


Metacontingency is a conceptual tool that allows behavior analysts to study behavioral processes at a cultural level. The majority of studies on metacontingencies concern cooperative behavior in humans. Skinner (1962), however, proposed some experimental setups for the study of cooperation using pigeons, which were followed by others. The present work was designed to present a setup that allows researchers to investigate metacontingencies using fish as subjects. In addition, it will present a pilot study. The results of the pilot may indicate that an aggregate product emitted by the pairs of fish can be shaped and maintained by its consequences. This enhances the generality of the metacontingency concept, demonstrating that the concept may describe cultural selection in non-human subjects.
La metacontingencia es una herramienta conceptual que permite a los analistas de la conducta estudiar los procesos conductuales a nivel cultural. La mayoría de los estudios sobre metacontingencias abordan la conducta cooperativa en humanos. Sin embargo Skinner (1962) propuso algunas configuraciones experimentales para el estudio de la cooperación utilizando palomas, que han sido seguidas por otros investigadores. Este trabajo fue diseñado para presentar una configuración que permita a los investigadores estudiar metacontingencias utilizando peces como sujetos. Adicionalmente se presentará un estudio piloto. Los resultados de la prueba piloto podrían indicar que un producto agregado emitido por los pares de peces puede moldearse y mantenerse por sus consecuencias. Esto fortalece la generalidad del concepto de metacontingencia, demostrando que éste puede describer selección cultural en sujetos no humanos.
Metacontingency is a conceptual tool that allows behavior analysts to study behavioral processes at a cultural level. This term was first coined by Glenn (1986), and has over the years been defined in different ways (e.g., Martone & Todorov, 2007). Its most recent version can be found in Glenn (2010) (see also Glenn et al., 2016). Glenn's recent formulation describes a metacontingency as a functional relation between (i) interlocking behavioral contingencies (IBCs) and their aggregate product (AP) with (ii) a selecting environment. IBCs are social contingencies defined by the mutual interaction that occurs between at least two organisms, in which each organism's behavior serves as a stimulus for the behavior of other. Aggregate product is an effect on the environment, originating from the coordinated actions of the IBCs (Todorov, 2013).
Experimental analysis in metacontingencies has increased over the last few years, and can be exemplified by the works of Costa, Nogueira, and Vasconcelos (2012), de Carvalho, Couto, Gois, Sandaker, and Todorov (2017), Ortu, Becker, Woelz, and Glenn (2012), Tadaiesky and Tourinho (2012), Vasconcelos and Todorov (2015), Vichi, Andery and Glenn (2009). Despite these recent experimental contributions, there have been experiments within the behavior analytic tradition investigating conditional relations in metacontingencies even before the appearance of the term (Glenn, 1986), and can be exemplified by the works of Arzin and Lindsley (1956), Lindsley (1966), and Schmitt and Marwell (1968). The results of all these experiments suggest “that a cooperative response could be treated exactly as an individual response and there are no new principles involved in this simple cooperative situation” (Lindsley, 1966, p. 475). In this case, cooperative responses are similar to the aggregate product definition (e.g., as used by Toledo et al., 2015). All of the preparations used on those experiments are alike in the sense that humans served as participants. A preparation for the study of cooperation (or metacontingencies – see below) between pigeons was presented by Skinner (1962). In one arrangement, Skinner placed two pigeons in adjacent compartments separated by a glass. On each side of the glass, three buttons were arranged in a vertical row. Pigeons could earn reinforcement whenever both pecked almost instantaneously on a corresponding pair of buttons generating an overlap between the closures of the circuits. It may be possible to translate his experimental preparation into the metacontingency concept: overlapping of the circuits may be defined as the aggregate product, and consequences was delivered by a “selecting environment”. Similar experimental setup using pigeon can be found in Velasco, Benvenuti and Tomanari (2012), and for rats, in Tan and Hackenberg (2015).
The benefits of using non-humans in the experimental analysis of metacontingencies was discussed by Velasco et al. (2012), and so will not be discussed here. Another line of research that investigates cooperative behavior had arisen within biology. Pitman and Durban (2011), for example, have applied naturalistic observation to study group of killer whales. Their studies have shown that groups of these mammals in the Antarctic marine ecosystem, developed cooperative hunting behavior to capture prey. A hunting episode may be summarized as follows: A seal is on an ice floe giving no chance for a whale to capture it by itself. A group of 4–7 whales coordinately respond quite orderly by producing waves that wash seals off the ice floe. This natural phenomenon shows that a group may function as a unit to produce an aggregate product (e.g., wave), and that consequences (capture a prey) may be responsible for selecting it. The present work was proposed to offer a setup that allows researchers to investigate metacontingencies in fish. Fish was chosen to simulate the whales’ natural situation in a laboratory setting, providing the opportunity to assess the role consequences in coordinated responses.
