






Las características biológicas de los seres vivos se transmiten a la descendencia a través de los gametos. Estos llevan la información genética de ambos padres codificada en el ADN y son las únicas células que pasan físicamente de padres a hijos a lo largo de las generaciones. Para ello, los gametos deben estructurarse de modo que al unirse el paterno y materno formando el zigoto, se mantenga el número de cromosomas de la especie humana y sus demás características. El proceso de la formación de los gametos por el que adquieren la mitad del número de cromosomas (uno de cada par cromosómico) se denomina meiosis y constituye la base del desarrollo embrionario normal.
The biological characteristics of human beings are transmitted to the descendents through the gametes. These carry the genetic information of both parents coded in the DNA and are the only cells that physically pass from parents to children over the generations. Thus, the gametes should be structured in such a way that when the paternal and maternal ones are joined, forming the zygote, the number of chromosomes of the human species and their other characteristics are maintained. The gamete formation process by which half of the chromosome number is acquired (one of each chromosome pair) is called meiosis, and makes up the basis of the normal embryonic development.
Las células madres de los gametos, al igual que las que constituyen los diferentes tejidos, inician su diferenciación durante etapas muy tempranas del desarrollo embrionario. En el ser humano, hacia los días 16–18 después de la fecundación, ciertas células epiblásticas del embrión comienzan su diferenciación como células germinales primordiales (CGP) y, hacia el día 21–22 de gestación, cuando se ha formado el embrión trilaminar, las CGP se localizan en la pared de la parte caudal del saco vitelino. Desde ahí, empiezan la migración hacia las primitivas glándulas sexuales del embrión, a las que llegan entre la 4.a y 5.a semana tras la fecundación (que corresponden a la 6.a y 7.a contando desde el primer día de la última regla). Desde ese momento hasta el final del 2.° mes desde la fecundación, las CGP inician una intensa multiplicación celular mediante sucesivas mitosis. A partir de ese momento, y durante el 3.er mes los procesos de desarrollo de las CGP van a diferir dependiendo del sexo del embrión.
En un feto femenino, durante el 3.er mes de gestación las primitivas ovogonias inician el proceso de meiosis con la duplicación del ADN y la progresión de las primeras etapas de la profase de la meiosis, hasta el estado de diplotena, en el que se paraliza el proceso. Esta paralización se produce una vez que finaliza el intercambio de segmentos cromosómicos entre las parejas de cromosomas homólogos (paterno y materno), llamado recombinación (crossing-over, o cross-over). A partir de este momento, el proceso de meiosis permanecerá paralizado hasta la pubertad de la niña, hacia los 11–14 años de vida, cuando se reanuda el proceso con la progresión de la meiosis para formar los gametos femeninos (fig. 1A-1). La meiosis femenina tiene también otra característica, que consiste en que sus dos divisiones de las anafases I y II son asimétricas. Así, durante la anafase I, ocurre la separación de los cromosomas paterno y materno, que forman las 23 parejas de cromosomas, hacia los 2 polos del huso. Sin embargo, en la fase final (telofase I) se forman dos células de diferente tamaño (fig. 1A-2), que son: el ovocito secundario, que se queda prácticamente con todo el citoplasma junto con un cromosoma de cada pareja (23 cromosomas, cada uno con 2 moléculas de ADN —llamadas cromátidas—), y otra célula más pequeña llamada 1.er corpúsculo polar (CPI), que está formada por los otros 23 cromosomas dobles y una pequeña cantidad de citoplasma. El proceso por el que se estructura esta división asimétrica es complejo, y se basa en que el huso de la primera división de la meiosis se sitúa fuera del centro celular. Una localización que debe establecerse de forma que uno de los polos del huso esté muy cerca de la membrana celular. De este modo, se favorece que solo se excluyan los cromosomas de ese polo con una pequeñísima cantidad de citoplasma. Para ello, se produce un fascinante mecanismo de interacción entre los microtúbulos del huso y filamentos de actina y miosina del citoplasma, que obligan al huso a disponerse fuera del centro celular1–2.

