




The paramedian lobule of the cerebellum is involved in learning to correctly perform motor skills through practice. Dendritic spines are dynamic structures that regulate excitatory synaptic stimulation. We studied plastic changes occurring in the dendritic spines of Purkinje cells from the paramedian lobule of rats during motor learning.
MethodsAdult male rats were trained over a 6-day period using an acrobatic motor learning paradigm; the density and type of dendritic spines were determined every day during the study period using a modified version of the Golgi method.
ResultsThe learning curve reflected a considerable decrease in the number of errors made by rats as the training period progressed. We observed more dendritic spines on days 2 and 6, particularly more thin spines on days 1, 3, and 6, fewer mushroom spines on day 3, fewer stubby spines on day 1, and more thick spines on days 4 and 6.
ConclusionThe initial stage of motor learning may be associated with fast processing of the underlying synaptic information combined with an apparent “silencing” of memory consolidation processes, based on the regulation of the neuronal excitability.
El lóbulo paramediano del cerebelo está involucrado en el desempeño correcto de las habilidades motoras a través de la práctica. Las espinas dendríticas son estructuras dinámicas que regulan la estimulación sináptica excitadora. En este trabajo se estudiaron los posibles cambios plásticos en espinas de células de Purkinje del lóbulo paramediano cerebelar de ratas, durante el aprendizaje motor.
MétodosSe entrenaron a ratas macho adultas durante un período de seis días, en un paradigma de aprendizaje motor acrobático y se cuantificó tanto la densidad como los tipos de espinas dendríticas en cada uno de los seis días de estudio, mediante una modificación al método de Golgi.
ResultadosLa curva de aprendizaje reflejó una disminución consistente de los errores cometidos en el transcurso de los días de entrenamiento. Así mismo, se observaron más espinas dendríticas en los días 2 y 6 y, en particular, más espinas delgadas en los días 1, 3 y 6, menos espinas en hongo el día 3, menos espinas gordas el día 1 y más espinas anchas los días 4 y 6.
ConclusiónEl período inicial de aprendizaje motor podría estar asociado con el procesamiento rápido de la información sináptica subyacente y con un aparente «silenciamiento» de los procesos de consolidación mnémica, en una base de regulación de la excitabilidad neuronal.
Neuroplasticity underlies brain repair in response to environmental factors that alter neurobiological processes. The neuropathological damage that causes motor dysfunction is commonly followed by plasticity responses that act to restore normal activity at different levels, including at the level of synaptic contacts. However, spontaneous recovery of the lost motor function is infrequent, which suggests that some of the neuroplastic events taking place during the recovery period are not effective. In fact, these events are largely unknown.
The recovery of certain skills requires them to be relearned, a process in which cerebellar activity plays a significant role.1–3 The paramedian lobule of the cerebellar cortex is closely related to motor learning. Purkinje cells (PC) in this region integrate afferent information from both extrinsic and intrinsic neural systems, which is necessary for encoding the patterns underlying motor learning. Plastic changes have been reported in the distal synapses between PCs and granule cell parallel fibres (PF), which underlie motor learning in rats trained to learn new motor skills.4–6 Similarly, increased numbers of dendritic spines have been reported in the distal dendrites of PCs after 26 days of acrobatic motor learning, in which the number of foot faults observed decreased during the first 6 days, then asymptotically stabilised.4,7,8 These findings point to the occurrence of neuroplastic events at the level of distal synapses between PFs and PCs mediated by dendritic spines in the cerebellar paramedian lobule during the critical period of motor learning. According to studies by Doyon and Benali2 and Dayan and Cohen,9 motor skills are learnt after several different stages, whose duration depends on the task. The first stage is called the “fast stage,” in which acquisition of motor skills is especially evident, with efficiency improving considerably as this stage progresses.
Dendritic spines may display several types of plastic changes, including neoformation, reabsorption, and geometric remodelling, especially related to the translation of afferent information.10–14 As PF–PC synapses are involved in modulating motor learning, we studied the cytoarchitecture of distal dendritic spines of PCs in the paramedian lobule of the cerebellum of rats trained according to a motor learning paradigm.
Material and methodsWe used 72 adult male Sprague–Dawley rats (250-300g); animals were kept in standard vivarium conditions with 12-hour light/dark cycles (07:00-19:00h), 45% to 50% humidity, room temperature of 22±2°C, and free access to water and conventional rodent chow. Our experimental protocols were approved by the research ethics committee of the National Institute of Health (Mexico) and were designed in accordance with the Guide for the Care and Use of Laboratory Animals (NIH Publication No. 8023, 1996 revision).
