



La toma de decisiones (TD) puede definirse como la selección de una alternativa dentro de un rango de opciones existentes, considerando los posibles resultados de las selecciones realizadas y sus consecuencias en el comportamiento presente y futuro. Tradicionalmente, se ha afirmado que desde el punto de vista anatómico la base neural fundamental de este proceso lo constituye la corteza prefrontal (CPF); sin embargo, nuevos estudios validan la hipótesis de la existencia una compleja red neural que incluyen estructuras tanto corticales como subcorticales.
ObjetivoLa presente revisión tiene como objetivo resumir la evidencia sobre las bases anatómicas relacionadas con el proceso de toma de decisiones tomando en consideración la información disponible hasta la actualidad, que valida la existencia de una compleja red neural que sirve de soporte a este complejo proceso neuropsicológico.
DesarrolloLa evidencia contemporánea indica que dentro de las bases neurales de la TD se encuentran regiones de la CPF como la corteza orbitofrontal, dorsolateral y el giro cingulado anterior. Además, el proceso es asistido por regiones subcorticales, como la amígdala, el hipocampo y el cerebelo.
ConclusionesLos resultados hasta el momento demuestran la importancia de las estructuras corticales y subcorticales en la toma de decisiones. Las bases neurales de la TD consisten en una compleja red neural con conexiones cortico-corticales y cortico-subcorticales, que incluyen tanto las subdivisiones de la CPF como las estructuras límbicas y el cerebelo.
Decision-making is the process of selecting a course of action from among 2 or more alternatives by considering the potential outcomes of selecting each option and estimating its consequences in the short, medium and long term. The prefrontal cortex (PFC) has traditionally been considered the key neural structure in decision-making process. However, new studies support the hypothesis that describes a complex neural network including both cortical and subcortical structures.
ObjectiveThe aim of this review is to summarise evidence on the anatomical structures underlying the decision-making process, considering new findings that support the existence of a complex neural network that gives rise to this complex neuropsychological process.
DevelopmentCurrent evidence shows that the cortical structures involved in decision-making include the orbitofrontal cortex (OFC), anterior cingulate cortex (ACC), and dorsolateral prefrontal cortex (DLPFC). This process is assisted by subcortical structures including the amygdala, thalamus, and cerebellum.
ConclusionsFindings to date show that both cortical and subcortical brain regions contribute to the decision-making process. The neural basis of decision-making is a complex neural network of cortico-cortical and cortico-subcortical connections which includes subareas of the PFC, limbic structures, and the cerebellum.
La toma de decisiones (TD) puede definirse como la selección de una alternativa dentro de un rango de opciones existentes, considerando los posibles resultados de las selecciones realizadas y sus consecuencias en el comportamiento presente y futuro1. Cada año el 40% de todas las muertes a nivel global son a causa de déficit en la TD en los componentes básicos de la autorregulación2. Este complejo proceso ha sido estudiado por diversas disciplinas científicas (psicología cognitiva, economía, ciencias de la computación y neuropsicología), tomando como referente variados modelos teóricos3-6. En este sentido, resulta de especial interés para los neurólogos y neuropsicólogos la hipótesis del marcador somático planteada por Damasio, la cual ofrece un referente teórico y metodológico al establecer las bases de la relación entre las estructuras corticales y subcorticales a la hora de decidir, así como las alteraciones relacionadas con ciertos daños cerebrales7,8.
