







Preadolescent children undergo developmental changes in inhibitory control. Maintenance of high levels of moderate-to-vigorous-intensity physical activity (MVPA) has been suggested to promote its maturation. We compared inhibitory control between children with low and high MVPA as well as their inhibitory processing stream and changes in cortical hemodynamics.
Method109 participants aged 10 to 13 years wore accelerometers over 7 days. Those with MVPA levels of 30 min/d or less and 60 min/d or more further performed a computerized Stroop Color-Word task. Electroencephalography and functional near-infrared spectroscopy were used to record changes in inhibitory processing and cortical hemodynamics, respectively.
ResultsAn interaction of MVPA group and sex indicated better interference in highly-active boys, but the opposite pattern in girls. Independent from sex, the high compared to low MVPA group showed greater P300 and PSW amplitudes, whereas no group differences were found for N200, N450, and changes in cortical hemodynamics.
ConclusionChildren with high MVPA differ from their less-active peers by a distinct inhibitory processing profile, which is characterized by altered allocation of attentional resources and conflict resolution. However, these alterations do not necessarily translate into better performance, especially since MVPA is linked with higher inhibitory control in boys only.
Cognitive development across childhood and adolescence is coupled with changes in physical fitness and activity (Tandon et al., 2016). This is also acknowledged by the World Health Organization (WHO), which recommends a daily amount of 60 min of moderate-to-vigorous-intensity physical activity (MVPA) to promote cognitive abilities along with other domains of health in children and adolescents (Chaput et al., 2020). Preadolescence is a critical period characterized by a pronounced decline in physical activity levels (Farooq et al., 2020), which in turn may affect the trajectory of these cognitive abilities. The maturation of inhibitory control is an important developmental target as it has been conceptualized as the common component of executive function, which summarizes cognitive processes engaged during the top-down control of behavior (Miyake & Friedman, 2012). Higher inhibitory control is predictive of better classroom behavior (Ludyga, Gerber, Brand, Möhring & Pühse, 2022a), academic performance (Allan, Hume, Allan, Farrington & Lonigan, 2014), and socio-emotional competence (Rhoades, Greenberg & Domitrovich, 2009). In turn, children with poor inhibitory control face the risk of developing problematic externalizing behavior and symptoms of Attention Deficit Hyperactivity Disorder (Bohlin, Eninger, Brocki & Thorell, 2012). Meta-analytical findings indicate that long-term physical exercise interventions elicit small benefits for inhibitory control in children and adolescents (Xue, Yang & Huang, 2019). While the term physical exercise refers to physical activities that are planned, structured and repetitive with the aim to maintain or increase the performance in one or more fitness dimensions, the term physical activity includes all unplanned, loosely-structured bodily movements that increase the energy expenditure above ∼1.0/1.5 MET (metabolic equivalent of the task; 1 MET = 1 kcal.kg−1 h−1) (Caspersen, Powell & Christenson, 1985). Due to this conceptual difference, their relation with inhibitory control can vary in terms of consistency. In this respect, results indicating higher inhibitory control or executive function among preadolescents with higher MVPA (Zeng et al., 2021; Zhu et al., 2023) are contrasted by studies showing either no association (Mücke, Andrä, Gerber, Pühse & Ludyga, 2018) or an association in subgroups only, such as overweight individuals (Patraca-Camacho et al., 2022). This discrepancy observable in behavioral outcomes, may be due to the issue that even if MVPA leads to alterations in the inhibitory processing stream, these might not necessarily translate into better performance. Consequently, a better understanding of this link requires a further exploration of its potential underlying neurocognitive processes.
The Stroop Color-Word task is a common way to assess the interference aspect of inhibitory control and requires participants to react to the color of ink of color words, while ignoring their meaning. When combined with electroencephalography (EEG), different cognitive control processes can be isolated via event-related potentials (ERPs). The presentation of a color word elicits the N200 at 200 to 300 ms, which is tied to monitoring the conflict between color of ink and word meaning (Pires, Leitão, Guerrini & Simões, 2014). The subsequent P300 peaks at around 300 ms and its amplitude is proportional to the allocation of attentional resources towards the task (Polich, 2007). In the following 400 to 500 ms latency range, processing of the stimulus conflict and the suppression of interference is indexed by the N450 (Di Russo & Bianco, 2023). The final ERP component of the inhibitory processing stream during the Stroop task is the positive slow wave (PSW) that peaks around 600 ms and shows a centro-parietal distribution comparable to the P300 (Heidlmayr, Kihlstedt & Isel, 2020). While both components reflect the engagement of executive processes, conflict resolution is only indexed by the PSW (Heidlmayr, Hemforth, Moutier & Isel, 2015). A review of observational and experimental studies found that physical activity is generally associated with a greater amplitude of the P300 in various executive function tasks across all ages (Kao et al., 2020). In contrast, evidence on the link between habitual movement behaviors and other ERP components is lacking in children. The few available findings from adults suggest that physically active individuals differ from less active peers in terms of N200 and N450 amplitudes (Aly & Kojima, 2020; Gajewski & Falkenstein, 2015), indicating a facilitation of conflict monitoring and processing, respectively. Consequently, MVPA has the potential to influence children's inhibitory control at various stages within the processing stream.