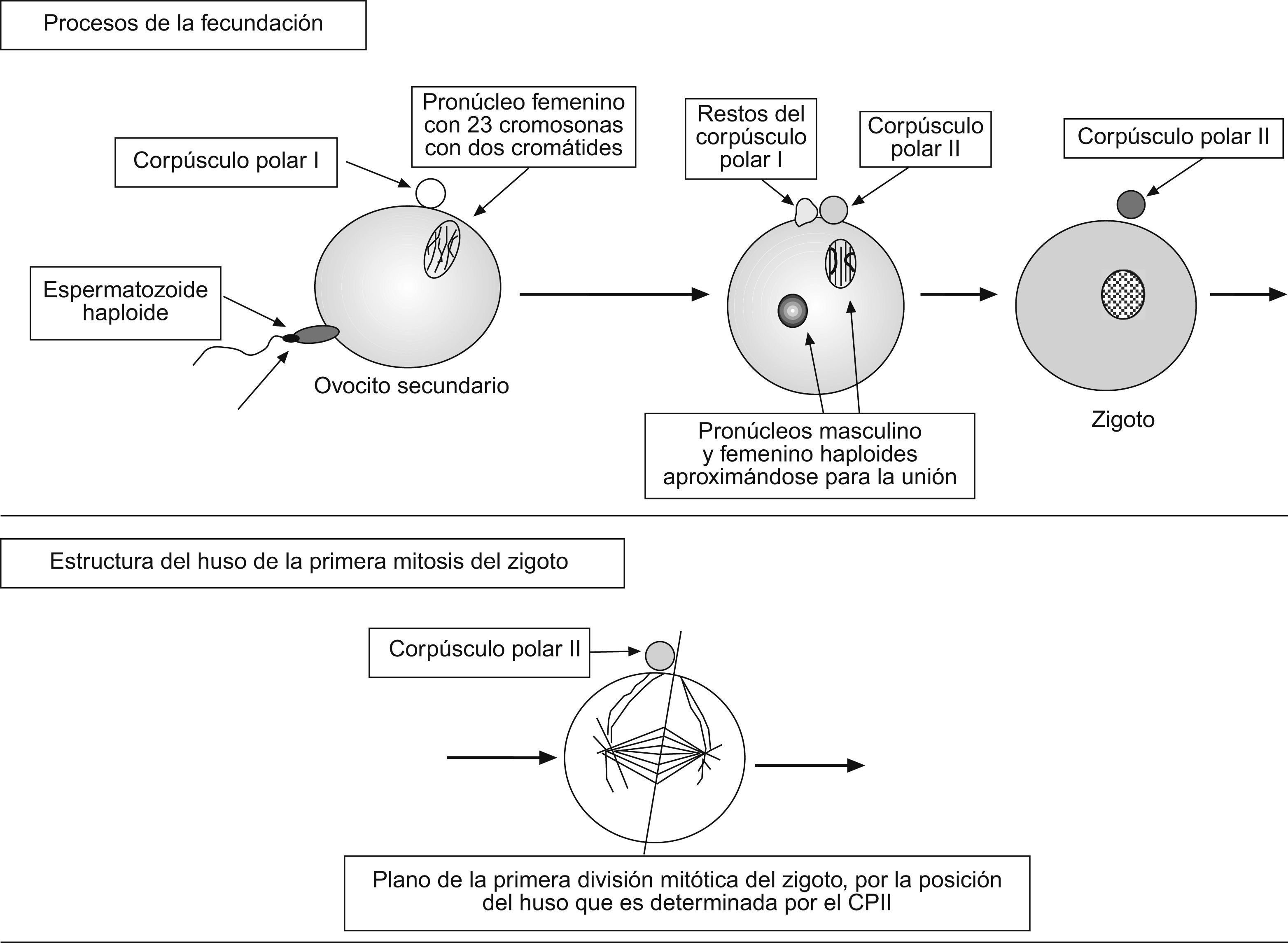
Como se indica en la fig. 1A-2, el CPI queda unido a la membrana del ovocito primario y marca el lugar de la siguiente división de la meiosis, que también es asimétrica y que ocurre solo en el ovocito secundario y no en el CPI. Su resultado es la formación del óvulo y el 2.° corpúsculo polar (CPII), que se excluirá cerca de donde está situado el CPI (fig. 1A-3). No obstante, la inmensa mayoría de los gametos femeninos nunca terminan el proceso de meiosis, porque lo que se ovula cada 28 días es el ovocito secundario junto con el CPI y solo si hay fecundación finaliza el proceso de meiosis (fig. 2). De hecho, es la entrada del gameto masculino la que estimula la finalización de la segunda anafase de la meiosis, con la exclusión del CPII (que tiene una importante función durante las primeras fases del desarrollo embrionario, como se expondrá más adelante). Por último, otra característica de los gametos femeninos, es que su formación cesa cuando la mujer tiene una edad de entre 45–50 años, con la menopausia.