Desde un punto de vista anatómico, se ha considerado la corteza prefrontal (CPF) como la región cerebral de mayor importancia a la hora de delimitar las bases neurales subyacentes a este proceso. Estructuralmente, la CPF está compuesta por 3 subregiones cerebrales: la corteza orbitofrontal (COF), el córtex cingulado anterior (CCA) y la CPF dorsolateral (CPFDL), las que en estrecha relación con otras regiones, como el tálamo, la amígdala y los ganglios basales, garantizan un adecuado proceso de TD9. Estas estructuras se organizan en 3 circuitos fundamentales que guardan relación con la TD: 1) la COF y las conexiones límbicas, estrechamente relacionadas con las recompensas y las decisiones de base emocional; 2) la corteza dorsolateral del córtex prefrontal (CPFDL) especializada en la integración de múltiples fuentes de información, y 3) la corteza cingulada anterior (CCA) implicada en el manejo de información conflictiva y el procesamiento del feedback asociado a las decisiones10. Además, el córtex prefrontal posee conexiones con otras regiones subcorticales que guardan relación con el funcionamiento de las estructuras mencionadas anteriormente11.
El presente trabajo tiene como objetivo resumir las bases anatómicas relacionadas con el proceso de TD tomando en consideración la evidencia disponible hasta la actualidad, que valida la existencia de una compleja red neural que sirve de soporte a este complejo proceso psicológico.
DesarrolloCorteza orbitofrontal y toma de decisionesLa CPF es una región cerebral de gran importancia para la TD. Entre sus estructuras se encuentras el córtex orbitofrontal (COF), el giro cingulado anterior (CCA) y la CPFDL. El COF es una estructura importante para las decisiones relacionada con los incentivos positivos (ganancias), así como las situaciones emocionales asociadas a los resultados de las selecciones (feedbacks)11. El COF posee conexiones con otras estructuras relacionadas con la TD (p. ej., la CPFDL y la CCA). Además, posee una elevada conectividad bidireccional con el córtex de asociación temporal, la amígdala y el hipocampo; ejerciendo una fuerte influencia en el procesamiento emocional12.
Funcionalmente, la COF se organiza de forma que la porción medial se especializa en la decodificación las recompensas, mientras que las porciones laterales evalúan los castigos13. En este sentido, las recompensas asociadas a reforzadores primarios (p. ej., el sabor) se decodifican en las regiones posteriores de la COF, mientras que los reforzadores secundarios más complejos (p. ej., el dinero), se decodifican en las regiones anteriores de esta estructura14. Usualmente, los pacientes con daño en la COF se caracterizan por presentar 2 tipos de déficit fundamentales. En primer lugar, estas personas muestran incapacidad para alterar sus decisiones en relación con una tarea, a pesar de la aparición de resultados desventajosos. En segundo lugar, el daño en la COF se relaciona con alteraciones en la empatía y la teoría de la mente9.
A nivel celular (en estudios con primates), se ha demostrado que, asociada a las recompensas, las neuronas de la COF muestran una activación elevada, particularmente cuando el primate anticipa elevadas recompensas en oposición a premios pequeños15. Por el contario, la activación de esta región disminuye considerablemente cuando asociado a las decisiones se anticipan elevadas pérdidas (castigos). Este nivel de excitación-inhibición de las neuronas de la región orbitofrontal se encuentra modulado además por la presentación previa de los estímulos y la inmediatez de las recompensas asociados a los mismos16.
En el caso de los estudios realizados con humanos, las evidencias se sustentan en lo fundamental en el análisis de pacientes con daño frontal durante la ejecución de tareas cognitivas. En el caso de las investigaciones que han empleado el Iowa Gambling Task (IGT)17 y el Cambridge Gambling Task (CGT)18 se ha confirmado la presencia de dificultades para alterar el patrón de decisiones inicial sobre la base de las contingencias que aparecen durante el transcurso del proceso decisional. Este déficit en las decisiones de base emocional se asocia fundamentalmente a las fuertes conexiones existentes entre la COF, la ínsula y el sistema límbico en general (hipocampo, amígdala, etc.)19.