In addition to the subtle cognitive processes that can be examined by ERPs, interference during the Stroop task further depends on local or global changes in cortical hemodynamics, which can be measured using functional near-infrared spectroscopy (fNIRS) (Ludyga, Mücke, Colledge, Pühse & Gerber, 2019). Children and adolescents performing tasks tapping into inhibitory control show an increase in oxygenated hemoglobin at left, middle and right frontal gyrus, with a greater response indicating a more mature developmental pattern (Yeung, Lee & Chan, 2020). MVPA seems to promote this response profile as male adults fulfilling the WHO recommendations on physical activity were characterized by a more pronounced increase of oxygenated hemoglobin than their less active counterparts during executive function tasks (Goenarjo et al., 2020). Similarly, higher levels of physical activity in female adults have been associated with increased hemodynamic response at frontal brain regions, which in turn explained variance in behavioral performance on an inhibitory control task (Cameron, Lucas & Machado, 2015). This relation seems to apply to various regions of the frontal lobe, given that higher physical activity levels have also been related to a greater increase of oxygenated hemoglobin in the inferior frontal cortex and dorsolateral prefrontal cortex (Makizako et al., 2013; Matsuda et al., 2017). While evidence from adults is consistent in terms of the direction of effects, less is known on whether the association of physical activity and changes of cortical hemodynamics in response to an inhibitory control task generalizes to children. The only study available for this age group showed no differences between children with low and high MVPA (Mücke et al., 2018). However, it should be noted that oxygenated hemoglobin was measured in response to tasks assessing separable components of executive function (i.e. cognitive flexibility and working memory updating). As changes in cortical hemodynamics measured with fNIRS and the cognitive processes covered by ERPs influence interference in the Stroop task, a combination of both allows the investigation of their relative contribution to differences in performance observed among children with variable MVPA levels.
We aimed to compare interference between preadolescent children fulfilling the WHO recommendations on MVPA and less-active peers. Additionally, we investigated potential group differences in the inhibitory processing stream and the task-related upregulation of oxygenated hemoglobin in the frontal gyrus. Based on the previous literature, we expected a lower interference in highly active children (Zeng et al., 2021; Zhu et al., 2023), complemented by a more effective cognitive control (at the level of resource allocation, conflict monitoring, processing or resolution) and a greater increase of oxygenated hemoglobin (Aly & Kojima, 2020; Cameron et al., 2015; Gajewski & Falkenstein, 2015; Kao et al., 2020; Matsuda et al., 2017).
MethodParticipantsThe present study reports data from the CASPA trial, which investigates the associations of physical activity, stress and inhibitory control. Flyers containing information on the study were distributed at schools, public spaces, sports clubs, and medical practices in Basel (Switzerland) to recruit children aged 10 to 13 years. Further inclusion criteria were MVPA of either at least 60 min/d or 30 min/d or less, the ability to fully engage in physical activity within the past 3 months, right-handedness, and corrected-to or normal vision. The MVPA cut-offs were chosen to differentiate between preadolescents that are clearly below and above average MVPA (43 to 50 min/d) reported for Swiss preadolescents (Verloigne et al., 2012). Moreover, the upper cut-off corresponded to the daily amount of physical activity recommended by the WHO (Chaput et al., 2020). Exclusion criteria were conditions that increase the health risk during physical activity (based on parent-report, e.g. acute infection, syncope, uncontrolled arrhythmia), attendance of special education services, presence of a psychological disorder, and sleep complaints. Written informed consent was obtained from legal guardians of the participants. The study protocol was approved by the local ethics committee (Ethikkommission Nordwest- und Zentralschweiz) and procedures were in accordance with the Declaration of Helsinki.
