






Se aislaron bacterias rizosféricas y endófitas a partir de rizósfera y tejidos de raíz de árboles de Eucalyptus nitens con el objetivo de evaluar su capacidad de promover el crecimiento en plántulas de la misma especie en condiciones de invernadero. Los aislamientos que incrementaron el crecimiento de las plántulas fueron identificados y caracterizados por su capacidad de producir ácido indolacético (AIA), solubilizar fosfato y expresar la 1-aminociclopropano-1-carboxilato (ACC) desaminasa. Los 105 aislamientos obtenidos fueron morfológicamente diferentes y solo 15 promovieron significativamente el crecimiento de plántulas de E. nitens. Los máximos incrementos observados fueron en el peso seco aéreo (142 %) y de la raíz (135 %); también aumentaron la altura de las plantas (50 %) y el largo de raíces (45 %) de las mismas. Las rizobacterias pertenecieron a los géneros Arthrobacter, Lysinibacillus, Rahnella y Bacillus. Los aislados identificados como A. phenanthrenivorans 21 y B. cereus 113 incrementaron la emergencia de E. nitens a los 12 días en un valor promedio de 3,15 veces con relación al control. R. aquatilis aislado 78 presentó la mayor producción de AIA (97,5±2,87μg/ml) en presencia de triptófano y el mayor índice de solubilización de fósforo (2,4). B. amyloliquefaciens aislado 60 fue positivo para la actividad ACC desaminasa. Los resultados obtenidos indican el potencial de las rizobacterias estudiadas como promotoras de emergencia y crecimiento de plántulas de E. nitens y su posible uso como inoculantes, ya que presentan más de un mecanismo de acción asociado a la promoción del crecimiento.
Rhizospheric and endophytic bacteria were isolated from the rizosphere and root tissue of Eucalyptus nitens. The objective of this work was to evaluate their capacity to promote growth in seedlings of the same species under greenhouse conditions. The isolates that improved seedling growth were identified and characterized by their capacity to produce indoleacetic acid (IAA), solubilize phosphates and increase 1-aminocyclopropane-1-carboxylate (ACC) deaminase activity. One hundred and five morphologically different strains were isolated, 15 of which promoted E. nitens seedling growth, significantly increasing the height (50%), root length (45%) as well as the aerial and root dry weight (142% and 135% respectively) of the plants. Bacteria belonged to the genus Arthrobacter, Lysinibacillus, Rahnella and Bacillus. Isolates A. phenanthrenivorans 21 and B. cereus 113 improved 3.15 times the emergence of E. nitens after 12 days, compared to control samples. Among isolated R. aquatilis, 78 showed the highest production of IAA (97.5±2.87μg/ml) in the presence of tryptophan and the highest solubilizer index (2.4) for phosphorus, while B. amyloliquefaciens 60 isolate was positive for ACC deaminase activity. Our results reveal the potential of the studied rhizobacteria as promoters of emergence and seedling growth of E. nitens, and their possible use as PGPR inoculants, since they have more than one mechanism associated with plant growth promotion.
La industria forestal en Chile se posiciona como un pilar fundamental en la economía, con más de dos millones de hectáreas de cobertura forestal compuestas principalmente por Pinus radiata D. Don y Eucalyptus spp.19. Eucalyptus nitens (Deane et Maiden) es una especie con una amplia extensión plantada en el sur de Chile31, de unas 133 411 hectáreas8, debido a que posee una madera con importantes ventajas para ser utilizada como materia prima pulpable55, sumadas a su rápido crecimiento y resistencia a bajas temperaturas11.
El éxito de las plantaciones depende en gran parte de la capacidad de las plantas provenientes de los viveros para tolerar el estrés producto del trasplante, ya que ese nuevo ambiente podría ser percibido como adverso y afectar tanto el crecimiento inicial como la supervivencia de las plantas16,17. La calidad de las plántulas producidas en vivero es considerada clave para determinar el éxito de la futura plantación74, y en términos generales, se considera que estas deben tener su parte aérea y raíces lo suficientemente desarrolladas como para aumentar la probabilidad de establecerse66.
