las células mesenquimales de la médula ósea40. Además, son capaces de ser inducidas a transdiferenciarse hacia diferentes tejidos: condrocitos, adipocitos, osteoblastos, miocitos y células endoteliales41,54–56.
En definitiva, poseen ventajas objetivas para constituir una buena fuente alternativa de células adultas autólogas para la terapia celular. El éxito observado en la diferenciación de ADSC en diferentes linajes mesenquimales genera un gran interés en su uso para la diferenciación hepática35–38.
TEJIDO ADIPOSO: FUENTE DE CÉLULAS MADRE MESENQUIMALESNo está claro cuál es el tipo de tejido que da origen a las células adiposas, pero se sabe que todas las células adiposas se originan por un proceso de diferenciación a partir de células mesenquimales primitivas. Hay dos tipos de tejido adiposo, el tejido adiposo blanco, también llamado unilocular, por contener sólo una gota central de lípidos, y el tejido adiposo marrón, o también denominado multilocular, por contener varias gotas de lípidos. El origen de ambos tipos de tejidos no es el mismo. El tejido adiposo blanco comienza a desarrollarse en el quinto mes de vida fetal en los islotes grasos. El proceso se inicia con la aparición en el tejido conectivo de invaginaciones vasculares de lóbulos de adipocitos. Los fibroblastos incluidos en los lóbulos comienzan a acumular lípidos hasta alcanzar la morfología característica del adipocito maduro57. En las células mesenquimales típicas aparecen gradualmente más gotas grasas haciendo a las células más redondeadas. Finalmente, estas gotas se fusionan en una gran vacuola central haciendo al núcleo cada vez más excéntrico. Durante el desarrollo del tejido adiposo podemos hablar de adipoblasto, célula pluripotencial que se replica activamente y que puede dar lugar al preadipocito, célula predestinada a diferenciarse hasta llegar a ser adipocito y, finalmente, adipocito, célula de morfología típica madura con la correspondiente maquinaria enzimática para la acumulación y movilización de triglicéridos58. Las células adiposas totalmente diferenciadas no sufren más procesos mitóticos y tras el nacimiento sólo se forman nuevas células adiposas a partir de células mesenquimales indiferenciadas. El tejido adiposo marrón se origina a partir de células mesenquimales indiferenciadas, pero el proceso es distinto. Primero las células se asemejan a las epiteliales y el tejido se hace lobulado, empiezan a aparecer gotas de lípidos en las células, convirtiendo al tejido en multilocular.
El tejido adiposo presenta una gran habilidad para llevar a cabo cambios de volumen durante la vida de un individuo. Estos cambios pueden producirse por un crecimiento hipertrófico, es decir, por un aumento de tamaño de cada una de sus células adiposas como consecuencia de la acumulación intracelular de lípidos, o por un crecimiento hiperplásico, resultado de la generación de nuevos adipocitos por división de células mesenquimales indiferenciadas59,60. Se ha observado que estos cambios están mediados por poblaciones celulares progenitoras incluidas en el propio tejido adiposo, concretamente en la fracción celular comprendida en la estroma vascular, fracción denominada células madre (progenitoras) derivadas de tejido adiposo (ADSC). El tejido adiposo presenta un elevado grado de plasticidad. Esta plasticidad se define como la capacidad de una célula madre adulta de un tejido específico para generar un tipo celular especializado diferente del de su origen embrionario. Para poder evaluar la plasticidad in vitro deben cumplirse unos criterios mínimos, como la capacidad de autorrenovación, la diferenciación morfológica y funcional hacia tipos celulares de su origen embrionario y, por lo menos, a un tipo celular de diferente linaje61. Las ADSC son células pluripotenciales que se replican continuamente, su morfología bajo ciertas condiciones de inducción pasa de ser fusiforme a adoptar otras formas características de otros tipos celulares (p. ej., poligonal en el caso de la diferenciación hacia fenotipo hepático), y son capaces de dar lugar a preadipocitos que posteriormente se diferenciarán hasta dar lugar a adipocitos adultos y, finalmente, pueden ser conducidas hacia una diferenciación condrogénica, osteogénica, miogénica, endotelial y, como se ha mostrado recientemente, hepática35-38,41,54–56.
Todas estas propiedades hacen de las ADSC una fuente alternativa de células adultas autólogas para terapia celular hepática. La posibilidad de obtener estas células del propio paciente evitaría los problemas derivados del rechazo y el tratamiento inmunosupresor, ya que se trataría de una terapia autóloga. Todo ello hace de las ADSC una herramienta muy útil en el desarrollo de un modelo celular diferenciado próximo al hepatocito humano adulto, tanto para terapia celular hepática como para los estudios de toxicología clínica.
DIFERENCIACIÓN DE LAS ADSC HACIA FENOTIPO HEPÁTICOAunque el número de estudios realizados sobre la potencial capacidad de las ADSC para diferenciarse a fenotipos hepáticos adultos es todavía muy reducido, los resultados son muy prometedores, ya que las ADSC alcanzan un grado significativo de expresión de funciones propias del hepatocito35–38. La inducción de esta diferenciación in vitro se basa en el desarrollo de protocolos que tratan de reproducir el proceso de desarrollo y diferenciación del hígado, teniendo en cuenta los factores de crecimiento, las citocinas y las hormonas que están implicados.