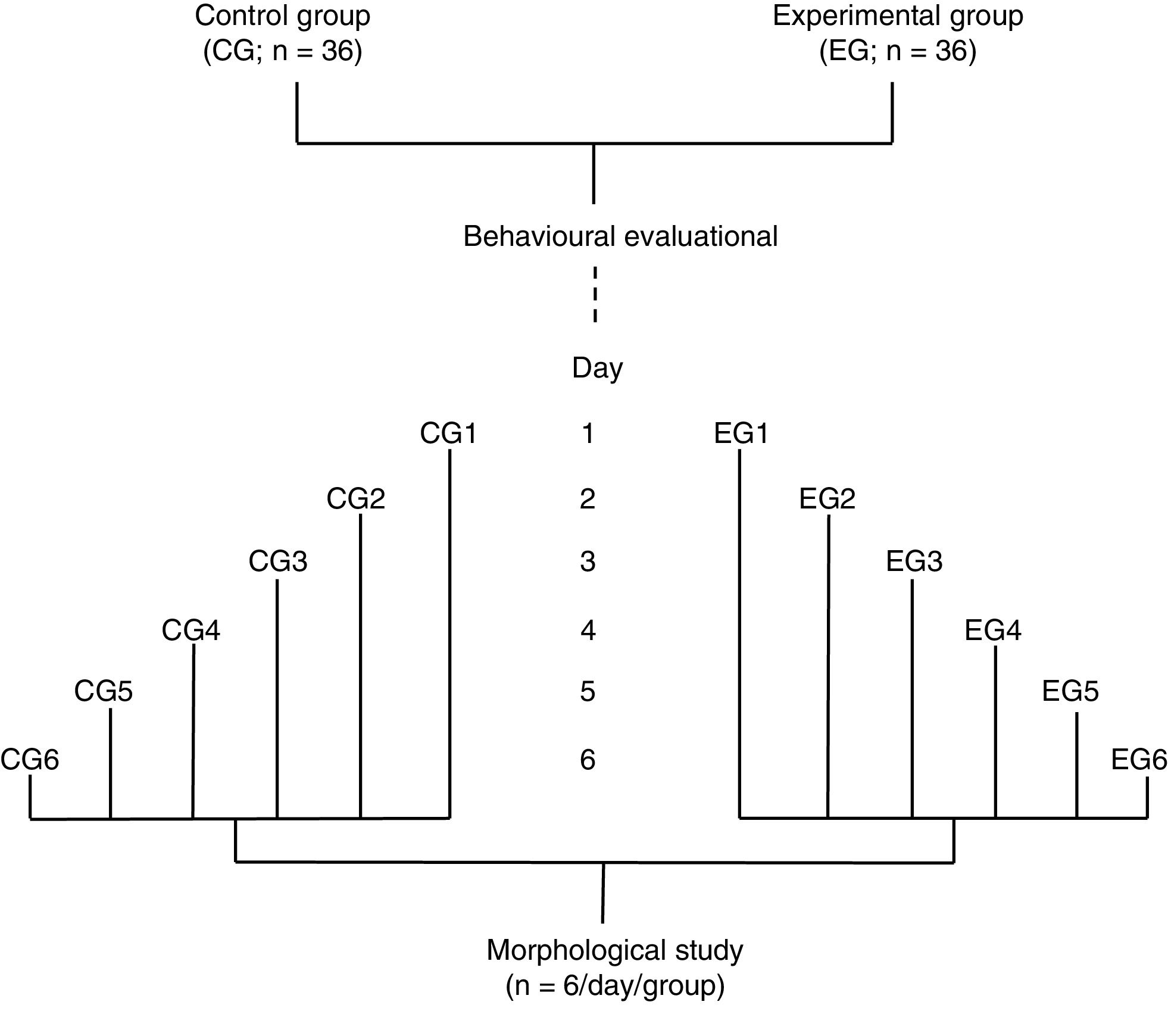
We established 2 groups: an experimental acrobatic group (EG, n=36), which completed an acrobatic motor learning test on 6 consecutive days, and a control group (CG, n=36), which travelled the same distance as the EG but on a flat, obstacle-free surface. The 36 rats in each group were subdivided into 6 subgroups (n=6 per subgroup). The test was performed on consecutive days, from day 1 (EG1/CG1) to day 6 (EG6/CG6), with the number of training days increasing by subgroup (Fig. 1). According to previous studies,6–8 the fast motor learning period2,9 occurs during the first week of training in a spaced time paradigm.15
Behavioural study
Experimental animals were trained to traverse an elevated course (4.25m at 1m above the floor) in 4 trials per day with no time limit, with intervals of 60seconds between trials, between 11:00 and 14:00. The course consisted of a steel chain (50cm length), a horizontal wooden ladder (10cm width×50cm length) covered with straws, a woven wire tube (30cm diameter×80cm length), a wooden staircase covered with abacus beads (10cm width×60cm length), parallel bars (5cm width×50cm length), parallel bars with a crossed truss (15cm width×60cm length), and a balance beam (2cm width×75cm length). A cage was placed at the end of the course for the animal to enter.7 Since rats needed motor coordination and balance to overcome the obstacles along the course, we recorded both the time needed to complete the test and the number of foot faults (errors) in each trial, as a measure of motor learning. The animals in all study groups were housed in groups of 6 per cage, in a room different to that used for training.