Numerosas especies de bacterias, en su mayor parte asociadas con la rizósfera, han demostrado capacidad para incrementar el crecimiento de las plantas; a estas se las denomina rizobacterias promotoras de crecimiento vegetal [en inglés, plant growth promoting rhizobacteria46 (PGPR)]. En diversos cultivos, la aplicación de PGPR en estadios tempranos de desarrollo ha incrementado la producción de la biomasa por un mayor crecimiento de raíces y tallos15,33,47. En viveros de especies forestales, las PGPR han permitido incrementar la supervivencia de las plantas después del trasplante, lo que se ha asociado a un mejor desarrollo de raíces y, en consecuencia, a un aumento de la captación de nutrientes75.
Las PGPR pueden inducir el crecimiento vegetal directamente o indirectamente. La influencia directa incluye la producción de fitohormonas, como por ejemplo, de ácido indolacético70 (AIA), del grupo de las auxinas; también de ácido giberélico (GA3), citoquininas24 y ácido abscísico21,26 (ABA); o bien la capacidad de producir la enzima 1-aminociclopropano 1-carboxilato (ACC) desaminasa, que reduce el nivel de etileno en las raíces34,54,71. Además, los mecanismos directos incluyen la liberación de fosfatos y micronutrientes y la fijación biológica de nitrógeno26. Los efectos indirectos se deben a la modificación del ambiente rizosférico y su ecología51, actuando como agentes de biocontrol de fitopatógenos mediante la liberación de sustancias como sideróforos, β-1, 3-glucanasas, quitinasas, antibióticos, pigmentos fluorescentes y cianidas10,35,67.
Géneros bacterianos como Bacillus y Pseudomonas han demostrado su acción promotora del crecimiento en especies forestales13,14,62. Puente et al.74 demostraron el efecto de la inoculación con Azospirillum brasilense sobre el crecimiento y la germinación de E. globulus. También se ha demostrado una estimulación en el enraizamiento de estacas de E. grandis, E. globulus y E. calmaldulensis25,42,56,87 y un efecto de biocontrol de la marchitez bacteriana en E. urophylla76,77. Sin embargo, a la fecha no existen investigaciones sobre la promoción del crecimiento en E. nitens.
Debido a la creciente expansión de las plantaciones de E. nitens en Chile, las rizobacterias representan una alternativa para optimizar el establecimiento y el crecimiento de esta especie. Por lo tanto, el objetivo de este trabajo fue caracterizar bacterias rizosféricas y endófitas promotoras del crecimiento de E. nitens y determinar los potenciales mecanismos de acción asociados, como la producción de AIA, la solubilización de fosfato y la expresión de la actividad 1-aminociclopropano-1- carboxilato (ACC) desaminasa in vitro.
Materiales y métodosSe colectaron plantas de E. nitens de un año de establecimiento en los predios forestales Choroito y Santa Adriana, ubicados en los 37° 32’ 07, 08” de latitud sur y 72° 44’ 46, 38”de longitud oeste, y también del predio Lo Álvarez, situado en los 37° 25’ 57, 93” latitud sur y 71° 54’ 52,78” longitud oeste; los tres predios se encuentran en la región del Bío Bío, República de Chile, y son propiedad de Forestal Mininco S.A. Las plantas fueron extraídas completas y transportadas en frío al laboratorio de Biopelículas y Microbiología Ambiental del Centro de Biotecnología de la Universidad de Concepción.