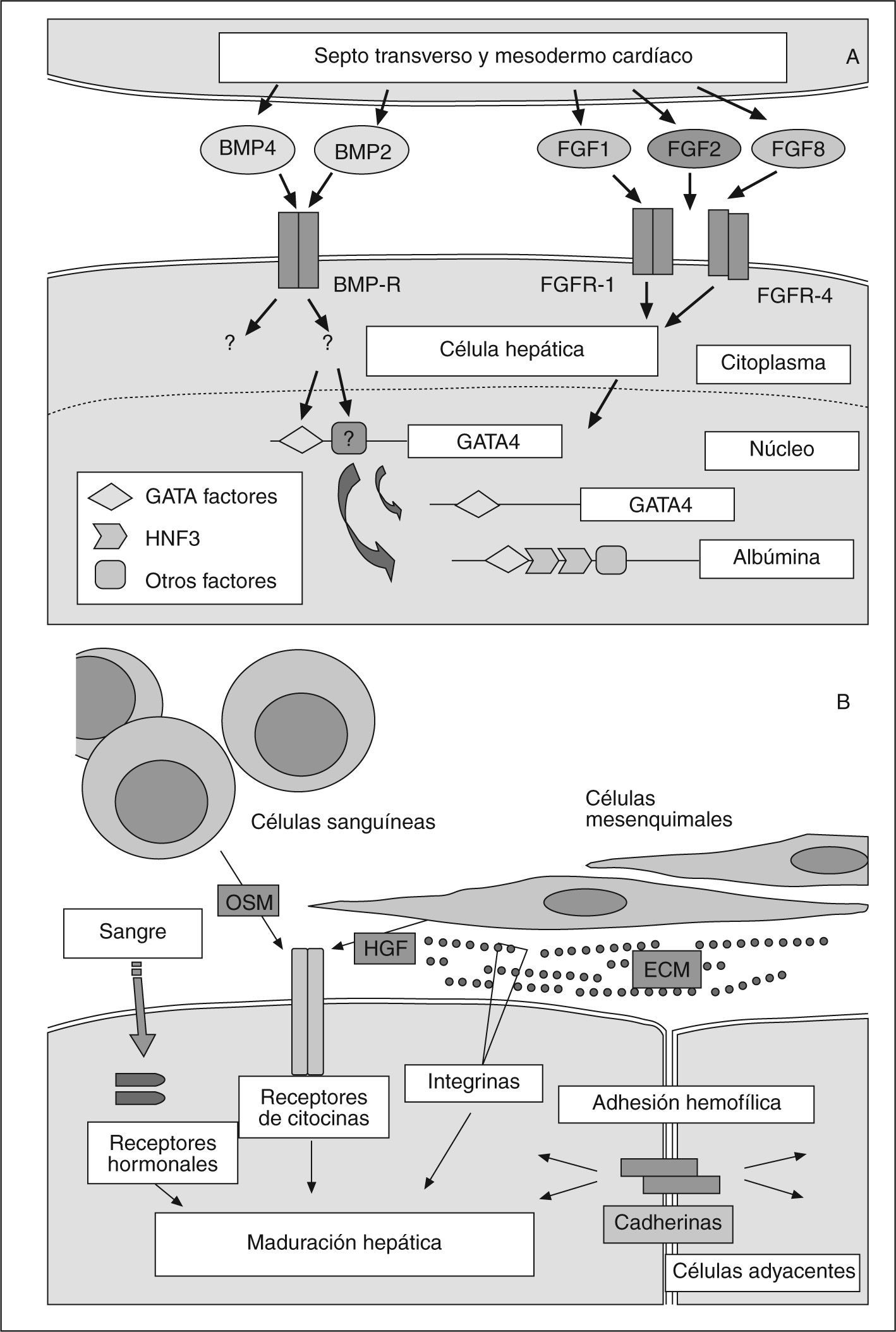
El desarrollo hepático embrionario se produce en múltiples estadios regidos por factores hormonales, así como por interacciones celulares y matriz-celular62. Por tanto, se trata de un proceso gradual condicionado por múltiples señales extracelulares e intracelulares perfectamente ubicadas en el tiempo. El hígado adulto deriva del endodermo63. La especificación hepática a partir del endodermo depende fundamentalmente de dos rutas de señalización63–65. A partir del mesodermo cardíaco se induce una fuerte respuesta morfogenética en el endodermo por medio de factores de crecimiento fibroblásticos (FGF)63–65. Por otra parte, las proteínas morfogenéticas óseas (BMP), secretadas a partir del mesénquima del septo transverso, son necesarias junto con los FGF del mesodermo cardíaco para inducir al endodermo ventral hacia el hígado (hepatoblasto) 63–67 (fig. 2A). En la formación del hígado adulto a partir del hepatoblasto, otros factores de crecimiento tienen su protagonismo. El factor de crecimiento hepático (HGF), involucrado en la regeneración hepática tras una lesión tisular, también regula la expresión de los marcadores de diferenciación hepáticos en presencia de glucocorticoides (p. ej., dexametasona)65,68,69. En la maduración y la diferenciación terminal, desempeñan un papel muy importante la oncostatina (OSM)70–72, los glucocorticoides, una elevada densidad celular69 y la matriz extracelular (ECM)73. La OSM estimula la expresión de marcadores de diferenciación hepáticos, induce cambios morfológicos y regula múltiples funciones específicas del hígado. Los glucocorticoides son fundamentales para la maduración hepática, mientras que la OSM incrementa los efectos de éstos; la OSM sola no induce fenotipos diferenciados, sino que es absolutamente imprescindible la presencia de una concentración fisiológica de glucocorticoides (dexametasona) para que la OSM ejerza sus acciones y efectos; por ello, la inducción de la maduración hepática requiere OSM y dexametasona70. La adherencia celular a la ECM es de vital importancia para el correcto desarrollo tisular. Los componentes moleculares de la matriz extracelular (colágenos, glucoproteínas y proteoglucanos) están físicamente interconectados con las proteínas del citosqueleto, a través de los receptores de membrana, como las integrinas; estas interacciones mecánicas son importantes en la determinación de la forma celular. Los factores proteínicos asociados con la ECM, como el factor de crecimiento epidérmico (EGF) y el factor de crecimiento transformante alfa (TGFα), también están implicados en la diferenciación celular inducida por la matriz. Así pues, la ECM debe actuar sobre la diferenciación por distintas rutas de señalización74,75 (fig. 2B).