Morphological studyImmediately after their final session of behavioural training, the 6 animals from the corresponding experimental and control groups were anaesthetised with intramuscular ketamine (30mg/kg) and intraperitoneal sodium pentobarbital (50mg/kg). They were immediately perfused with 200mL of phosphate-buffered saline (pH 7.4; 0.01M) containing sodium heparin (1000IU/L) as an anticoagulant and procaine hydrochloride (1g/L) as a vasodilator.16 Animals were subsequently perfused with 200mL of a phosphate-buffered 4% formaldehyde fixative solution. Both solutions were perfused at a flow rate of 11.5mL/minutes. Brains were obtained by craniectomy and were kept in 100mL of fresh fixative solution for 48hours. A block of tissue from the left cerebellar hemisphere, containing the paramedian lobule,17 was dissected and impregnated using a modified version of the Golgi technique.18 Using 75μm thick sagittal slices, we performed a “blind” study of 6 clearly visible PCs from each rat. In each of the 6cells studied per rat, we counted dendritic spines in a total section of 50μm from 3 to 4 terminal apical dendritic branchlets distal to the soma (Fig. 2, left panel).
, mushroom (m), stubby (s), and wide (w) spines (arrows), similar to the ones counted in our study. Scale bar: 2μm.")
Left panel: photomicrograph of a Purkinje cell from the cortex of the cerebellar paramedian lobule. Arrows point to the distal dendritic branchlets where spines were counted. Scale bar: 100μm. Right panel: photomicrograph representing thin (t), mushroom (m), stubby (s), and wide (w) spines (arrows), similar to the ones counted in our study. Scale bar: 2μm.
We quantified the numerical and proportional density of thin, mushroom, stubby, and wide spines, according to previously established criteria11,19–23 (Fig. 2, right panel). Spines were counted by direct observation at 2000× using a magnification changer coupled to a light microscope and a 100× Apochromat objective and an image analyser (LAS 4.0).
Statistical analysisMean values for both the time taken to complete the course and the number of errors, as well as the density and morphology of dendritic spines, were calculated for the 6 animals studied each day; comparisons were made between subgroups 1, 2, 3, 4, 5, and 6 using two-way ANOVA (group×day), followed by the t test for independent samples.