Para el aislamiento de bacterias rizosféricas se seleccionaron las raíces laterales jóvenes18, las cuales fueron seccionadas y lavadas con agua destilada esterilizada (ADE), posteriormente fueron suspendidas en solución salina tamponada [phosphate buffered saline (PBS)] y sonicadas durante 15min (40kHz) (Branson 2510 MT, Branson Ultrasonic Corporation, México), y la suspensión rizosférica así obtenida fue filtrada mediante gasa estéril. Una fracción (1ml) del filtrado fue diluida sucesivamente al décimo hasta lograr una dilución 10−5 de la original, y se utilizó una alícuota del filtrado (100μl) para sembrar cada placa87. Se emplearon diferentes medios de cultivo para tratar de aislar a algunos géneros de PGPR frecuentemente comunicados7. Para el aislamiento de Pseudomonas se utilizó King B45 (KB) al 10 %, para el de Streptomyces el medio elegido fue agar almidón caseína49 (AC) suplementado con extracto de suelo38 y cicloheximida (50μg/ml) (Sigma Aldrich, St. Louis, EE.UU.). Para el aislamiento de Bacillus, alícuotas (1ml) de las muestras se sometieron a un choque térmico a 80°C durante 20min y luego se sembraron en el medio de esporulación CCY85. Para el aislamiento de las bacterias endófitas se procedió según el protocolo de Magallan y Dion57, empleando los mismos medios de cultivo utilizados para el aislamiento de bacterias rizosféricas. En todos los casos, la incubación fue a 28°C durante 3 a 5 días. Adicionalmente, se realizaron recuentos de las bacterias endófitas y rizosféricas —unidades formadoras de colonia (UFC)— obtenidas a partir de los tejidos de las raíces y de suelo rizosférico, respectivamente. Luego se procedió a aislar las colonias con morfotipos diferentes (forma, borde, tamaño, color y apariencia), las cuales fueron traspasadas al mismo medio en el que fueron obtenidas. Las colonias puras fueron almacenadas a −80°C en glicerol (Riedel-de Haën, Alemania) al 20 %.
Para la producción del inóculo bacteriano utilizado en los ensayos, los aislados fueron crecidos en caldo tripticasa de soja [en inglés, tryptic soy broth (TSB)] (Becton, Dickinson and Company, Le Pont de Claix, Francia) al 50 % e incubados a 28°C y 120 r.p.m. durante 48h, la concentración se ajustó a 1×107UFC/ml.
Para el ensayo de promoción de crecimiento fueron utilizadas semillas de E. nitens, proporcionadas por Forestal Mininco S.A. Las semillas se trataron inicialmente de acuerdo con el procedimiento usual seguido en condiciones productivas, este consistió en un primer remojo en una solución de ácido giberélico (AG3) (200mg/ml) (Sigma Aldrich, St Louis, EE.UU.) durante una noche y una posterior desinfección superficial en etanol (70 % v/v) por 10s e hipoclorito de sodio (5 % v/v) durante 10min, seguida de lavados sucesivos en ADE. Por último, las semillas fueron tratadas mediante inmersión en el inóculo bacteriano, con la adición de carboximetilcelulosa (CMC) (Protex, China) al 0,6 % p/v en agitación (120 r.p.m.) durante 30min, luego se dejaron secar a temperatura ambiente.
Se utilizó un sustrato natural (corteza de pino compostada, no esterilizada) dispuesto en tubetes plásticos de 70ml de capacidad, que también fue tratado con cada suspensión bacteriana (relación 1:10 v/v), y 48h después se procedió a la siembra. El sustrato presentó las siguientes características fisicoquímicas: materia orgánica 46 %; nitratos (N-NO3), 13,10mg/kg; amonio (N-NH4), 9,10mg/kg; N disponible, 923,10mg/kg; N total, 0,54 %; fósforo disponible, 0,05 %; potasio, 0,14 %; calcio, 0,53 %; magnesio, 0,14 %; sodio, 0,04 %; conductividad eléctrica, 0,2 dS/m; relación C/N, 49,5; y pH en agua (1:2), 5,65. El ensayo fue realizado en el invernadero de la Facultad de Ciencias Forestales de la Universidad de Concepción durante un periodo de tres meses. A las cuatro semanas después de la siembra se realizó una segunda inoculación con 5ml de suspensión bacteriana ajustada a 1×107UFC/ml, direccionada a la base de las plántulas emergidas.
El diseño utilizado para el ensayo fue completamente al azar, con 21 plantas por cada tratamiento. Fueron incluidos 116 tratamientos, con 105 aislados bacterianos obtenidos de ejemplares de E. nitens, 10 aislados con antecedentes de haber promovido el enraizamiento de estacas de E. globulus obtenidos previamente por Díaz et al.25 y un tratamiento control, que consistió en la aplicación de caldo de cultivo TSB al 50 % sin bacterias. Se evaluó la emergencia de las plántulas a los 12, 14, 16, 18, 20, 24 y 28 días después de la siembra, y al cabo del tercer mes se colectaron las plantas, para evaluar altura de la parte aérea, largo de raíces, peso seco de la planta y de raíces totales (secadas a 70°C durante 48h). El análisis de la emergencia consistió en la estimación y comparación de las probabilidades de emergencia de las plántulas entre tratamientos. Se consideró que el número de plantas emergidas siguió una distribución binomial con probabilidad de emergencia P. El análisis inferencial se efectuó mediante regresión logística y las comparaciones de probabilidades de emergencia versus el testigo se ajustaron según el método de Agresti1,2. Para el análisis de las variables de promoción del crecimiento se realizó un análisis no paramétrico de Kruskal Walllis (p<0,05), utilizándose el software R.