")
Diferenciación hepática. A: posible modelo de la especificación temprana hepática. B: rutas promotoras de los estadios terminales del desarrollo hepático. (Modificado de Haynesworth et al46.)
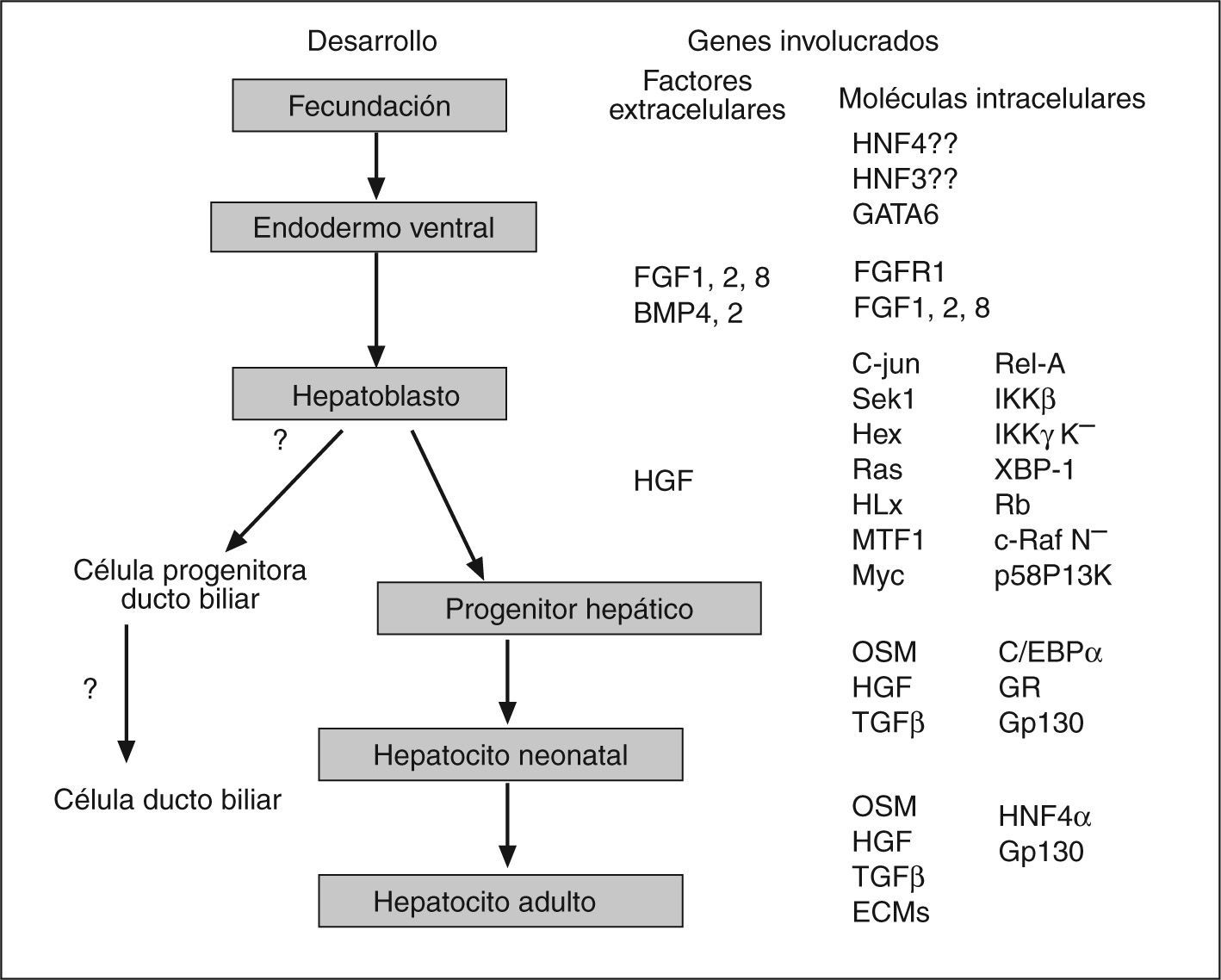
Se sabe que la diferenciación y otros procesos celulares básicos están influidos por interacciones humorales, intercelulares y matriz-celular; sin embargo, el principal mecanismo responsable de la diversidad fenotípica es la expresión de un set de genes específicos para cada tipo celular. La diferente expresión genética entre tipos celulares está determinada, en la mayoría de los casos, por una regulación transcripcional mediada por una combinación específica de factores de transcripción76. Diversos estudios indican que la expresión específica de genes típicamente hepáticos requiere la unión de múltiples y distintos factores de transcripción abundantes en el hígado (liverenriched transcription factor [LETF]) a regiones reguladoras, lo que permite la activación transcripcional de los genes necesarios para llevar a cabo las funciones típicamente hepáticas77. Sin embargo, la regulación de la transcripción no sólo depende de la activación por factores de transcripción, sino que hay una gran cantidad de correguladores que desempeñan un papel clave en muchos procesos biológicos, incluidos los programas de diferenciación celular77,78 (fig. 3).