ProceduresPrior to the laboratory visit, participants filled in the Insomnia Severity Index, the Family Affluence Scale and the Strengths and Difficulties Questionnaire and received accelerometers to record physical activity. This objective assessment was complemented by a 7-day physical activity recall protocol. If participants were injured or reported illness, the assessment was repeated after a few weeks. Participants fulfilling the inclusion criteria were assigned to a low (≤ 30 min/d) or high (≥ 60 min/d) MVPA group based on their individual physical activity levels and completed two laboratory visits. However, only the outcomes of the first visit were used for the present study. Participants first completed a computer-based Stroop task for familiarization, which was repeated after 20 min to assess performance. The second measurement was combined with simultaneous EEG-fNIRS recordings. Anthropometric characteristics (height, weight, and body mass index) were collected and participants filled in the Pubertal Development Scale. Assessments were performed in a dimly-lit room and at an environmental temperature between 20 and 22 °C, with surrounding noise kept to a minimum.
Physical activityParticipants wore triaxial accelerometers (ActiGraph wGT3X-BT, ActiGraph, Pensacola, USA) on the hip over seven consecutive days, which included a regular school week. Data was recorded with a sampling rate of 50 Hz and submitted to ActiLife 6.13.4 (ActiGraph, Pensacola, USA) for offline processing. The datasets of participants were considered valid, if the recordings included at least six days with 10 h of wear-time per day (during daytime). Wear-time validation was performed with the algorithm by Troiano et al. (2008). From the counts per minute, daily time spent in light, moderate, and vigorous intensity was calculated based on an algorithm that is suitable for the age range of the current sample (Mattocks et al., 2007). Following the accelerometer recordings, participants recalled their physical activity with a 7-day protocol. Moderate and vigorous intensity had to be differentiated and participants also recalled the timing and duration of their activities. The recall protocol was used to replace non-wear time that was due to physical activities that could not be recorded with accelerometers, because they posed an increased risk for injury of the participants (e.g. martial arts) and/ or for damaging the device (e.g. swimming).
Cognitive taskThe computerized Stroop task was administered with E-Prime 3.0 (Psychology Software Tools). Visual stimuli were the German color words “blau” (blue), “grün” (green), and “gelb” (“yellow”), which were selected based on an equal word length. These color words were written in the same color of ink on compatible trials and a different color of ink on incompatible trials. The two trial types were presented in different blocks and the order of the color words followed a randomized order. Participants were instructed to press a button corresponding to the meaning of the color word, but to ignore its color of ink. Each trial commenced with a fixation period (i.e. black screen with fixation cross) that varied randomly between 900 and 1100 ms. The subsequent stimulus was presented against black background for 200 ms and responses were collected within 1100 ms following its onset. The next trial was initiated after the response window, meaning that trial and block duration was equal among all participants. Prior to cognitive testing, four stimuli examples with the correct response were shown. To ensure that participants understood the task, they first completed one practice block with 12 trials for each trial type. Subsequently, eight compatible and eight incompatible blocks with the same amount of trials were administered. The order of the blocks was alternated. Accuracy and average reaction time on response-correct trials were extracted. Additionally, interference was calculated by subtracting the reaction time on compatible from incompatible trials.
Combined EEG-FNIRS recordings32 active electrodes and 16 optodes (8 illumination sources, 8 light detectors) were placed in a single head cap for combined EEG-fNIRS recordings. The arrangement of the electrodes followed the 10:20 system. AFz served as ground and the vertex channel was used as online reference. Prior to the recordings, a highly-conductive gel was applied between the scalp and electrodes to reduce impedance to 10 KΩ or lower. Data was amplified using the actiCHamp (Brain Products GmbH, Gilching, Germany), digitized at a sampling rate of 512 Hz, and recorded with the software BrainVision Recorder 1.21 (BrainProducts GmbH, Gilching, Germany).