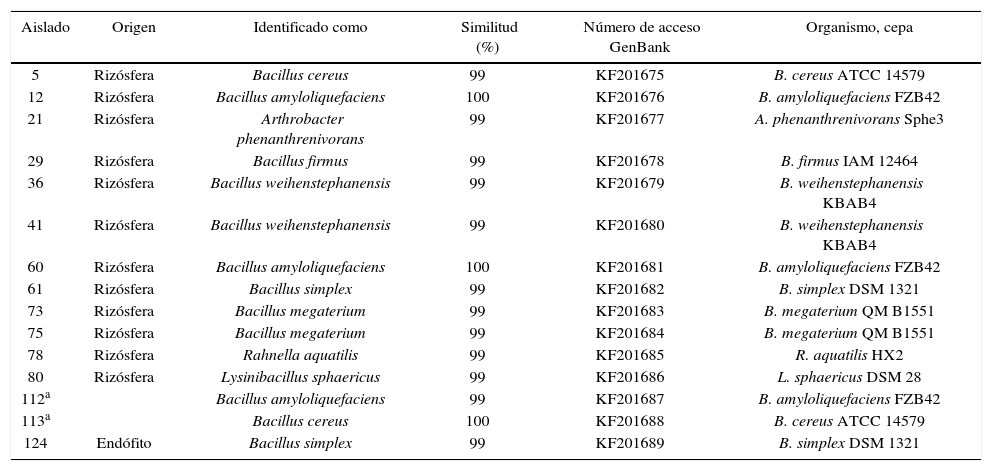
Los aislados con actividad PGPR fueron identificados con métodos genómicos, el ADN bacteriano fue aislado mediante el kit ZR fungal/bacterial DNA MicroPrep™ (Zymo Research, Irvine CA, EE.UU.) y utilizado como templado para la amplificación por PCR del 16S ADNr. Para la amplificación por PCR se emplearon los cebadores 27-F [5’- GAGTTTGATCCTGGCTCAG-3´]50 y 1542-R [5’- AGAAAGGAGGTGATCCAGCC-3’]68, que amplifican para un producto de PCR de 1520 pb. La mezcla de reacción consistió en 1X buffer Flexi DNA polymerase (Promega, Madison, EE.UU.), 1,5mM MgCl2, 0,5mM de cada dNTP, 0,5mM de cada partidor y 0,025mM de Go Taq polymerase (Promega, Madison, EE.UU.). Las condiciones de amplificación fueron las siguientes: un ciclo de preincubación a 95°C por 2min, 30 ciclos de amplificación en tres segmentos, con desnaturalización a 95°C por 30s, hibridación a 54°C por 30s y extensión a 72°C por 1,5min, y la extensión final a 72°C durante 2min. Los productos de PCR fueron secuenciados en Macrogen Inc. (Corea del Sur). Las secuencias del gen 16S ADNr fueron analizadas con el programa de búsqueda Basic Local Alignment Search Tool (BLAST), para encontrar identidad entre las secuencias (≥ 99 %), y posteriormente fueron depositadas en la base de datos de secuencias nucleotídicas GenBank (tabla 1).