")
Moléculas extracelulares e intracelulares implicadas en el desarrollo hepático. (Modificado de Haynesworth et al46.)
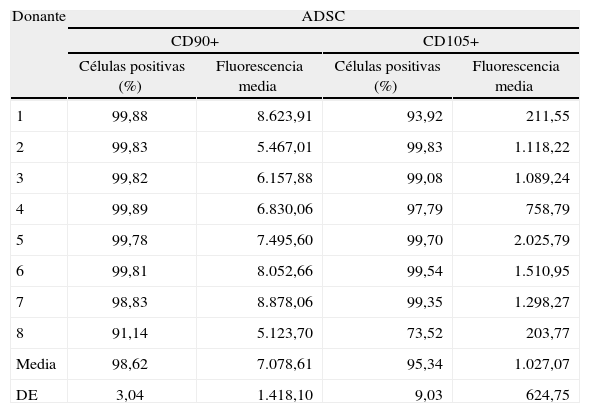
El análisis por citometría de flujo muestra que las células ADSC recién obtenidas en estado indiferenciado expresan antígenos de superficie, marcadores de células madre mesenquimales humanas (CD13+, CD90+ [Thy1] y CD105+ [endoglina]); en cambio, son negativas para el marcador hematopoyético (CD45–). En algunos estudios38 se realiza una selección de la población CD105+; sin embargo, otros autores no lo han considerado necesario, dado que el 95% de las ADSC que obtienen son CD105+, lo que muestra además la coexpresión del otro marcador mesenquimal, CD90 (Thy1)36. Además, como se muestra en la tabla I, no se observan cambios significativos en la expresión de ambos antígenos de membrana entre las células obtenidas de lipoaspirados de distintos donantes36.
Expresión de CD90 y CD105 en ADSC tras el cultivo primario
| Donante | ADSC | |||
| CD90+ | CD105+ | |||
| Células positivas (%) | Fluorescencia media | Células positivas (%) | Fluorescencia media | |
| 1 | 99,88 | 8.623,91 | 93,92 | 211,55 |
| 2 | 99,83 | 5.467,01 | 99,83 | 1.118,22 |
| 3 | 99,82 | 6.157,88 | 99,08 | 1.089,24 |
| 4 | 99,89 | 6.830,06 | 97,79 | 758,79 |
| 5 | 99,78 | 7.495,60 | 99,70 | 2.025,79 |
| 6 | 99,81 | 8.052,66 | 99,54 | 1.510,95 |
| 7 | 98,83 | 8.878,06 | 99,35 | 1.298,27 |
| 8 | 91,14 | 5.123,70 | 73,52 | 203,77 |
| Media | 98,62 | 7.078,61 | 95,34 | 1.027,07 |
| DE | 3,04 | 1.418,10 | 9,03 | 624,75 |
ADSC: adipose derived stem cells (células madre en la estroma adiposa); DE: desviación estándar.
Modificado de Talens-Visconti et al36.