A continuous-wave, dual-wavelength (760 and 850 nm) fNIRS system (NIRSport, NIRx Medical Technologies, Berlin, Germany) acquired data at a sampling rate of 7.8125 Hz, which was recorded using the software Aurora 2021.9 (NIRx Medical Technologies, Berlin, Germany). The fNIRS optodes (LED light sources and avalanche photodiode detectors) were placed over regions of the frontal lobe (Supplement, Figure S1) and assigned to regions of interest based on corresponding EEG positions (Scrivener & Reader, 2022). Inter-optode distances of 3.0 cm were maintained with the use of spacers to achieve the best trade-off between light penetration depth and signal-to-noise ratio (Herold, Wiegel, Scholkmann & Müller, 2018). Additionally, short-distance detectors were mounted in 0.8 cm distance to the light sources to control for changes in extracerebral hemodynamics (Tachtsidis & Scholkmann, 2016). Participants wore a loosely attached, optically opaque overcap to prevent ambient light from interfering with the fNIRS data recording. Before each measurement, signal level optimization was performed automatically by the software and optodes were adjusted until the coefficient of variation and the signal level reached an acceptable range of <7.5 % and >0.5 mV, respectively.
Data processingBESA Research 7.1 (Brain Electric Source Analysis, Gräfelfing, Germany) was employed for processing of EEG data. Following the detection of blinks and eye movements based on virtual VEOG and HEOG channels, continuous data was submitted to automatic adaptive artifact correction. This approach first uses principle component analysis to contrast dominant brain activity against artefacts and then subtracts reconstructed artefact components from the data based on spatial filtering (Ille, Berg & Scherg, 2002). High-pass (forward phase shift of 0.3 Hz; slope 6 dB/octave) as well as low-pass filters (zero-phase shift of 30 Hz; slope 24 dB/octave) were applied and baseline-correction was performed using the 200 ms period prior to stimulus onset. Subsequently, artefacts that survived the initial correction procedure were rejected based on individual amplitude and gradient thresholds. The reference was changed to the average of mastoids. Segments with correct responses that contained no artefacts were created for the −200 to 1000 ms latency range and averaged separately for the trial types. Difference waves were calculated by subtracting the compatible from the incompatible trials. The number of good segments did not differ between the low and high MVPA group. The criteria for the extraction of ERP components (as mean amplitude in a specified time range) was guided by the previous literature (Heidlmayr et al., 2020; Kóbor et al., 2015; Polich, 2007) and adjusted, if necessary, based on the visual inspection of the grand average waveform of all participants. Within the latency range from 270 to 350 ms, the N200 was extracted from the fronto-central region (FC1, FC2, F3, F4, Fz) and the P300 from the parietal region (P3, P4, Pz). In the subsequent 400–500 ms time window, the N450 as calculated as mean amplitude at frontal region (F3, F4, Fz). Lastly, the centro-parietal (CP1, CP2, Pz) mean amplitude 450–650 ms following stimulus onset was used to extract the PSW.
The processing of fNIRS data was performed with Homer3 v1.52.0 (Huppert, Diamond, Franceschini & Boas, 2009). In the processing stream, missing values were first replaced by spline interpolation of the existing values. With the function hmrR_PruneChannels, channels with a dRange lower than 500 μV or higher than 1 V, and a quotient of mean(d)/standard deviation(d) below 6.67 were removed from further analysis. After conversion from light intensity to optical density, motion artefacts were first detected using hmrR_MotionArtifactByChannel (time range 0.5 s, masked data around motion artefact 0.5 s, standard deviation threshold 25, amplification threshold 5) and then corrected using spline interpolation (p = 0.99) and wavelet correction (interquartile range = 1). Surviving artefacts were detected by repeating hmrR_MotionArtifactByChannel, and then rejected using hmrR_StimRejection. The subsequent bandpass filter used a thresholds of 0.01 HZ (high pass filter) and 0.09 Hz (low pass filter) to exclude instrumental noise and systemic noise (e.g. heart rate, breathing, Mayer waves) from the signal (Herold et al., 2018). The filtered data was converted from optical density to concentration changes of the chromophores (i.e., oxygenated and deoxygenated hemoglobin) using a partial path length factor of 1. A general linear model using ordinary least squares as a solving method, a consecutive sequence of Gaussian functions as a basis function, and a polynomial drift correction of third order was applied to estimate the hemodynamic response function. Therein, block averages were defined as the time range −3 s to 40 s, and short separation regression was performed with the nearest available short-distance channel (Gagnon et al., 2012). Hemodynamic response functions were exported and mean values over the task period (0 – 26 s) were calculated at (left and right) middle and superior frontal gyrus, with four channels averaged per region of interest.