Identificación de los aislados promotores de crecimiento en Eucalyptus nitens basada en la secuenciación del gen 16S del ADNr
| Aislado | Origen | Identificado como | Similitud (%) | Número de acceso GenBank | Organismo, cepa |
|---|---|---|---|---|---|
| 5 | Rizósfera | Bacillus cereus | 99 | KF201675 | B. cereus ATCC 14579 |
| 12 | Rizósfera | Bacillus amyloliquefaciens | 100 | KF201676 | B. amyloliquefaciens FZB42 |
| 21 | Rizósfera | Arthrobacter phenanthrenivorans | 99 | KF201677 | A. phenanthrenivorans Sphe3 |
| 29 | Rizósfera | Bacillus firmus | 99 | KF201678 | B. firmus IAM 12464 |
| 36 | Rizósfera | Bacillus weihenstephanensis | 99 | KF201679 | B. weihenstephanensis KBAB4 |
| 41 | Rizósfera | Bacillus weihenstephanensis | 99 | KF201680 | B. weihenstephanensis KBAB4 |
| 60 | Rizósfera | Bacillus amyloliquefaciens | 100 | KF201681 | B. amyloliquefaciens FZB42 |
| 61 | Rizósfera | Bacillus simplex | 99 | KF201682 | B. simplex DSM 1321 |
| 73 | Rizósfera | Bacillus megaterium | 99 | KF201683 | B. megaterium QM B1551 |
| 75 | Rizósfera | Bacillus megaterium | 99 | KF201684 | B. megaterium QM B1551 |
| 78 | Rizósfera | Rahnella aquatilis | 99 | KF201685 | R. aquatilis HX2 |
| 80 | Rizósfera | Lysinibacillus sphaericus | 99 | KF201686 | L. sphaericus DSM 28 |
| 112a | Bacillus amyloliquefaciens | 99 | KF201687 | B. amyloliquefaciens FZB42 | |
| 113a | Bacillus cereus | 100 | KF201688 | B. cereus ATCC 14579 | |
| 124 | Endófito | Bacillus simplex | 99 | KF201689 | B. simplex DSM 1321 |
La relación filogenética se infirió usando el método Neighbor-Joining80, validado utilizando el árbol consenso boostrap inferido de 500 réplicas29. Los análisis filogenéticos fueron llevados a cabo mediante el paquete de software MEGA 486.
Con aquellos aislados que presentaron actividad PGPR se realizaron ensayos in vitro para determinar la ocurrencia de los mecanismos asociados a la promoción del crecimiento vegetal. Para la detección de AIA, las bacterias fueron multiplicadas en caldo TSB (50 %) e incubadas a 28°C y 120 r.p.m. durante 24h, posteriormente se ajustó la concentración a 1 en la escala Mc Farland (3×108cel/ml) y una alícuota de 2 % v/v de cada cultivo fue inoculado en caldo nutriente con L-triptófano (5mg/ml) (MP Biomedicals LLC, Illkirch, Francia) y sin este, e incubados en las mismas condiciones durante el lapso de cinco días. Luego las muestras fueron centrifugadas a 20 000 r.p.m. a 4°C durante 20min y se reveló el sobrenadante con la solución de Salkowski [4,5g de FeCl3 (AppliChem, Darmstadt, Alemania) en H2SO4 10,8 M] en una relación 1:2 muestra-reactivo36 durante 30min en la oscuridad. Para la cuantificación se realizó una curva estándar con las concentraciones: 1, 5, 10, 15, 20, 25, 30, 35 y 40μg/ml de AIA (Sigma Aldrich, St Louis, EE.UU.). Las lecturas se efectuaron en un lector de placas (Infinite® 200 PRO, Tecan, Männedorf, Suiza) a 530nm de longitud de onda y el control se calibró con el caldo de cultivo suplementado con triptófano en ausencia de inóculo. Los ensayos fueron realizados por triplicado bajo un diseño completamente al azar, se empleó el análisis estadístico de varianza (ANOVA) y las medias fueron comparadas mediante el test LSD (p < 0,05). Los datos fueron analizados con el paquete estadístico Statistical Analysis Software (SAS), versión 8.
En presencia de triptófano la producción de AIA de los aislados fue confirmada mediante el análisis cuantitativo por cromatografía líquida de alta eficacia [en inglés, high-performance liquid chromatography (HPLC)] (SPD-M10Avp, Shimadzu, EE.UU.). Para la separación cromatográfica, las muestras fueron filtradas en discos de 0,22 μm de porosidad e inyectadas en forma isocrática en una columna de fase reversa C18. Como fase móvil se utilizó una mezcla de metanol-ácido acético 0,5 % en una proporción 60:40 (v/v), a una velocidad de flujo de 0,7ml/min y a temperatura ambiente. La detección se realizó por fluorescencia utilizando 280nm de excitación y 340nm de emisión9.