La formación de los gametos masculinos sigue el mismo proceso de meiosis pero, al contrario de lo que ocurre en los gametos femeninos, las 2 divisiones de la meiosis masculina forman células de igual tamaño y composición (divisiones simétricas). Además, en el 3.er mes de embarazo de los fetos masculinos, al contrario de lo que ocurre en los femeninos, las pre-espermatogonias no se multiplican, sino que entran en un periodo de latencia en el que van a permanecer prácticamente hasta la pubertad del niño. En la pubertad (fig. 1B), inician un intenso proceso de multiplicación celular mediante mitosis sucesivas y continuas, que forman las espermatogonias y estas los espermatocitos primarios, en los que se inicia el proceso de la meiosis (fig. 1B-1). En esta, se producirán 4 espermatidas con la mitad del número de cromosomas (fig. 1B-3), las cuales seguirán un proceso de transformación celular para adquirir la estructura de espermatozoides maduros. Esta última transformación es un proceso complejo, que no solo incluye la pérdida del citoplasma y la adquisición del flagelo, sino una importante modificación de la estructura de la cromatina del núcleo. Básicamente se trata de cambiar la configuración de la estructura de la cromatina para que el ADN se compacte mucho más, de forma que se pueda incluir en el pequeño espacio del núcleo del espermatozoide (que es la célula más pequeña de los seres humanos).
La cromatina de todas las células está constituida por un octágono de 4 histonas en las que se enrolla la cadena de ADN, formando unas estructuras llamadas nucleosomas (fig. 3A). Ese enrollamiento permite condensar la molécula del ADN en el núcleo. Sin embargo, el espermatozoide es la única célula humana que no tiene nucleosomas, porque durante su formación substituye las histonas de los nucleosomas por protaminas. Estas protaminas facilitan el plegamiento de la molécula del ADN en forma de ondulaciones (fig. 3B), lo que permite una mayor compactación del ADN.

Otra diferencia entre el esperma y los ovocitos es que la transformación de espermatogonias en espermatozoides es un proceso continuo desde la pubertad y durante prácticamente toda la vida del hombre adulto. Esto implica que siempre se está produciendo síntesis (duplicación) de ADN para la formación de los gametos, lo que no ocurre en los gametos femeninos (ovocitos), cuya duplicación del ADN ocurrió durante el desarrollo embrionario. Por otra parte, mientras que la profase de la meiosis en la mujer tiene un proceso de formación que puede durar hasta los 50 años de vida posnatal, en las células masculinas dura 22 días, y la de todo el proceso de formación de los espermatozoides unos 65–70 días, que es el tiempo del recambio espermático.
FecundaciónPor todo lo expuesto en los apartados anteriores, en el momento de la fecundación los 2 gametos son muy diferentes, fundamentalmente en los siguientes aspectos.
- 1.
El gameto masculino es ya haploide (tiene 23 cromosomas con una sola molécula de ADN o cromátida). Por el contrario, el femenino está en fase de ovocito secundario (fig. 1A-1) y, por tanto, en reposo en la segunda metafase de la miosis. Es decir que tiene 23 cromosomas (1 de cada pareja), pero los 23 cromosomas siguen teniendo doble cantidad de ADN (2 moléculas de ADN o 2 cromátidas).
- 2.
El gameto masculino está prácticamente limitado al contenido del ADN (23 cromosomas con una cromátida). Sin embargo, el ovocito secundario tiene un gran citoplasma con todas las estructuras duplicadas, ya que el CPI apenas se quedó con una parte muy pequeña del mismo. Además, durante su proceso de maduración hasta la ovulación, sintetiza una gran cantidad de proteínas y factores de transcripción maternos (se han identificado más de 14.000, de familias distintas). Estos factores son esenciales para regular los procesos de transición de los 2 pro-núcleos, paterno y materno, para que la formación del zigoto sea correcta y para que se sucedan, en forma adecuada, todas las etapas iniciales del desarrollo embrionario hasta la implantación3–5.
En el momento de la fecundación (fig. 2), con la entrada del gameto masculino en el citoplasma del ovocito, este finaliza la 2.a división de la meiosis con la expulsión del CPII en forma similar a la exclusión del CPI. Además, el CPII queda anclado en la membrana del zigoto, cerca del CPI y, desde esa posición va tener mucha importancia para formación de los primeros blastómeros.
Primera división del zigotoBasados en estudios sobre ratones (cuyos mecanismos son muy similares a los de los seres humanos), se está avanzando mucho en el conocimiento de los procesos que ocurren durante las primeras fases embrionarias hasta la implantación. Se ha demostrado que el CPII determina la posición de los polos del huso de la primera mitosis por la que el zigoto dará lugar a los 2 primeros blastómeros. De hecho, a través del anclaje de fibras de los 2 polos del huso en la zona del CPII (Fig. 2), se determina la posición de la metafase, de forma que el eje de separación de los cromosomas hacia los polos durante la anafase sea perpendicular al CPII. Es más, observaciones muy recientes han mostrado que las células resultantes de esta primera división no son idénticas, y que esa diferencia va a determinar las células que formarán el polo embrionario y las que darán lugar al trofoblasto6.
En todas las fases del desarrollo embrionario y fetal, tanto las CGP, su evolución y maduración en gametos, como de las células somáticas que constituirán el cuerpo del futuro hijo, se producen numerosos procesos genéticos y epigenéticos, que determinan su desarrollo final. Estos aspectos los desarrollaremos en otro capítulo de este curso.