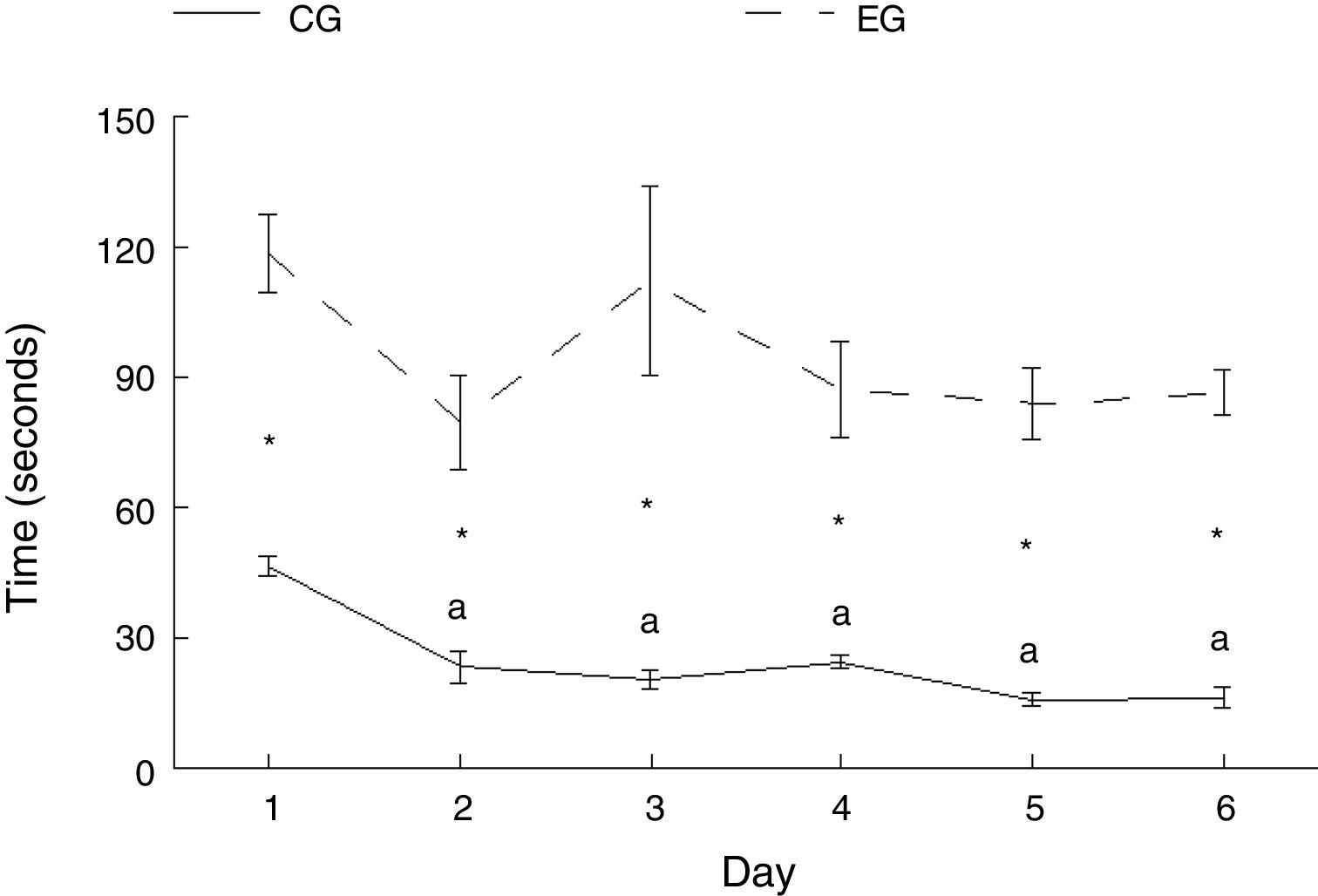
ResultsMotor learningTimeExperimental animals took longer to complete the course than control animals on all training days: day 1, t=−7.847, P<.0001; day 2, t=−4.847, P<.001; day 3: t=−4.137; P<.002; day 4, t=−5.543, P<.0001; day 5, t=−8.801, P<.0001; day 6, t=−11.675, P<.0001. There were no differences in the time taken by experimental animals to complete the course on any of the 6 days of study. Furthermore, the control subgroups showed differences in time to complete the test (F=21.614, P<.0001); the rats in subgroups CG2 (P<.0001), CG3 (P<.0001), CG4 (P<.0001), CG5 (P<.0001), and CG6 (P<.0001) travelled the required distance quicker than those in subgroup CG1. No differences were observed between subgroups CG2, CG3, CG4, CG5, and CG6 (Fig. 3).
; CG: control group; EG: experimental group.")
Graph showing the time taken by experimental and control animals to complete the behavioural task over the 6-day training period. Mean±SEM. Statistical significance was set at P=.05. Asterisks: significant difference between EG and CG; a: significant difference vs day 1 (CG); CG: control group; EG: experimental group.