Statistical analysisStatistical analyses were performed with SPSS 28.0 for Windows (IBM, Armonk, NY, USA). In advance, one-way ANOVAs were applied to investigate group differences in MVPA, anthropometrics, perceived stress, socio-economic status, sleep, pubertal status, and psychopathology. Pearson correlations were calculated between background variables showing group differences and outcomes (interference, ERP components, changes in cortical hemodynamics). Variables showing a statistically significant association with one or more outcomes served as covariates in subsequent main comparisons. Group differences in interference were investigated using one-way ANOVA with MVPA (low, high) and sex (boys, girls) as between-subject factors, while accuracy was accounted for. Sex was included as factor, given that girls and boys differ in the development of inhibitory control (Usai, 2023) as well as changes in MVPA across childhood and adolescence (Farooq et al., 2020). Group differences in cerebral oxygenation changes were examined by employing ANOVA with the same between-subject factors and repeated measures on trial type (compatible, incompatible) as well as region of interest (left middle frontal gyrus, right middle frontal gyrus, superior frontal gyrus). Main effects of group and trial type as well as interactions with group were reported. With regard to cognitive control, differences between the low and high MVPA group were analyzed by applying a MANOVA on N200, P300, N450, and PSW amplitudes with repeated measures on trial type. Multivariate main effects of trial type and group as well as interactions with group were followed-up with univariate analyses. For all analyses, three-way interactions were decomposed using Bonferroni-corrected post hoc tests. The level of statistical significance was set to p ≤ 0.05.
ResultsOf 111 recruited participants, 109 (61 boys, 48 girls) had complete data on all outcomes and were included in the statistical analyses. Scores on the Pubertal Development Scale indicated that boys were in early pubertal stage (4.6 ± 1.7) and girls in midpubertal stage (6.5 ± 2.8). The groups differed in their MVPA levels (25.1 ± 8.9 min/d vs. 70.8 ± 12.2 min/d), F(1, 107) = 336.77; p < 0.001; η2 = 0.759. The high MVPA group reported less perceived stress, had a higher socioeconomic status, and a lower pubertal status than the low MVPA group (Table 1). Additionally, there was a trend towards lower psychopathology in the high MVPA group. However, none of these variables were associated with interference, r ≤ 0.10, p ≥ 0.284, or P300 and PSW amplitudes, r ≤ (-)0.14, p ≥ 0.149. A trend towards a statistical significant correlation was found between N200 (on incompatible trials) and perceived stress as well as N450 (on incompatible trials) and pubertal status. For fNIRS data, greater changes in cortical hemodynamics were related to lower pubertal status and perceived stress, r ≤ −0.20, p ≥ 0.038. Consequently, both variables were entered as covariates for analyses on cortical hemodynamics.
Comparison of anthropometrics, socio-economic status, sleep and psychopathology between groups with low and high moderate-to-vigorous-intensity physical activity.