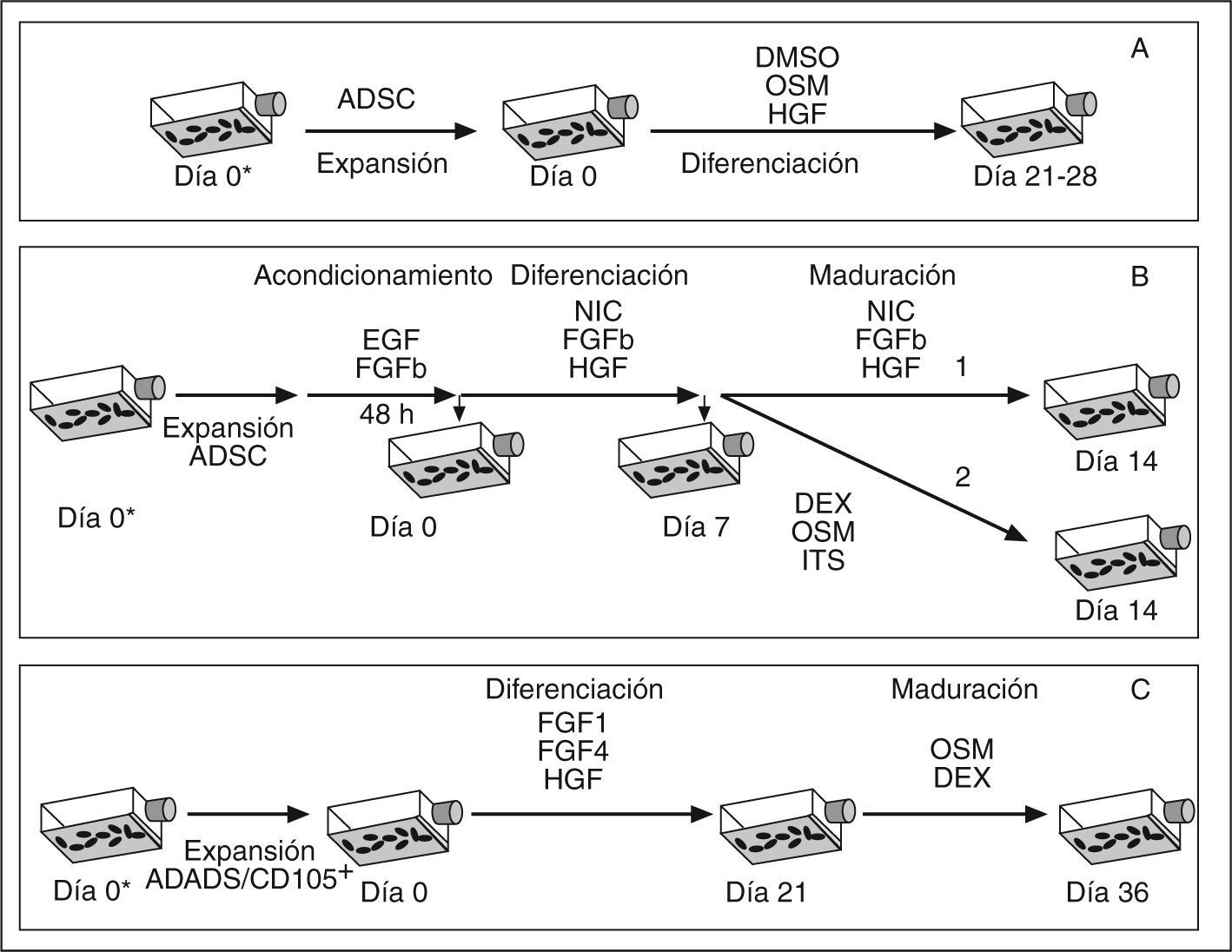
Hasta el momento, se han desarrollado diferentes protocolos in vitro para conseguir la diferenciación hepática de ADSC. Una de las propuestas se basa en el tratamiento de las células durante 3-4 semanas con un «cóctel» de factores que parecen ser críticos para inducir el fenotipo hepático, compuesto por dimetil sulfóxido (DMSO), HGF y OSM35 (fig. 4A). Otras propuestas se basan en el tratamiento secuencial con factores exógenos. Uno de los protocolos consta de un período de acondicionamiento de las células de 2 días, durante los cuales se cultivan con medio complementado con EGF y FGF. A continuación las células se mantienen durante 7 días en un medio complementado con HGF y nicotinamida (etapa de diferenciación) y, finalmente, las células se cultivan de 7 a 21 días con medio complementado con OSM, dexametasona e ITS Premix (insulina, transferrina y selenio) (etapa de maduración)36. Paralelamente, se ha investigado el tratamiento con HGF durante un período más prolongado37 (fig. 4B). Este segundo protocolo dio como resultado un mayor grado de diferenciación, en el que se obtuvieron unos niveles de expresión de genes hepáticos mayores que con el protocolo anterior. Otro estudio publicado recientemente también utiliza un protocolo secuencial para inducir la diferenciación hepática, que consiste en mantener las células adiposas en un medio de cultivo basal que contiene hidrocortisona EGF, insulina y dexametasona al que se añade HGF, FGF1 y FGF4 durante 3 semanas (etapa de diferenciación). Durante las 2 semanas siguientes las células se mantienen en el mismo medio, pero en este caso complementado con OSM y dexametasona a mayor concentración (etapa de maduración) (fig. 4C)38.

Protocolos de diferenciación hepática, A35, B37, C38. En todos ellos, el cambio de medio se efectúa cada 48 h y la retirada del suero se realiza el día 0. En el protocolo A y C el suero utilizado procede de bovino fetal, mientras que el utilizado en la etapa de expansión del protocolo B es de procedencia humana.