Para detectar la capacidad de las bacterias PGPR de solubilizar fósforo (P), estas fueron cultivadas en el medio NBRIP63, con Ca3(PO4)2 (Sigma Aldrich, St Louis, EE.UU.) como fuente de P insoluble, la incubación se efectuó a 28°C durante 5 días. Aquellas con crecimiento en el medio y con la formación de un halo alrededor de las colonias fueron consideradas solubilizadoras y evaluadas mediante el índice de solubilización (IS) = diámetro del halo (mm)/diámetro de la colonia (mm). La capacidad de solubilización se categorizó en tres niveles: bajo (IS < 2), intermedio (IS entre 2 y 4) o alto60 (IS > 4). Las evaluaciones fueron realizadas a los 3 y 5 días, los aislados fueron “solubilizadores tempranos” cuando el halo se formó hasta el tercer día y “solubilizadores tardíos” cuando ocurrió después del tercer día60. Los ensayos fueron realizados por triplicado.
La actividad ACC desaminasa fue determinada cualitativamente de acuerdo con el método de Penrose y Glick72. Los aislados crecieron en el medio peptona proteasa, caseína hidrolizada, anhidro y glicerol (PAF)72 a 28°C y 120 r.p.m. durante 24h, luego se transfirió una alícuota del cultivo (2 % v/v) al mínimo de sales Dworkin y Foster (DF)27 suplementado con 1-aminociclopropano-1-ácido carboxílico (ACC) (MP Biomedicals LLC, Illkirch, Francia) como única fuente de N, y se continuó incubando en las condiciones mencionadas antes. Una alícuota de este cultivo fue transferida a un nuevo caldo DF que se incubó durante 48h, la presencia de turbidez se tomó como un resultado positivo.
Resultados y discusiónA partir de las muestras de raíces de E. nitens se determinaron las poblaciones bacterianas cultivables, que fueron de 5,2×105 UFC/g para rizósfera y 3,1×104 UFC/g para endófitos. Se obtuvieron un total de 105 aislados con diferentes morfotipos, de los cuales 76 fueron de rizósfera y 29 endófitos. Entre estos aislados bacterianos, 15 incrementaron el crecimiento de las plántulas de E. nitens para las variables altura y biomasa de la parte aérea y largo y biomasa de raíces con relación al control (p<0,05), aunque estos aislados no presentaron diferencias entre sí.
Los aislados promotores de crecimiento correspondieron a 12 rizosféricos, uno endófito y dos aislados por Díaz et al.25, los que generaron un incremento de 25 % a 50 % de la parte aérea de las plantas y de 55 % a 142 % en el peso seco de la mismas (fig. 1A). El largo de las raíces se incrementó en un 19 % a 45 %, y la biomasa en un 58 % a 135 % (fig. 1B). Las PGPR se identificaron como pertenecientes a los géneros Bacillus, Lysinibacillus, Arthrobacter y Rahnella (tabla 1), por tanto, vinculadas con tres familias bacterianas: Bacillaceae, Micrococcaceae y Enterobacteriaceae, con un predominio de la primera, que comprendió el 87 % de los aislamientos. La figura 2 exhibe las relaciones filogenéticas entre las especies aisladas. Numerosos estudios realizados con el género Bacillus han demostrado su potencial en la promoción de crecimiento y se han comunicado efectos promotores por acción del biocontrol40, por la solubilización de fosfatos y por la producción de fitohormonas, como AIA39,83. Las especies de Bacillus conforman un grupo predominante en diferentes tipos de suelos30 y se han encontrado estrechamente relacionadas con la promoción del crecimiento en una amplia variedad de plantas22,48 y con el control de fitopatógenos40, a lo que se suma de manera destacada la capacidad de producir endosporas, que les permite sobrevivir por largos períodos en suelo y en almacenamiento64. Las respuestas a la inoculación con PGPR de especies forestales ha sido variable74. Los incrementos de biomasa encontrados en este trabajo fueron en promedio de 89 % y 98 % para la parte aérea y las raíces, respectivamente. Puente et al.74 registraron incrementos de 86 % y 54 % en las variables mencionadas, respectivamente, al tratar plántulas de E. globulus con A. brasilense, mientras que Mohamad et al.61 obtuvieron un incremento de la biomasa del 44 % en plántulas de E. calmadulensis inoculadas con Azotobacter chroococcum y Bacillus megaterium. Entre los mayores incrementos de biomasa documentados, cabe mencionar los resultados de Teixeira et al.87, quienes hallaron incrementos de hasta 223 % en la biomasa de raíces al tratar estacas de Eucalyptus spp. con una cepa de Bacillus subtilis, y los de Díaz et al.25, que observaron un incremento de 168 % en la biomasa de raíces de estacas de E. globulus al ser tratadas con Bacillus mycoides (C2T18).