Notes: FAS = Family Affluence Scale; ISI = Insomnia Severity Index; Moderate-to-vigorous-intensity physical activity; PSS = Perceived Stress Scale; SDQ = Strengths and Difficulties Questionnaire.
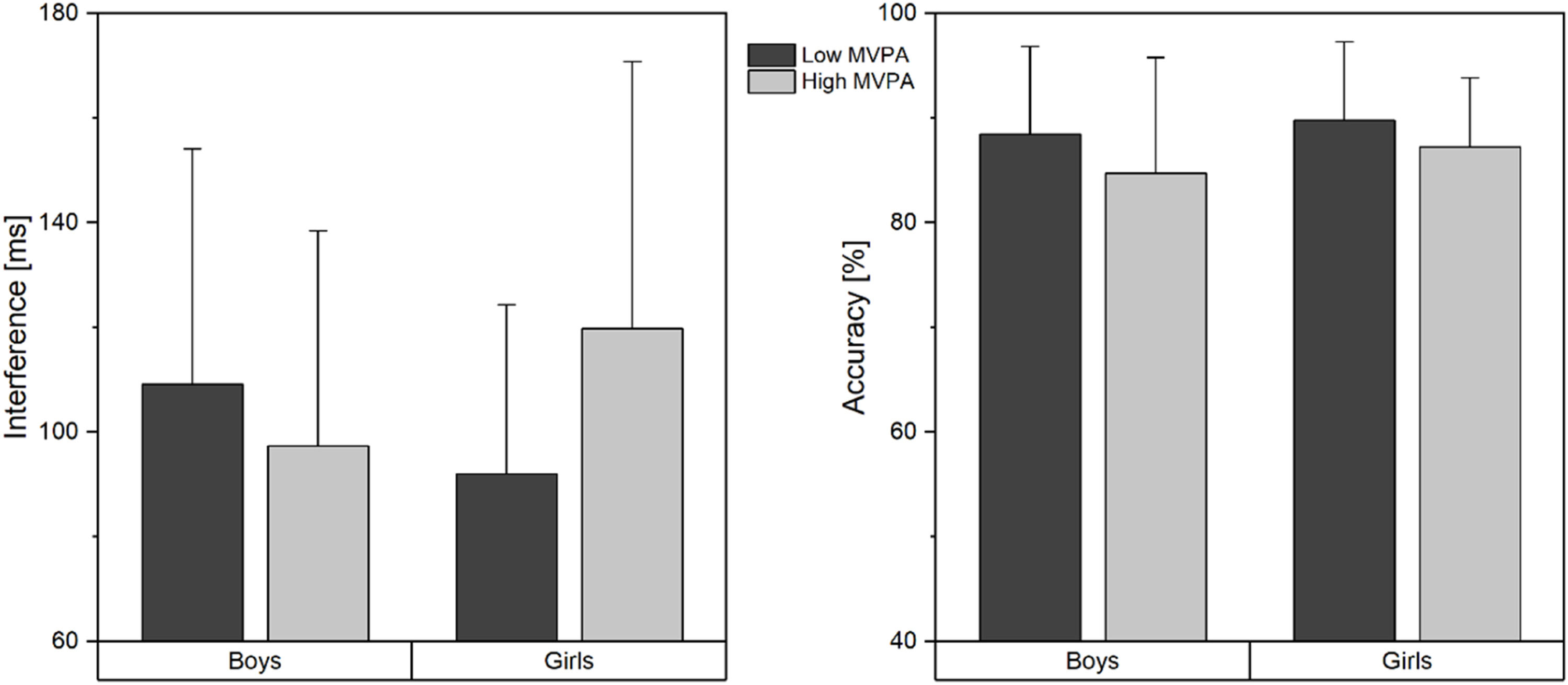
With regard to cognitive performance, the univariate analysis yielded no main effect of group on interference, F(1104) = 1.46, p = 0.230, η2 = 0.014, when accuracy was controlled for. However, there was an interaction of group and sex, F(1104) = 5.35, p = 0.023, η2 = 0.049, indicating that boys with high MVPA showed lower interference than boys with low MVPA, while this pattern was reversed for girls (Fig. 1).
Event-related potentials on the Stroop task. Notes: MVPA = Moderate-to-vigorous-intensity physical activity.")
The event-related potential waveforms of both groups are displayed in Fig. 2. The MANOVA yielded a main effect of trial type, Wilks's λ (4103) = 0.518, p < 0.001, η2 = 0.482, and group, Wilks's λ (4103) = 0.904, p = 0.033, η2 = 0.096, on cognitive control processes. Follow-up univariate analyses of the main effect of trial type showed that only for PSW, compatible trials elicited a lower amplitude than incompatible trials, F(1106) = 68.94, p < 0.001, η2 = 0.394. The univariate analyses of the group effect further revealed that participants with high MVPA showed a higher P300 amplitude, F(1106) = 7.81, p = 0.006, η2 = 0.069, and PSW amplitude, F(1106) = 4.32, p = 0.040, η2 = 0.039, compared to participants with low MVPA (Fig. 3). In contrast, no main effect of group was found for N200 and N450 amplitudes.
 as well as the N450 (400–500 ms) and PSW (450 to 650 ms). MVPA = Moderate-to-vigorous-intensity physical activity.")
Event-related potential waveforms displayed for groups with low and high moderate-to-vigorous-intensity physical activity. Notes: Solid lines represent compatible trials, whereas dashed lines represent incompatible trials. Topographic plots are shown for N200 and P300 (270 to 350 ms) as well as the N450 (400–500 ms) and PSW (450 to 650 ms). MVPA = Moderate-to-vigorous-intensity physical activity.
 elicited by compatible and incompatible trials displayed for participants with low and high moderate-to-vigorous-intensity physical activity. Notes: MVPA = Moderate-to-vigorous-intensity physical activity.")
Waveforms of oxygenated hemoglobin in response to the task are shown in the Supplement (Fig. S2). The results of the repeated measures ANOVA (adjusted for perceived stress and pubertal status) support a main effect of trial type on cerebral oxygenation, F(1,94) = 4.38, p = 0.039, η2 = 0.045, indicating a higher concentration on compatible compared to incompatible trials. In contrast, there was no main effect of group on the concentration of oxygenated hemoglobin (Fig. 4), F(1,96) = 0.01, p = 0.940, η2 < 0.001.
 at middle frontal gyrus and superior frontal gyrus displayed for participants with low and high moderate-to-vigorous physical activity. Notes: MFG = Middle frontal gyrus; MVPA = Moderate-to-vigorous intensity physical activity; SFG = superior frontal gyrus.")