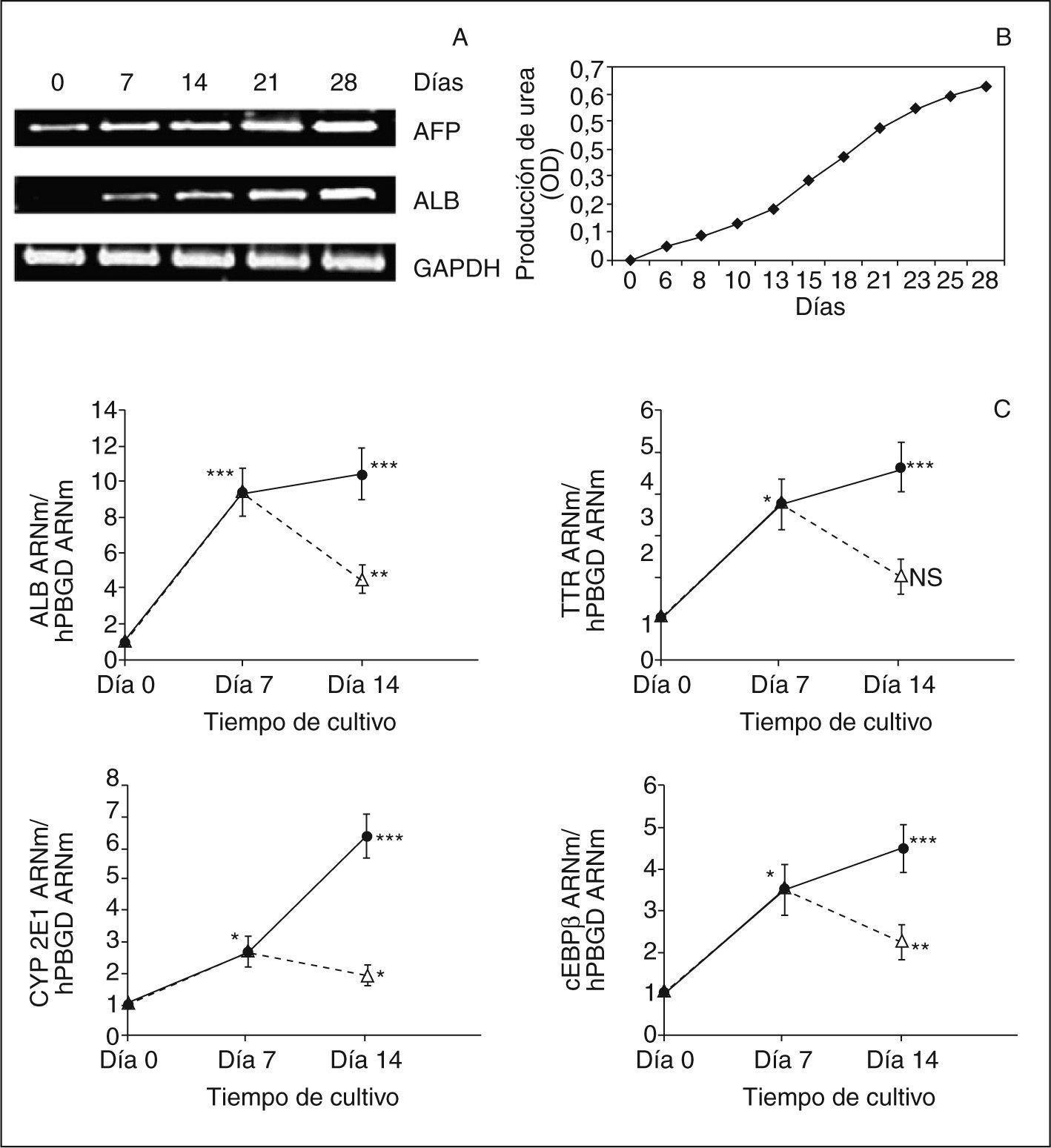
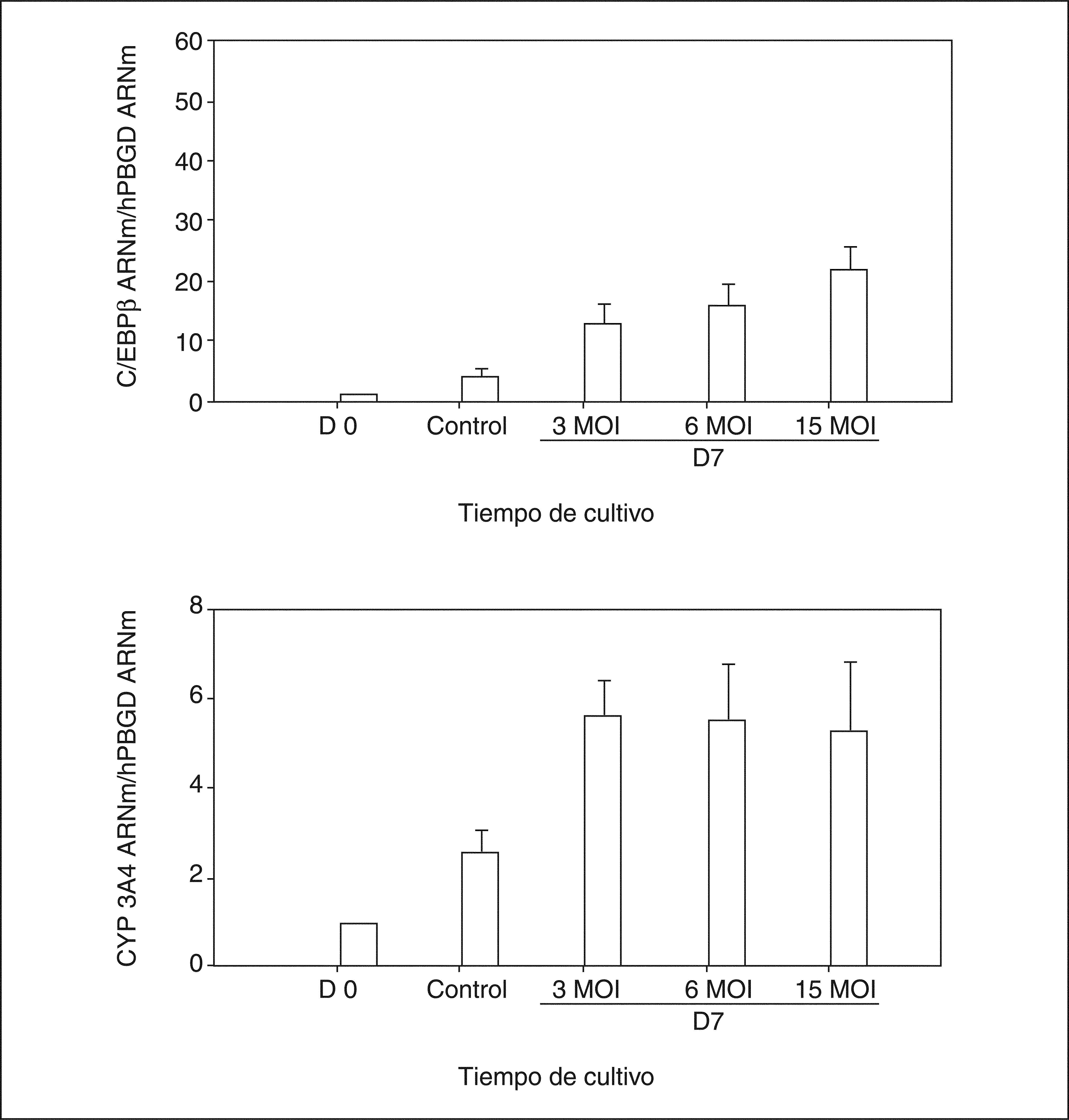
En todos los estudios realizados, la morfología celular pasa de formas fusiformes en estado indiferenciado a adoptar formas poligonales características de los hepatocitos, al final del período de diferenciación. La caracterización funcional dio como resultado un aumento creciente de la expresión de marcadores hepáticos, como albúmina, alfafetoproteína35–38 (fig. 5A y C), proteínas de membrana (CK18)38, captación de LDL38, enzimas de biotransformación, CYP 3A4 y 2E136,38 (fig. 5C), y factores de transcripción abundantes en hígado, HNF4α y C/EBPβ36,37 (fig. 5C), así como una disminución en la expresión de marcadores mesenquimales, como el Thy1 (CD90)36,37 (fig. 5C). La expresión de albúmina y alfafetoproteína se confirmó por inmunohistoquímica35,36. Tras inducir la diferenciación, se observa un incremento en la producción de urea dependiente del tiempo35,38, no detectable en las células desdiferenciadas35,38 (fig. 5B). Respecto a la capacidad de biotransformación de las ADSC diferenciadas, determinada por la expresión de isoenzimas del citocromo P450, CYP3A4 y CYP2E1, se observó un incremento significativo, que mostraba niveles de expresión muy superiores a los obtenidos en la línea celular del hepatoma humano HepG236. Si bien estas células con fenotipo fetal expresan genes característicos de hepatocitos fetales, como la albúmina79, expresan valores muy bajos o formas no activas de determinados factores de transcripción hepáticos, cuyo