This work has two main folds. One is to present a pilot experiment, which may indicate that an aggregate product emitted by pairs of fish is sensitive to metacontingency relations; the second is to demonstrate how the setup can be automated in order to increase the efficiency and experimental control in future experiments using fish as subjects.
MethodSubjectsFour (∼5cm) Rainbow fish (Melanotaenia boesemani) served as subjects. The experimenter provided a total daily ration of 80 semi-floated micro pellets of Hikari fish food needed to maintain them at free-feeding body weight. This daily ration was derived by determining the amount of food the fish would consume within 15min, given free access. The total amount of food was only provided during and after experimental sessions (the experimenter provided the missing amount of pellets to both fish 5min after finishing each session). Fish were housed in a collective tank which were filtered, temperature controlled (25–28°C), and chemically balanced. They were maintained on a 12-h light-dark schedule.
ApparatusThe experiment took place in a different tank (26cm wide, 52cm deep, and 30cm high) from their living tank. Before starting the experiment, a plastic glass partition was inserted into the tank. There were two similar “housing” (7cm wide, 10cm deep, and 9cm high) at each side of the partition, located at its lower end (see Fig. 1). Micropellet fish food served as reinforcement and was presented on the surface of the water at the opposite side of the “housing”. The micropellets were manually delivered by the experimenter. Clicks of a pen provided stimuli to the occurrence of target responses, which also signaled delivery of reinforcement. The main illumination was provided by a light mounted 3cm above the tank. A camera (Canon) recorded the experimental events. Videos of the session were analyzed after each session.
Target response
The target response was defined as both fish staying at the same time inside each of their “housing”. The experimenter emitted three clicks of a pen whenever both fish were simultaneously inside their “housing”. The clicks, therefore, are defined as the aggregate product. An aggregate product that depended on the behavior of both individuals (both being simultaneously inside their “housing”).
Experimental conditionsThe experiment had two experimental conditions (AB design). The first condition was a usual operant level – baseline. In this condition, target responses were measured in a 15-min session duration. After this unique session, the target response was shaped through differential reinforcement of successive approximation, as a usual shaping procedure. As target responses depended on behavior of both organisms, the shaping procedure considered the behavior of one in relation to the other. For example, no consequences