: longitud y peso seco de la parte aérea; (B): longitud y peso seco de las raíces, después de tres meses de crecimiento en invernadero. Las barras indican el error estándar.")
Crecimiento de plántulas de Eucalyptus nitens inoculadas con 15 diferentes aislados promotores de crecimiento. (A): longitud y peso seco de la parte aérea; (B): longitud y peso seco de las raíces, después de tres meses de crecimiento en invernadero. Las barras indican el error estándar.
. Los números de acceso del GenBank se hallan entre paréntesis.")
Árbol filogenético basado en la secuenciación del gen 16S del ADNr, que muestra la relación evolutiva entre los 15 aislados promotores de crecimiento inferida mediante el método Neighbor-Joining. Las distancias evolutivas se computaron con el empleo del método de máxima verosimilitud.
Los números en las ramas indican el porcentaje de réplicas de árboles en los cuales los taxones asociados se agrupan para el test de bootstrap (500 réplicas). Los números de acceso del GenBank se hallan entre paréntesis.
Con relación a la emergencia, A. phenanthrenivorans aislado 21 y B. cereus aislado 113 promovieron la mayor emergencia temprana en E. nitens (p<0,05), con probabilidades de emergencia de 0,8 y 0,78 a los 12 días después de la siembra comparado con 0,25 del testigo, lo que implica un incremento promedio de la emergencia para este tiempo de 3,15 veces. Sin embargo, a partir del día 14 no se observaron diferencias entre los tratamientos y el control. El incremento de la emergencia por acción de las PGPR ha sido comunicado con anterioridad23,89. Se ha descrito la promoción del crecimiento y el incremento en el porcentaje de germinación por cepas de Arthrobacter sp. y B. cereus41,79,92. Algunas PGPR han estimulado la emergencia de las semillas hasta el doble del testigo no tratado32, esto se ha asociado a un incremento en la síntesis de hormonas, como giberelinas y auxinas32,41.
En el ensayo de selección de PGPR, el sustrato utilizado fue natural (no esterilizado), condición que implica que las bacterias aplicadas debieron competir con la microbiota natural habitante del compost5. Se ha comunicado que la esterilización del sustrato puede tener resultados variables en la promoción del crecimiento en las plantas13,32. En la condición de sustrato de este ensayo, las bacterias inoculadas pudieron ejercer diferentes efectos, ya sea el biocontrol sobre bacterias patógenas7 o sobre aquellas bacterias que producen compuestos inhibidores del crecimiento de las plantas91.
Todos los aislados con acción PGPR en E. nitens produjeron AIA, independientemente del suministro de triptófano, sin embargo, la producción de AIA bacteriano en presencia de triptófano fue mayor (tabla 2). El triptófano es considerado un precursor del AIA, consecuentemente, su adición al cultivo bacteriano incrementó la biosíntesis del AIA4,20,88. Las PGPR R. aquatilis aislado 78 y B. megaterium aislado 75 presentaron las mayores concentraciones de AIA. El aislado 78 exhibió la mayor producción de AIA a las 24h, mientras que la producción de B. megaterium aislado 75 alcanzó su valor máximo a las 120h. Otros autores han comunicado la capacidad de producción de AIA de especies de Rahnella y Bacillus, tanto en presencia como en ausencia de triptófano28,43,52.