Corteza prefrontal dorsolateral y corteza cingulada anteriorPor otra parte, una TD óptima necesita de la participación de otras estructuras corticales de elevada especialización. Se hace referencia especialmente a la CPFDL y a la corteza cingulada anterior (CCA)20. La CPFDL ocupa las regiones laterales y superiores de los lóbulos frontales y se organiza a través del eje dorso-ventral. La porción dorsal de la CPFDL se encarga de la monitorización de la memoria de trabajo y la región ventral de la recuperación de la información almacenada en las regiones posteriores de asociación cortical11,21. De igual forma que la COF, la CPF ventromedial posee fuertes conexiones límbicas y cortico-corticales, especialmente a través de las regiones temporales, parietales y occipitales21,22.
En el caso de la corteza cingulada anterior (CCA) ocupa la porción medial del córtex prefrontal. Esta región posee conexiones cortico-corticales con la COF y la CPFDL, así como proyecciones subcorticales hacia el núcleo acumbens. Ha quedado demostrado que la porción anterior de la CCA guarda relación directa con la depresión relacionándose con la TD con base emocional23. El papel fundamental de la CCA en la TD se relaciona con la modulación de otras regiones prefrontales como la COF y la CPFDL. Esta función es ejercida fundamentalmente a través del análisis de situaciones que resultan ambiguas o conflictivas24. Además, la CCA participa en la optimización de las decisiones futuras sobre la base de las contingencias previas recibidas a lo largo del proceso de selección de opciones25. Esta característica ha sido comprobada a través del uso de resonancia magnética funcional (fMRI) mostrando un aumento de la activación de la CCA en los periodos en los que se reducen las recompensas o cuando se incrementa la retroalimentación negativa asociada a selecciones incorrectas26.
Tálamo y ganglios basales: implicaciones en la toma de decisionesNo obstante, el adecuado funcionamiento de las regiones prefrontales no depende únicamente de las estructuras frontales como la COF, la CPFDL y la CCA, sino que además requiere de las conexiones establecidas a través de la materia blanca y las estructuras subcorticales como el tálamo, los ganglios basales y el cerebelo, con las cuales la CPF está interconectada. Por esta razón, un daño en estas regiones puede alterar considerablemente el proceso de TD9. Es conocido que todas las regiones prefrontales involucradas en la TD envían proyecciones hacia el núcleo caudado y al putamen a través del fascículo subcalloso de Muratoff o de la cápsula externa27,28. En respuesta a este proceso, los ganglios basales orientan la información hacia el tálamo, el cual envía estímulos de retroalimentación (feedbacks) hacia las regiones corticales originales29.
Por otra parte, la COF y la CCA poseen proyecciones hacia el estriado ventral30, por lo cual un daño en esta región provocaría dificultades en la TD adaptativa, ya que genera síntomas de desinhibición, disminución de respuestas asociadas a las recompensas y los comportamientos lábiles31. Como se explicó anteriormente, los ganglios basales orientan la información hacia el tálamo, el cual lo proyecta nuevamente hacia el neocórtex. El núcleo mediodorsal está recíprocamente conectado con la COF y la CPFDL, la amígdala y el córtex olfatorio32. Las lesiones de este núcleo talámico producen alteraciones en la TD puesto que provocan apatía, abulia y desinhibición33.
Amígdala, cerebelo y toma de decisionesLa amígdala es otra región subcortical involucrada en la TD. Esta estructura del sistema límbico desempeña un papel fundamental en la decodificación emocional de los estímulos ambientales34. La importancia de la amígdala en relación con el proceso de TD se debe en lo fundamental a su participación en los mecanismos de recompensa. Esta función es ejercida a través de su interacción con el estriado ventral facilitando las asociaciones estímulo-recompensa35-37.
Estudios metabólicos en los que se ha empleado tomografía por emisión de positrones (PET, en inglés) han reportado una considerable activación amigdalina en contextos que implican recompensas potenciales38. Esto representa un importante elemento en los procesos de TD en los que se necesita continuamente realizar balances de ganancias y pérdidas para la selección de una opción sobre otra. Otros estudios que han empleado también han comprobado una importante activación de esta región en contextos de recompensas probables39. Sin embargo, esta activación parece estar referida más a la activación neural que provoca el estímulo (arousal) que al valor del mismo (valencia, por ejemplo, agradable/desagradable). A esta conclusión se ha arribado luego de comprobar los niveles de actividad registrados por la amígdala ante estímulos de recompensa vs. castigo, en los que no se mostraron diferencias en su actividad en relación con la valencia40,41.