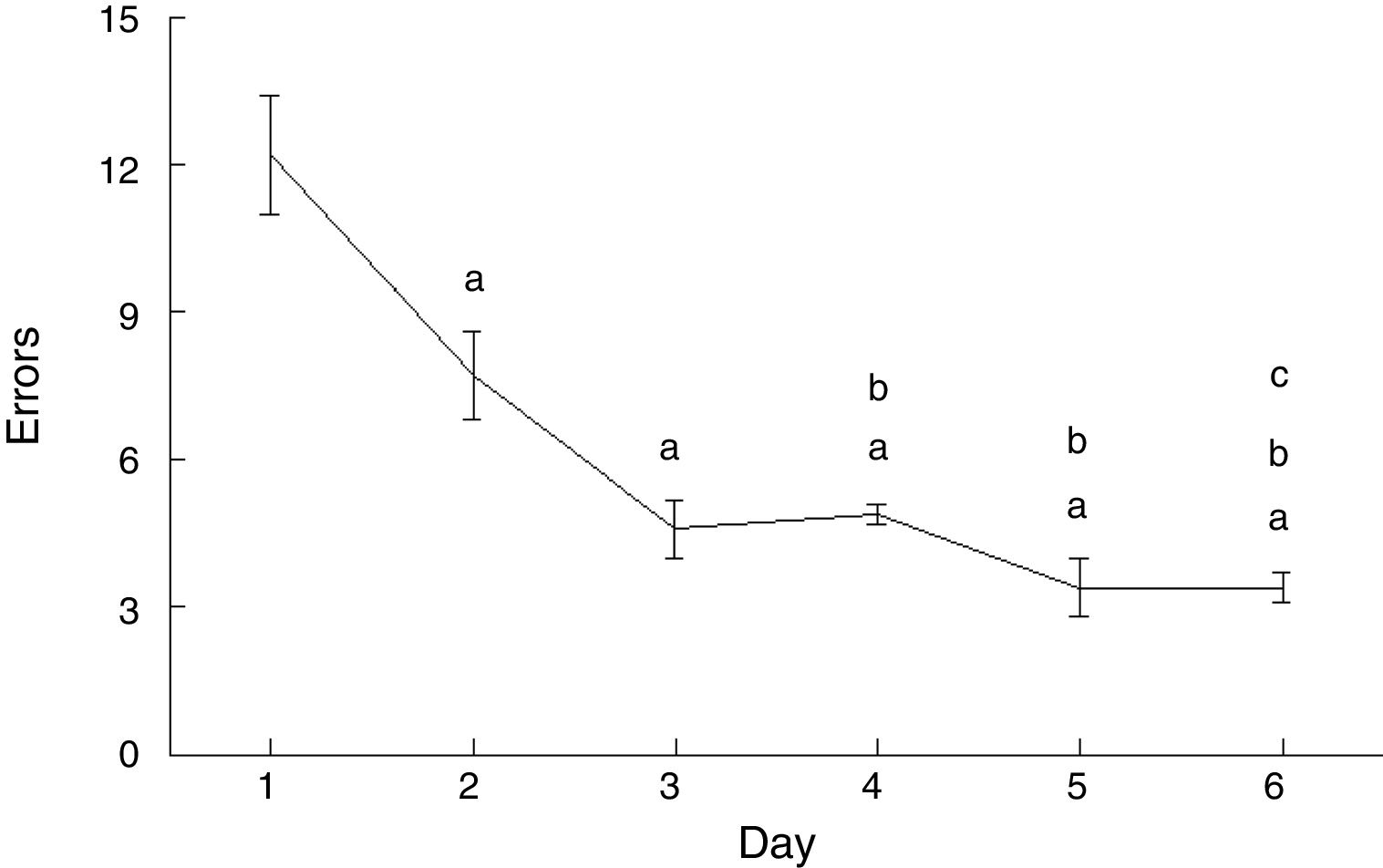
The number of errors made by experimental animals decreased as the days of training advanced (F=20.169, P<.0001). Compared to day 1, rats made fewer errors on days 2 (P<.003), 3 (P<.0001), 4 (P<.0001), 5 (P<.0001), and 6 (P<.0001). Furthermore, EG4 (P<.004), EG5 (P<.004), and EG6 rats (P<.004) made fewer errors than those in the EG2 subgroup, with EG6 rats also making fewer errors than EG4 rats (P<.02) (Fig. 4).