Changes of the concentration of oxygenated hemoglobin (means±standard deviation) at middle frontal gyrus and superior frontal gyrus displayed for participants with low and high moderate-to-vigorous physical activity. Notes: MFG = Middle frontal gyrus; MVPA = Moderate-to-vigorous intensity physical activity; SFG = superior frontal gyrus.
Partial correlations that controlled for accuracy showed that higher negativity of the N200 amplitude (elicited from incompatible trials) was related to a higher interference, r = −0.20, p = 0.039. Other ERP components and changes of oxygenated hemoglobin recorded via fNIRS were not associated with interference (see Supplement).
DiscussionIn contrast to our initial hypothesis, there was no difference between groups in interference, unless its interaction with sex was considered. Boys with high MVPA had a lower interference on the Stroop task than boys with low MVPA, whereas this pattern of results was reversed in girls. These findings were decoupled from the neurocognitive profile of the groups. Independent from sex, participants with high compared to low MVPA showed increased P300 and PSW amplitudes, but these ERP components did not explain variance in interference.
A meta-analysis of experimental studies indicated that across all ages, male participants can expect greater benefits for executive function and other cognitive domains than their female counterparts following a structured physical exercise intervention (Ludyga, Gerber, Pühse, Looser & Kamijo, 2020). Although exercise is an integral part of physical activity, the role of sex is even more pronounced in our study, given that the direction of effects differed between boys and girls. The lack of an interaction of group and sex on the inhibitory processing stream and changes of oxygenated hemoglobin indicates that this pattern of results cannot be explained by the neurocognitive mechanisms we focused on. An alternative explanation might be derived from the experiences made during physical activity. Longitudinal findings underline that during the transition to adolescence, enjoyment of (school-based) physical activity decreased over time in girls, but remained unchanged in boys (Cairney et al., 2012). This might be due to a set of barriers of physical activity, which are present for both sexes, but experienced more frequently by girls, e.g. teasing from both the same and the different sex (Slater & Tiggemann, 2011) as well as perceived lack of energy (Rosselli et al., 2020). The effort needed to overcome these challenges might have influenced the association of MVPA and inhibitory control. This is supported by a review suggesting that mood, which might respond differently in girls and boys, is one of the mechanisms by which physical exercise and physical activity elicits improvements in different cognitive domains (Stillman, Esteban-Cornejo, Brown, Bender & Erickson, 2020). While mood offers an explanation of high interference despite high MVPA in girls, it remains unclear why those with low MVPA show better performance on the Stroop task. In this respect, a review of experimental evidence has suggested that similar to physical exercise, performing arts and related activities benefit executive function by a common skill acquisition process (Tomporowski & Pesce, 2019). Thus, girls might compensate low MVPA with greater involvement in other activities that equally promise cognitive benefits, which is partly supported, for example, by a higher rates of engagement of female individuals in performing arts (Lagaert, van Houtte & Roose, 2017).
The results of the neurocognitive measures were not consistent with the influence of group and sex observed for behavioral performance. The lack of group differences in N200 and N450 amplitudes indicated that MVPA levels had no impact on the monitoring and processing of the color-word conflict. Additionally, the trial type did not modulate these components in our sample, which can be interpreted as a difficulty to effectively detect the conflict. In contrast to N200 and N450, the P300 that peaks between these two components was greater in the high compared to the low MVPA group. An increased P300 amplitude, which is thought to reflect more effective allocation of attentional resources by inhibiting task-irrelevant information (Polich, 2007), is in line with a quantitative synthesis showing a similar change following acute and long-term physical activity (Kao et al., 2020). A mechanism underlying the sensitivity of the P300 might be the phasic activity of the neuromodulatory locus coeruleus-norepinephrine system (LC-NE) that is reflected by this ERP component. More specifically, it has been suggested that the P300 marks the response of the LC-NE to internal decision-making processes and the subsequent potentiation of information processing by the noradrenergic influence (Nieuwenhuis, Aston-Jones & Cohen, 2005). This has been supported by a correlation of P300 amplitude and tonic pupil dilation, which serves as a marker of the LC-NE (Murphy, Robertson, Balsters & O'connell, 2011). Engaging in a physical activity or physical exercise bout is known to engage arousal mechanisms by activating the LCNE (Dietrich & Audiffren, 2011) and this is reflected by a tonic dilation of pupils, which in turn, has been linked with improved inhibitory control (Shigeta et al., 2021). Consequently, an increased P300 amplitude in the high MVPA group can be interpreted as a facilitation of the LC-NE responsiveness due to repeated stimulation by a range of physical activities.