Este resultado, unido a la conocida peculiaridad de la amígdala de poseer un acelerado nivel de habituación frente a los estímulos42, permite comprender el papel que desempeña esta región frente a situaciones de decisión novedosas que requieren evaluaciones rápidas que garanticen respuestas adaptativas en breves periodos. Sin embargo, su función podría estar subordinada esencialmente a decisiones de fuerte contenido emocional con las cuales es sujeto decisor no posee familiaridad. Este argumento gana respaldo en estudios que han demostrado una hipoestimulación de esta estructura en tareas de TD que han empleado fMRI43.
Otra región que ha demostrado desempeñar un papel importante en la TD es el cerebelo. La función del cerebelo ha sido tradicionalmente atribuida al desarrollo de la postura, el control motor y la coordinación. Sin embargo, un número importante de estudios realizados a partir de 1990 han demostrado que las funciones de esta estructura no se reducen a estos procesos44. Por ejemplo, las áreas posteriores del cerebelo participan en funciones de alto nivel, como la memoria de trabajo, el procesamiento lingüístico, el análisis de la información espacial y la regulación emocional45. El cerebelo constituye también un nodo integral en el circuito cortico-subcortical con las estructuras prefrontales, modulando su funcionamiento integral28. Además desempeña un importante papel en el desarrollo de los procesos atencionales46.
Un estudio reciente comparó el rendimiento en la TD en pacientes con daño cerebelar con 2 grupos adicionales, uno con daño frontal y otro de control. Los resultados mostraron que los pacientes con daño en el cerebelo mostraron un rendimiento más bajo en la TD en comparación con los controles, aunque superior si se comparaba con el grupo con daño frontal44. Esta evidencia se suma a los datos obtenidos por otras investigaciones que han relacionado la TD con el funcionamiento del cerebelo47. De acuerdo con estos resultados, el cerebelo