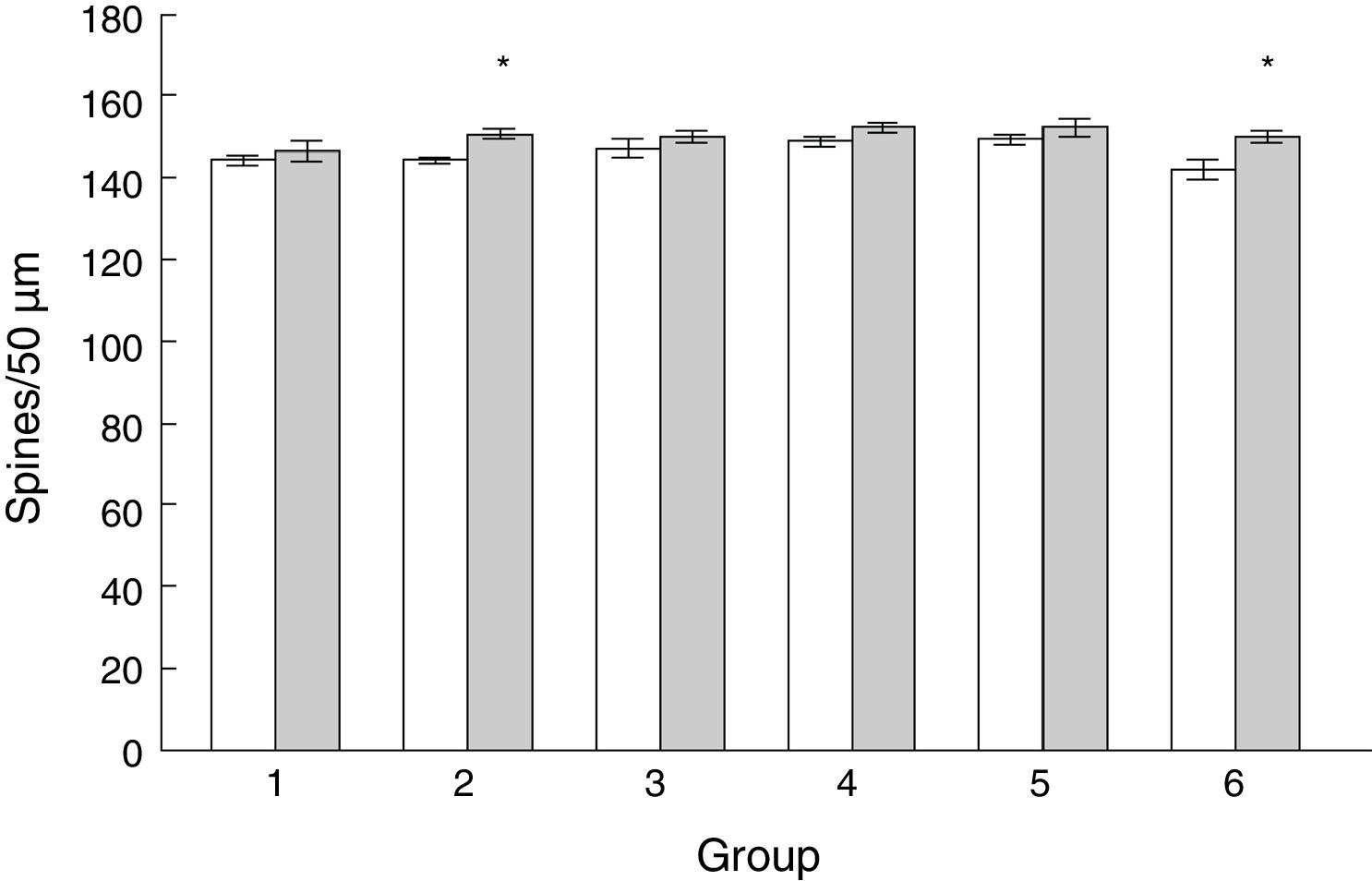
Density of dendritic spines. Statistical significance was established at P<.05; (a) significant difference vs day 1; (b) significant difference vs day 2; (c) significant difference vs day 3.")
Density of dendritic spines on PCs of the paramedian lobules of animals from the EG2 (t=−3.713, P<.004) and EG6 subgroups (t=−2.577, P<.02) was higher than in the CG2 and CG6 subgroups (Fig. 5).
Types of dendritic spines and control animals (white bars). Mean±SEM. Statistical significance was set at P=.05 (asterisks).")