were provided if only one fish approached the “housing” space. However, consequences were provided whenever both fish approached the “housing” space. The difference between the usual shaping procedure, and the one used here, was that the present procedure considered the behavior of one individual relating to the other, rather than individual behaviors. Pair 1 after a session of 45min. For pair 2, shaping was reached after a single session of 15min. From the first session after shaping, the target behavior was maintained at a fixed-ratio 1 (FR 1), for nine consecutive daily sessions of 15min each.
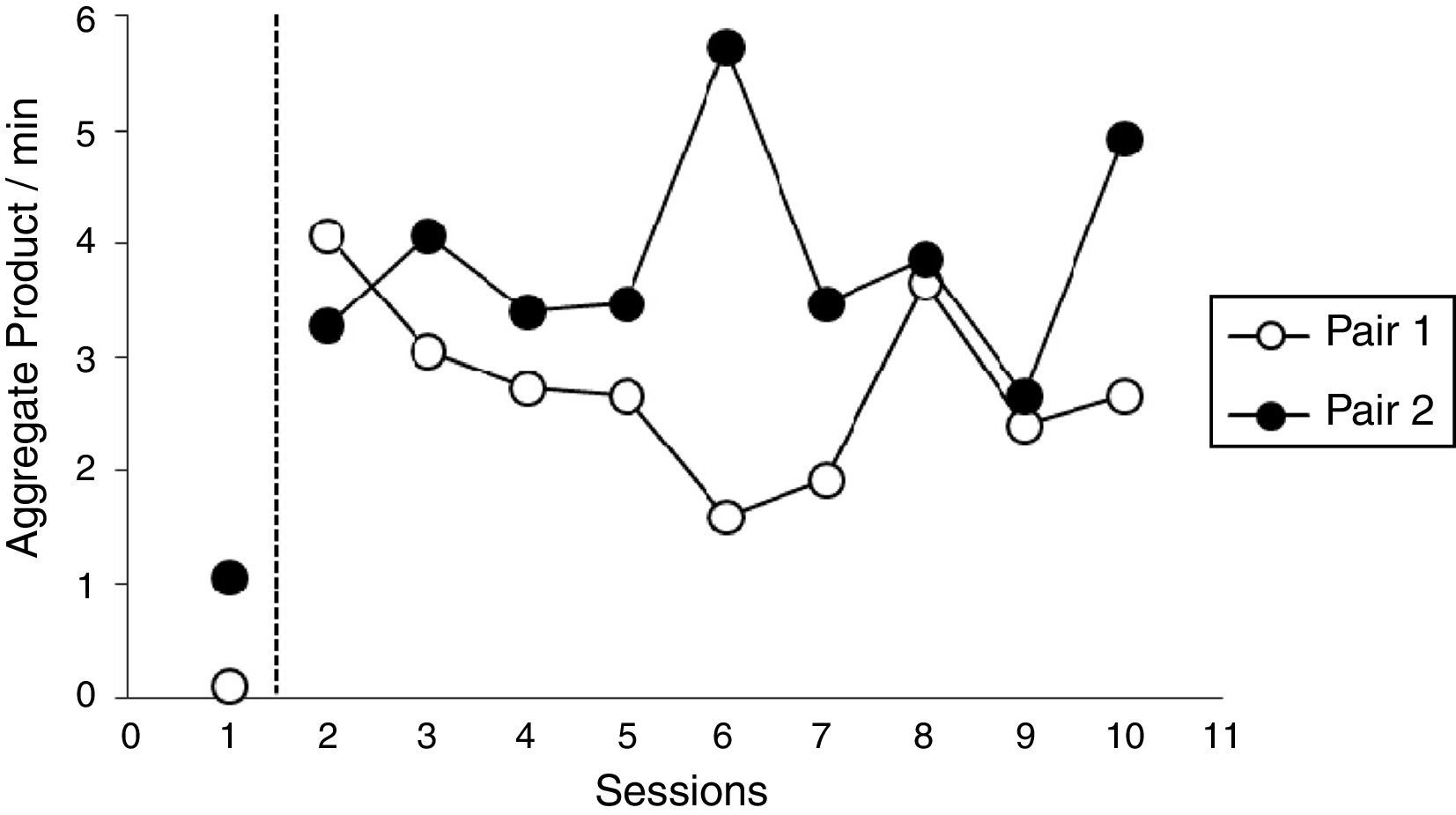
Results and discussionFig. 2 shows the target response rate (aggregate product/min) for all ten sessions, during operant level and FR 1 conditions: rate was calculated by dividing the total responses within a session by session duration (15min). For pair 1, response rate for the operant level was zero, meaning that both fish emitted no target response during the operant level. Responses were maintained at a fairly high rate during the nine following sessions under FR 1 – an average of 2.75. Pair 2 shows a higher response rate during the operant level compared to pair 1, a rate of 1 target response per minute. Under FR, pair 2 showed an average of 3.8 responses per minute, a much higher rate compared to baseline.

The present results demonstrate that a target aggregate product can be shaped and maintained by its consequences. This result is very much in accordance with the metacontingency experimental literature with humans as participants (e.g., Vasconcelos & Todorov, 2015). Vasconcelos and Todorov (2015) procedure, for example, also “followed the usual demonstration of shaping of an operant with low relative frequency of occurrence: initial determination of operant level followed by differential reinforcement of successive approximations to the target response”. Their results shows a consistent selection of coordinated responses as function of a contingent consequence. The present results enhance the generality of their findings by showing that cultural selection is not a special case for humans (e.g., de Carvalho & Sandaker, 2016). It has been shown that when consequences are delivered to coordinated responses of fish, selection for these responses takes place.
Studies with fish may be interesting to evaluate the effects of consequences in a metacontingency on the establishment of conditioned social stimulus. In order to measure social stimulus control one can do the following. After an aggregate product has been established through shaping, one fish of the pair is detained inside the “housing”, while the other is placed inside the tank afterwards. The latency for the second fish to meet the first in the “housing” is assessed. This effect would be measured after the