pertenece a un grupo de regiones cerebrales que se activan tanto en la TD bajo incertidumbre, como en situaciones de certidumbre, desempeñando un papel fundamental en las representaciones internas de los eventos de incertidumbre, facilitando así las predicciones de las consecuencias futuras, así como de los procesos inductivos.
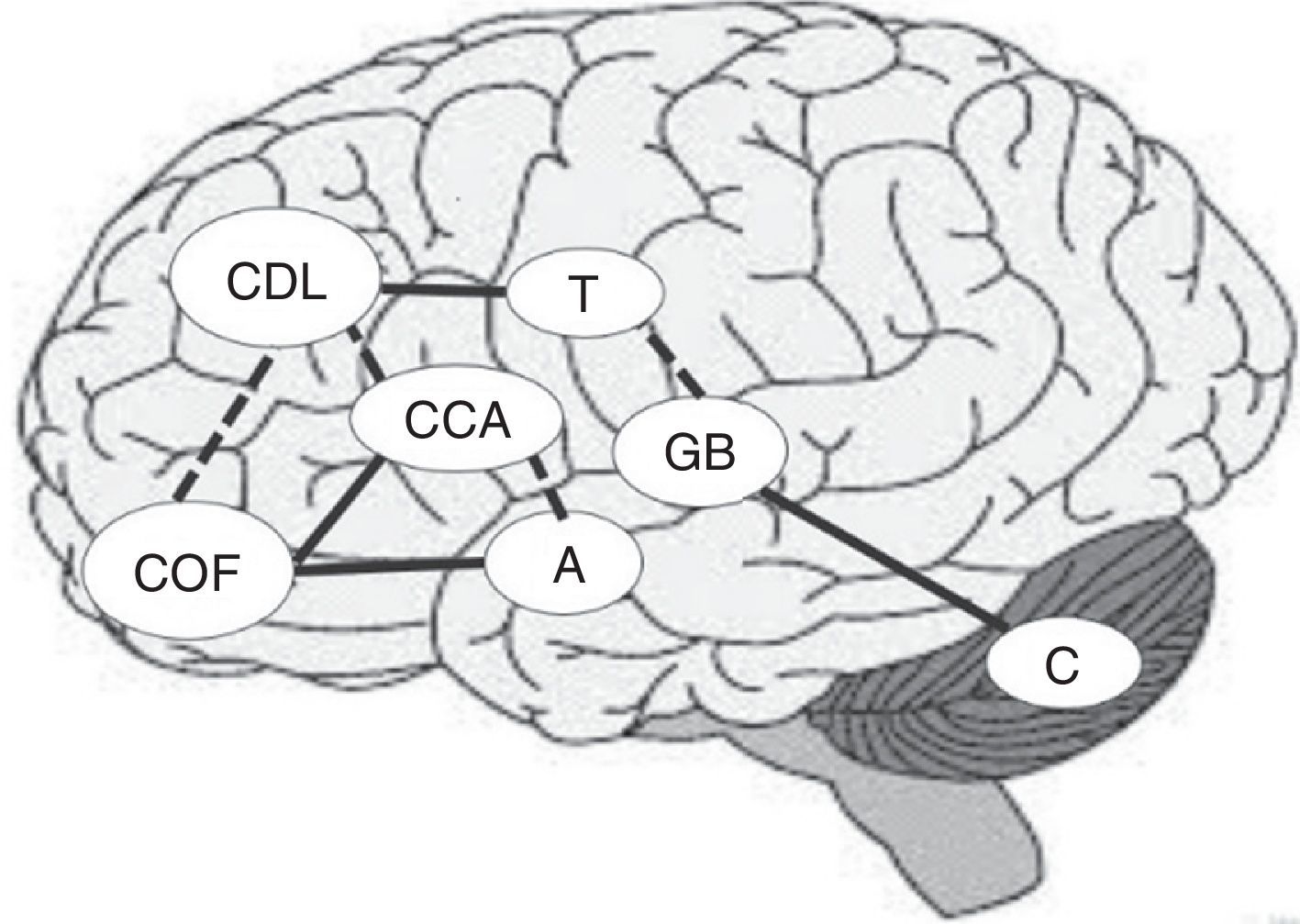
Es conocido que junto al córtex prefrontal derecho y los ganglios basales, el cerebelo está implicado en las representaciones internas de las distancias geográficas y de los eventos temporales48,49. Por esta razón, si se encuentra dañada la capacidad para establecer conexiones temporales entre las acciones y sus consecuencias, también podrían mostrarse dificultades para aprender de las experiencias, dificultando el rendimiento en la TD al no poder identificar adecuadamente las decisiones ventajosas y desventajosas44. La figura siguiente muestra las principales áreas implicadas en el proceso de TD y las interconexiones entre las regiones teniendo en cuenta los distintos circuitos abordados. Con líneas continuas se muestran las regiones con mayor grado de conectividad y con líneas discontinuas aquellas que participan fundamentalmente en situaciones de incertidumbre (fig. 1).
Daño cerebral y toma de decisiones: disfunciones en patologías neurológicas
A través del estudio sistemático de pacientes con alteraciones neurológicas, se ha podido comprobar, en un número significativo de patologías, alteraciones en la TD. Especialmente, se han encontrado disfunciones decisionales en pacientes con daño cerebral traumático (DCT)50,51, en la demencia frontotemporal (variante comportamental)52, en la enfermedad de Parkinson y en pacientes con dependencia de las drogas53, pacientes con daño frontal y cerebral causado por accidentes cerebrovasculares44 en pacientes autistas y en individuos con diagnóstico de esquizofrenia54,55, en pacientes con hemorragia subaracnoideas56 y también en el trastorno bipolar57.