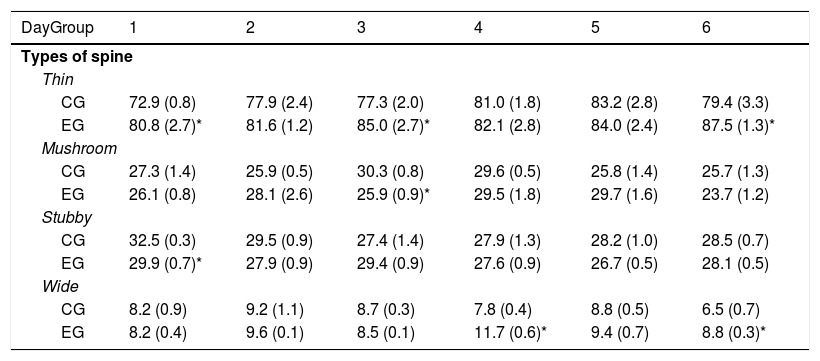
Animals in EG1, EG3, and EG6 subgroups had more thin spines than those in the CG1, CG3, and CG6 subgroups (t=−2.709, P<.02; t=−2.223, P<.05; and t=−2.235, P<.04; respectively). Furthermore, the EG3 subgroup showed fewer mushroom spines than the CG3 subgroup (t=3.452, P<.006). Density of stubby spines was lower in EG1 rats than in CG1 rats (t=3.059, P<.01), whereas animals in the EG4 subgroup showed more wide spines than those in the CG4 subgroup (t=−4.902, P<.001); EG6 subgroup showed the same difference with regard to CG6 (t=2.976, P<.01) (Table 1).
Proportional density of the different types of dendritic spines in Purkinje cells of the cerebellar paramedian lobule, on each day of behavioural training.
| DayGroup | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| Types of spine | ||||||
| Thin | ||||||
| CG | 72.9 (0.8) | 77.9 (2.4) | 77.3 (2.0) | 81.0 (1.8) | 83.2 (2.8) | 79.4 (3.3) |
| EG | 80.8 (2.7)* | 81.6 (1.2) | 85.0 (2.7)* | 82.1 (2.8) | 84.0 (2.4) | 87.5 (1.3)* |
| Mushroom | ||||||
| CG | 27.3 (1.4) | 25.9 (0.5) | 30.3 (0.8) | 29.6 (0.5) | 25.8 (1.4) | 25.7 (1.3) |
| EG | 26.1 (0.8) | 28.1 (2.6) | 25.9 (0.9)* | 29.5 (1.8) | 29.7 (1.6) | 23.7 (1.2) |
| Stubby | ||||||
| CG | 32.5 (0.3) | 29.5 (0.9) | 27.4 (1.4) | 27.9 (1.3) | 28.2 (1.0) | 28.5 (0.7) |
| EG | 29.9 (0.7)* | 27.9 (0.9) | 29.4 (0.9) | 27.6 (0.9) | 26.7 (0.5) | 28.1 (0.5) |
| Wide | ||||||
| CG | 8.2 (0.9) | 9.2 (1.1) | 8.7 (0.3) | 7.8 (0.4) | 8.8 (0.5) | 6.5 (0.7) |
| EG | 8.2 (0.4) | 9.6 (0.1) | 8.5 (0.1) | 11.7 (0.6)* | 9.4 (0.7) | 8.8 (0.3)* |
EG: experimental group; CG: control group.
Mean±SEM.
The paramedian lobule of the cerebellum participates in the integration of information related to motor learning, as has been reported in studies showing increased metabolic activity in this cerebellar region during the first days of acquisition of motor skills.9,24 Therefore, cerebellar activity may be influenced by changes in synaptic information processing between PFs and PCs,4–6 due to the remodelling of dendritic spines mediated by motor learning. In line with this, we observed an increase in the number of dendritic spines in PCs during the fast motor learning period, corresponding to a reduction in the number of errors made as rats completed the acrobatic course.
Although the distance travelled by experimental and control animals was the same, the time required to complete the course with obstacles was longer, which could be explained by the fact that motor requirements were more complex for the experimental group. Although the time taken to complete the task was not significant in comparison with day 1, we did observe a downward trend, which would be partially consistent with previous reports.7,8
As reported in previous studies,7,8 the number of errors made by experimental rats decreased with the days of training. This could be interpreted as a gradual and progressive increase in the precision and control of motor activity and demonstrates the appearance of a learning process.25,26 Synaptic inputs to the apical dendritic region of PCs are directly related to the organisation of the psychoneural events leading to better performance of motor skills1,4–8,27; according to other authors,28 the underlying synaptic stimulation might be related both to the potentiation and to the depression of synapses between parallel fibres and PC dendritic spines. Thus, the gradual increase in efficiency observed over the training period may have involved structural modifications to the synapses in the cerebellar paramedian lobule,15 which may correspond to plastic changes in dendritic spines, as observed in our study.