Similar to P300, the amplitude of the PSW was greater in the high MVPA group. Compared to other components elicited by the Stroop task, its functional significance is less clear. A recent review has suggested that the PSW mainly reflects conflict resolution, although the subsequent semantic reactivation of word meaning has also been attributed to this component (Heidlmayr et al., 2020). Among the processes examined in our study, only the PSW increased for trials that presented conflicting information, which might be interpreted as a higher demand on resources recruited for the resolution of the color-word interference. Based on source analysis, a neural generator of the PSW is the anterior cingulate cortex (Hanslmayr et al., 2008), which is recruited at various stages of processing a perceptual or response conflict (Kim, Kroger & Kim, 2011). In this respect, previous findings in young adults showed that higher MVPA levels were related to better inhibitory control via increased thickness of the anterior cingulate cortex (Bento-Torres et al., 2019). Consequently, an increased PSW might result from the influence of MVPA on its function or structure. It should be noted, however, that increased PSW did not contribute to performance on the Stroop task.
With regard to the fNIRS findings, changes in cortical hemodynamics did not differ between groups and could not explain the interaction of group and sex for interference. Our results are inconsistent with previous studies in other age groups, showing that physical activity leads to a greater oxygenation hemoglobin concentration within different regions of the frontal lobe during executive function tasks (Cameron et al., 2015; Makizako et al., 2013; Matsuda et al., 2017). This discrepancy might be due to age differences as the oxygenation response to interference during the Stroop task is known to increase from childhood to adolescence (Yeung et al., 2020). We observed no increase of oxygenated hemoglobin from compatible to incompatible trials, thus reflecting an immature response pattern. This might be compensated by other mechanisms, meaning that behavioral performance on the Stroop task does not rely on the oxygenation of the frontal lobe. Conversely, MVPA might not influence inhibitory control via oxygenation changes at this developmental stage.
While our findings provide insights into the role of MVPA for the individual inhibitory processing profile, alterations cannot be considered an advantage or disadvantage as this profile was independent from behavioral performance on the Stroop task. As changes in subtle cognitive processes are expected to precede observable performance differences, it remains unclear if altered resource allocation and conflict resolution translates into such differences in the long-term. The cross-sectional design of the study further limits conclusions on the direction of effects. Increased P300 and PSW amplitudes can be seen as an adaptation in response to high MVPA, but they could also characterize an inhibitory control trait that is more likely to engage in MVPA. On a methodological level, we opted for an objective assessment of MVPA, complemented by recall protocols. This approach limits conclusions to the influence of the quantity rather than quality of physical activity. However, the type of activity can also influence the inhibitory processing stream in adolescents (Ludyga, Mücke, Andrä, Gerber & Pühse, 2022b), meaning that a combination of both approaches might promise a better understanding of the influence of MVPA on interference. In our analyses, we considered important confounders, such as sex, socio-economic status, and pubertal status. However, the low interference in girls with low MVPA provides an indication for the engagement in compensatory activities. Consequently, participation in other leisure activities, with potential to enhance inhibitory control, needs to be controlled. Similarly, physical activity experiences and enjoyment should be considered along with the quantity of these activities. This psychosocial domain may help to better understand the moderating role of sex on the association of MVPA and interference in preadolescents.
ConclusionsIn preadolescent children, the association of MVPA and inhibitory control differs between sexes. Boys fulfilling the MVPA recommendations by the WHO show better inhibitory control than their less active peers, whereas this pattern is reversed for girls. Neither changes in the inhibitory processing stream, nor the oxygenation response of the frontal cortex underlie this moderating effect of sex. However, preadolescent children with high MVPA show a distinct processing profile, which is characterized by alterations in the allocation of attentional resources and conflict resolution during an inhibitory control task that elicits a stimulus conflict. This profile may either be the result of regular engagement in MVPA or an inhibitory trait that preselects MVPA as a preferred leisure activity.
The study was funded by the Swiss National Science Foundation [32003B_197076].