En el caso específico de la demencia frontotemporal, se ha comprobado que, unidas a los síntomas de desinhibición comportamental, las conductas perseverativas y la hiperoralidad21, también aparecen alteraciones en la TD morales a partir de la dificultad de estos pacientes para apreciar las consecuencias sociales de sus acciones52. Esta peculiaridad se debe fundamentalmente a que en esta patología, a pesar de encontrase preservada la CPFDL, existe un marcado deterioro en la COF, la cual, como se ha abordado anteriormente, se encarga del procesamiento emocional de las situaciones de decisión y de la inhibición conductual9.
Por otra parte, los pacientes aquejados de DCT o quirúrgico también presentan marcadas alteraciones en la TD. En estos pacientes son frecuentes los traumatismos en las regiones temporales y también en la COF9, lo cual provoca usualmente daños en la materia blanca que provocan lesiones axonales difusas. En los pacientes con DCT, las alteraciones en la TD se expresan comportamentalmente a través de una marcada impulsividad y dificultades en la planificación de acciones que se manifiesta a través de conductas agresivas58.
Evaluación clínica de la toma de decisionesEn lo referente al diagnóstico, varias son las pruebas que pueden emplearse en la evaluación de la TD para la detección de alteraciones neurológicas y neuropsicológicas, tanto en pacientes con daño cerebral como también en personas aparentemente sanas. Dentro de las más empleadas se encuentran el Balloon Gambling Task59, las tareas de probabilidad de selección de riesgos60, así como el IGT y el Columbia Card Task61-63. Igualmente, y teniendo en cuenta los objetivos de la evaluación, puede emplearse además el test de cartas de Wisconsin (WCST)64-67 y el Risky Choice Task68.
Especialmente, se debe destacar el IGT, herramienta que ha demostrado una elevada sensibilidad para la detección de daños en las estructuras prefrontales y también en zonas subcorticales implicadas en la TD17,43. A partir de su empleo, se han detectado dificultades decisionales en la esquizofrenia69,70, el déficit de atención con hiperactividad71, alteraciones en el control de impulso con agresividad72 y la enfermedad de Huntington73, entre otras muchas alteraciones. Como ventaja adicional, esta prueba cuenta con una versión especialmente diseñada para niños denominada Tarea del Burrito Hambriento (Hungry Donkey Task), con un principio de funcionamiento similar al IGT pero ajustada a las peculiaridades psicológicas y neuropsicológicas de los menores74. Finalmente, el IGT se destaca por ser la prueba de evaluación de TD que mayor respaldo empírico ha recibido a través del uso de las neuroimágenes, herramientas neurofisiológicas y escalas comportamentales75.
ConclusionesLa evidencia acumulada hasta el momento demuestra la importancia de las estructuras corticales y subcorticales en la TD. Las estructuras del neocórtex imprescindibles para el funcionamiento adaptativo de este proceso son la COF, el córtex cingulado anterior y la corteza prefrontal dorsolateral. En el caso de las regiones subcorticales de importancia para la TD se encuentran estriado ventral, los ganglios basales, la amígdala y el cerebelo. Desde un punto de vista funcional, se ha demostrado la existencia de un circuito característico en situaciones de decisión bajo incertidumbre. Las interconexiones detectadas implican al córtex prefrontal dorsolateral (relacionado con la memoria de trabajo), la ínsula y el córtex cingulado posterior (asociado a la representación de estados emocionales), el córtex prefrontal mesial orbitofrontal y ventromedial (en la integración de los procesos descritos anteriormente) y el estriado ventral y el córtex cingulado anterior (en la implementación de conductas de decisión propiamente dichas).
El daño en cualquier región de esta red neural limitaría la realización óptima de decisiones, dificultando la adaptación del organismo a su contexto cotidiano e impidiendo un funcionamiento saludable.
Conflicto de interesesLos autores declaran no tener ningún conflicto de intereses.