same situation had previously been evaluated to the shaping procedure. As latency diminishes after conditioning, one may suggest that consequences in metacontingencies develop conditioned social stimulus control. This seems to happen in humans (e.g., de Carvalho et al., 2017), but we do not know exactly in what extent verbal behavior (e.g., by following rules) has to do with it (see also Velasco et al., 2012). Therefore, the present setup may be useful for controlling this kind of variable. The preparation is one example that shows how an established aggregate product permits researchers to use it as baseline to investigate the effects of different variables on coordinated responding (e.g., schedules of reinforcement, reinforcement magnitude, antecendent stimuli, punishment, and so on)
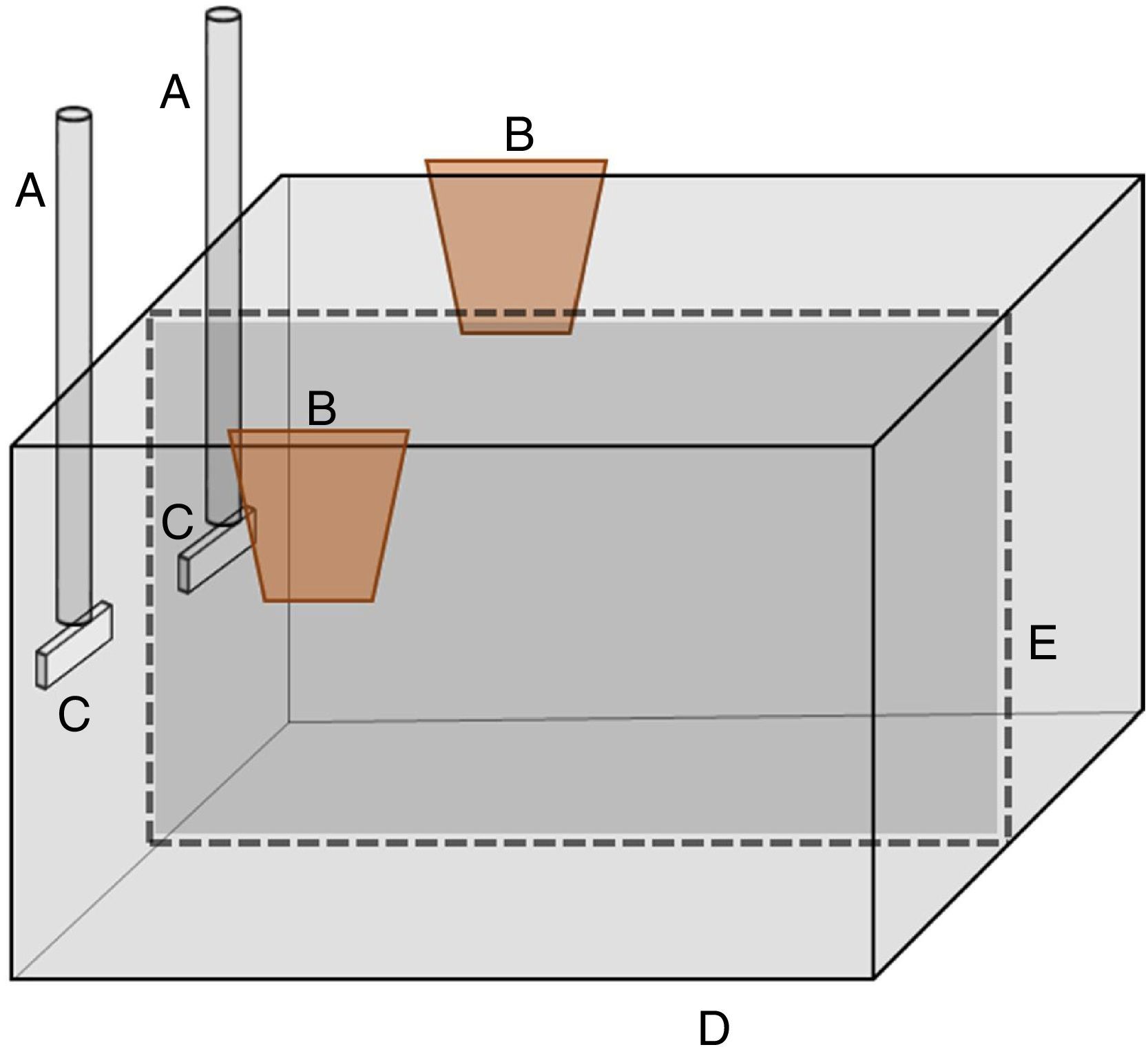
Automatization of the setupThere are in the behavioral analytic literature examples on how experimental preparations can be automatized based on interface and computer software (e.g., Escobar & Lattal, 2010; Escobar & Perez-Herrera, 2015; Escobar, Hernández-Ruiz, Santillán, & Perez-Herrera, 2012; Iversen, 2002, 2008). The automatization of the present preparation can surely be made through the interface and Visual Basic programming as presented by Manabe, Dooling, and Takaku (2013). Thus, I would suggest that researches assemble the aquarium as shown in Fig. 3. The aquarium (D) would contain two response keys (C) two feeders (B) a plastic glass partition (E). (E) would eliminate any kind of undesirable interactions such as fighting. The automatization of the present preparation would allow behavioral scientists to investigate the effects of many variables that pertains to the cooperating behavior of two fish.
. (C) are the response keys, (A) where the sensors are placed, (E) is a plastic glass partition, and (B) is the feeders.")
Automatic programming seems not to be very important when evaluating the effects of a simple metacontigency situation as in the pilot. However, the more interesting variables are included in the analysis, the more complex the experimental preparation becomes. Ferster and Skinner (1957) referring to the usefulness of automatic programming in works on schedules of reinforcement, stated that “the reaction time and variability of the human operator introduce errors in programming which defeat the purpose of such research. The required level of precision can be achieved only through the use of mechanical or electrical devices”. As fish becomes a recurrent subject in the experimental analysis of metacontingency, automatic programming becomes a required condition.
The author is greatful to three anonymous reviewers and Johan Viklund for suggestions that improved the manuscript.
Peer review under the responsibility of Asociación Mexicana de Comportamiento y Salud.