resultado son unos valores bajos o indetectables de expresión de marcadores hepáticos, particularmente de enzimas de biotransformación (sistema P450)36. Sin embargo, la expresión del factor de transcripción hepático C/EBPβ aumenta considerablemente durante el proceso de diferenciación de las células ADSC (fig. 5C), así como el HNF4α36. Mediante la utilización de vectores de expresión adenovirales, se ha mostrado que ambos factores de transcripción C/EBPβ y HNF4 desempeñan un papel clave en la regulación del proceso de diferenciación36,80 (fig. 6). Diversos estudios recientes demuestran que muchos de los LETF conocidos están implicados en el control de la expresión de los genes CYP en el hígado81,82. Diversos estudios también demuestran que tanto la expresión de los LETF como la de los genes CYP están estrechamente asociadas al estado diferenciado del hepatocito.
 diferenciadas en hepatocitos. A. gel de electroforesis que muestra los resultados de RT-PCR para 3 marcadores hepáticos. (Modificado de Seo et al35.) B. producción de urea por ADSC cultivadas con HGF, OSM y DMSO. La producción de urea fue cuantificada a 492 nm con un espectrofotómetro en 4 muestras diferentes. (Modificado de Seo et al35.) C. análisis de la expresión de ARNm de 4 marcadores hepáticos por RT-PCR determinados en distintos períodos del protocolo de diferenciación, protocolo 1 (círculo negro) y protocolo 2 (triángulo blanco). Resultados normalizados con hPBGD. *p < 0,05; **p < 0,01; ***p < 0,001. (Modificado de Talens-Visconti et al37.)")