The higher number of dendritic spines reported at training days 2 and 6 suggests a higher level of presynaptic activity as a result of motor learning. Several studies have associated the increased number of dendritic spines with higher presynaptic activity,29,30 including that related to motor learning.19,28,31 In addition to spine density, plastic changes in the types of dendritic spines may also participate in the neurobiological events underlying the processing of information related to motor learning. PCs display several types of dendritic spines, characterised by their shape; these include the thin, mushroom, stubby, and wide types.19–21 Thin spines are associated with fast, highly efficient synaptic neurotransmission, the induction of long-term potentiation,32 and events related to information acquisition or new learning.13,14 Furthermore, synaptic stimuli in mushroom spines are processed in the cell nucleus, which has been associated with the consolidation of information previously acquired as traces of memory.13,14 When synaptic stimulation of neurons is sufficiently high to prevent synaptic integration,19 the functional activity of stubby and wide spines would participate by regulating the excitability of the postsynaptic neuron,11,12,33 as it lacks a slender neck.34,35
A trend towards greater abundance of thin spines was observed on all training days, with the number being significant at days 1, 3, and 6. As the animals were exposed to a new experience on the first day of training, and because transmission of this new information mediated by synapses with thin spines on PCs involves fast information processing,36 the increase in this type of spines seems to have been functionally significant. The increased density of thin spines may also be causally related to the nature of the behavioural task. The volume of the heads of large spines has been demonstrated to decrease after spaced training and to change to the extent that they transform into thin spines,15 which may be related to long-term potentiation and depression associated with synaptic activity in the contacts between PFs and PCs.28 Therefore, the increased density of thin spines in the long term during training may account for the increasingly accurate performance of the skills needed to resolve the behavioural task, that is, motor learning.
The density of mushroom spines was only different at training day 3, with experimental animals showing lower levels than control rats. As mushroom spines are related to information storage,13,14 this finding is relevant when considered in association with the higher number of thin spines. Together, these events may suggest that synaptic activity related to the information storage underlying motor performance is secondary to the acquisition of this information, as reflected by the predominance of thin spines. If this were the case, the capacity to acquire new information would not be conditioned by consolidation of the information, represented by mushroom spines; this is translated into a “released” capacity to consistently acquire novel information throughout the training period. This would be consistent with previous reports showing an increase in all types of spines (including mushroom spines) 4 weeks after the implementation of a motor learning paradigm similar to the one used in this study, in which the learning curve is not modified after the first week of training, following an initial stage of consistent learning.4,7,8 This suggests that the information required to perform a task efficiently and consistently has been consolidated, presumably with considerable participation of mushroom spines as mediators of the synaptic processes associated with motor learning.
The ratio of stubby spines was lower on training day 1, whereas the ratio of wide spines was higher on days 4 and 6. The smaller number of stubby spines on day 1 may mean that synaptic inputs to PCs may be highly specific when correlated with the proportional increase in thin spines observed on the same day of training. Furthermore, the higher number of wide spines recorded in the last days of training suggests that, despite active learning of motor skills, the increasingly efficient performance of a task would involve functionally redundant synaptic activity. This would be reflected in an increase in the proportion of spines participating in the regulation of afferent synaptic activity, such as wide spines.
This evidence suggests a close relationship between the acquisition of new motor skills and the plasticity of dendritic spines. In this sense, motor learning has been reported to induce “non-linear” plastic changes in dendritic spines during successive stages of training,37 which is clearly consistent with our findings.
Some other synaptic connections between neuronal elements of the cortex of the cerebellar paramedian lobule would be involved in the integration of information related to motor learning38–40; this probably also occurs in other cerebellar lobules.41 Furthermore, possible variations in the cytoarchitecture of PCs from different areas42 of the same paramedian lobule could be considered a source of plastic changes in the morphology of spines, which may affect motor learning. These possibilities require further study.
This study underscores the significance of plastic events at the synaptic level in the initial stages of motor learning, favouring the acquisition of motor skills in normal or neuropsychopathological conditions.
FundingThis study was financed by the Health Research Fund of the Mexican Institute of Health, with registry number FIS/IMSS/PROT/G14/1336.
Conflicts of interestThe authors have no conflicts of interest to declare.
Please cite this article as: González-Tapia D, González-Ramírez MM, Vázquez-Hernández N, González-Burgos I. El aprendizaje motor induce cambios plásticos en las espinas dendríticas de las células de Purkinje del cerebelo de ratas. Neurología. 2020;35:451–457.
Part of this work was presented at the 45th Congress of the Society for Neuroscience.
recomendados

Neurología (English Edition) sigue las recomendaciones para la preparación, presentación y publicación de trabajos académicos en revistas biomédicas