Caracterización funcional de células madre en la estroma adiposa (ADSC) diferenciadas en hepatocitos. A. gel de electroforesis que muestra los resultados de RT-PCR para 3 marcadores hepáticos. (Modificado de Seo et al35.) B. producción de urea por ADSC cultivadas con HGF, OSM y DMSO. La producción de urea fue cuantificada a 492 nm con un espectrofotómetro en 4 muestras diferentes. (Modificado de Seo et al35.) C. análisis de la expresión de ARNm de 4 marcadores hepáticos por RT-PCR determinados en distintos períodos del protocolo de diferenciación, protocolo 1 (círculo negro) y protocolo 2 (triángulo blanco). Resultados normalizados con hPBGD. *p < 0,05; **p < 0,01; ***p < 0,001. (Modificado de Talens-Visconti et al37.)
 hacia fenotipo hepático. La transducción adenoviral causa un aumento dosis-dependiente en los valores de ARNm C/EBPβ determinado por RTPCR. Paralelamente, se observa una regulación positiva del CYP3A4, marcador hepático, y negativa para el Thy1, marcador de células mesenquimales. Los controles no fueron transducidos. (Modificado de Talens-Visconti et al36.)")
Evaluación del factor de transcripción C/EBPβ en la diferenciación de células madre en la estroma adiposa (ADSC) hacia fenotipo hepático. La transducción adenoviral causa un aumento dosis-dependiente en los valores de ARNm C/EBPβ determinado por RTPCR. Paralelamente, se observa una regulación positiva del CYP3A4, marcador hepático, y negativa para el Thy1, marcador de células mesenquimales. Los controles no fueron transducidos. (Modificado de Talens-Visconti et al36.)
Estos trabajos muestran que la transdiferenciación hepática de las ADSC origina células funcionales y trasplantables. En cuanto a la funcionalidad, si bien los valores de expresión de los marcadores hepáticos alcanzados distan de los encontrados en el hígado, muestran una gran proximidad a los valores observados en los hepatocitos adultos en cultivo primario y son muy superiores a los descritos en hepatoblastomas humanos. Un aspecto muy importante para asegurar el éxito del trasplante es su capacidad para implantarse o quedar retenidas en el hígado receptor. En relación con ello, se ha descrito que las células diferenciadas in vitro implantadas en ratones desnudos NOD/SCID con fallo hepático agudo inducido por CCl4, se incorporan directamente en el hígado, confirmado por la inmunotinción de albúmina humana en el hígado del ratón. Todo ello pone de manifiesto el extraordinario potencial hepatogénico y terapéutico de las ADSC35,38 para el trasplante celular.
PERSPECTIVAS DE FUTUROLos resultados obtenidos recientemente con las células progenitoras adultas derivadas de tejido adiposo despiertan un gran interés. Poseen ventajas objetivas para constituir una buena fuente alternativa de células adultas autólogas. La terapia celular autóloga evitaría complicaciones y costosos tratamientos de supresión del sistema inmunológico para prevenir rechazos o las contaminaciones provenientes de un donador. Por otra parte, la posibilidad de crioconservación de estas células sin pérdida de funcionalidad incrementaría su disponibilidad.
La obtención de células de fenotipo hepático derivadas de procesos de diferenciación in vitro de células progenitoras adultas de tejido adiposo confirma aún más, si cabe, el elevado grado de plasticidad de este tipo celular y su posibilidad de dar lugar a estirpes celulares, no sólo originadas a partir del mesodermo, sino también del endodermo y el ectodermo.
El gran conocimiento que se ha alcanzado sobre las funciones celulares y su regulación hace que el método de trasplante celular se presente con una gran posibilidad de éxito con los conocimientos médicos actuales (a diferencia del xenotrasplante o la terapia génica). Esto lo convierte en una opción con posibles resultados a corto plazo. Sin embargo, la disponibilidad de este recurso celular requiere una optimización del protocolo de diferenciación y un mayor conocimiento de los mecanismos celulares y moleculares que controlan la transdiferenciación a hepatocitos.
Los modelos celulares hepáticos existentes en la actualidad no poseen las características necesarias para cubrir las necesidades de estudio en farmacotoxicología y biomedicina. El establecimiento de un modelo celular hepático diferenciado tendría gran utilidad en estudios de metabolismo y toxicidad de medicamentos, en estudios de fisiopatología hepática, en aplicaciones clínicas, como el biorreactor hepático, o como fuente de recursos para la terapia celular. Los modelos celulares actuales presentan en común una desdiferenciación y una pérdida de funciones características del hígado, como la biotransformación de xenobióticos. Por tanto, las perspectivas de futuro derivadas de la obtención de células de fenotipo hepático a partir de ADSC son muchas, ya que no sólo se trata de un recurso abundante de células para terapia celular, sino que a su vez supone el establecimiento de un nuevo modelo celular diferenciado útil para estudios de toxicidad y metabolismo de fármacos.
Sin embargo, muchos son los factores involucrados en la organogénesis hepática, cuyo papel no se ha explorado todavía. Por ello, en el futuro próximo uno de los objetivos debe ser perfeccionar y optimizar el protocolo de diferenciación actual, investigando el efecto de otros factores que pudieran estar involucrados en la diferenciación (BMP-2, BMP-4, activina A)66,67. Por otra parte, los componentes de la matriz extracelular, como el colágeno o la fibronectina, también desempeñan un papel clave en la organogénesis del hígado74,75. Es de interés investigar su contribución en la transdiferenciación in vitro.
AGRADECIMIENTOSLos autores agradecen la ayuda prestada por la fundación ALIVE y la Unión Europea (Proyectos LSHB-CT-2004-504761, LSHB-CT- 2004-512051 y LSSB-CT-2005-037499).