Producción de AIA bacteriano en presencia y en ausencia de triptófano, solubilización de fósforo y actividad ACC desaminasa por las bacterias promotoras de crecimiento en Eucalyptus nitens
| Aislados | Producción de AIA (μg/ml)a | Solubilización de P (IS)b | Actividad ACC desaminasa | ||
|---|---|---|---|---|---|
| Sin triptófano | Con triptófano | 3 días | 5 días | ||
| B. cereus aislado 5 | 2,27±0,1ab | 5,56±1,0d | - | - | - |
| B. amyloliquefaciens aislado 12 | 1,63±0,2def | 4,99±0,7d | 1,19±0,1 | 1,19±0,4 | - |
| A. phenanthrenivorans aislado 21 | 2,54±0,6a | 6,39±2,8d | 1,20±0,1 | 1,02±0,2 | - |
| B. firmus aislado 29 | 2,07±0,1bc | 5,13±0,7d | - | - | - |
| B. weihenstephanensis aislado 36 | 1,08±0,2g | 5,38±0,2d | - | - | - |
| B. weihenstephanensis aislado 41 | 1,72±0,5de | 6,35±0,3d | - | - | - |
| B. amyloliquefaciens aislado 60 | 1,85±0,1bcd | 6,06±0,3d | 1,07±0,1 | 1,1±0,1 | + |
| B. simplex aislado 61 | 1,61±0,2def | 4,60±0,1d | - | - | - |
| B. megaterium aislado 73 | 1,55±0,3def | 11,18±2,2c | - | - | - |
| B. megaterium aislado 75 | 1,28±0,1fg | 17,04±5,8b | 1,10±0,4 | 1,46±0,6 | - |
| R. aquatilis aislado 78 | 1,80±0,3cd | 28,09±0,6a | 2,00±0,2 | 2,44±0,3 | - |
| L. sphaericus aislado 80 | 1,50±0,1efg | 12,07±2,3c | - | - | - |
| B. amyloliquefaciens aislado 112 | 1,36±0,1efg | 6,42±0,3d | 1,17±0,1 | 1,12±0,1 | - |
| B. cereus aislado 113 | 2,54±0,3a | 7,28±0,3d | - | - | - |
| B. simplex aislado 124 | 1,55±0,2def | 6,03±0,7d | - | - | - |
La cantidad de AIA bacteriano puede ser un factor importante en la promoción del crecimiento. En presencia de triptófano, la producción de la fitohormona en las cepas más activas se encuentra entre 80 a 100μg/ml88, rango dentro del cual se ubicó la producción en R. aquatilis aislado 78. Sin embargo, existe controversia respecto del impacto de las auxinas exógenas sobre el desarrollo de la planta, y se han documentado efectos positivos y también negativos3,73,84. Las consecuencias para la planta son usualmente una función de dos factores: (1) la cantidad de AIA producida que se vuelve disponible para la planta; (2) la sensibilidad de los tejidos de la planta a cambios en la concentración de AIA53.
Con relación a la capacidad de las bacterias de solubilizar el P, el 40 % de los aislamientos con capacidad promotora de crecimiento fueron capaces de manifestar in vitro dicha propiedad (tabla 2). Los IS de los aislamientos considerados “solubilizadores tempranos” fueron bajos a los tres días de incubación, en cambio, R. aquatilis aislado 78 dio un IS máximo a los cinco días del ensayo (tabla 2). Si bien existe una diversidad de especies bacterianas que poseen capacidad de solubilizar componentes fosfatados inorgánicos insolubles, como fosfato tricálcico, fosfato dicálcico, hidroxiapatitas y roca fosfórica37, los principales solubilizadores del P citados se hallan entre las especies de Pseudomonas, Bacillus, Rhizobium, Burkholderia, Achromobacter, Agrobacterium, Micrococcus, Aerobacter, Flavobacterium y Erwinia69. Se han descrito bacterias promotoras del crecimiento vegetal del género Eucalyptus por su capacidad de solubilizar P61. Entre ellas, el género Rahnella ha sido asociado a la solubilización de fosfatos en diversas investigaciones44,65,90. Becerra et al.6 aislaron bacterias solubilizadoras de P de suelos cultivados con uchuva (Physalis peruviana) y entre los mejores aislados identificaron a R. aquatilis, con IS intermedio de 3 a las 48h de incubación. Otros autores han reportado IS altos en especies de los géneros Klebsiella y Enterobacter12,59. Sin embargo, el impacto de la solubilización y nutrición con P en el crecimiento de plantas in vivo e in vitro ha dado resultados variables22. Maliha et al.58 atribuyen a la capacidad buffer del medio el éxito de la solubilización de fosfatos por parte de los microorganismos, ya que se ha demostrado que la solubilización de los complejos Ca-P está mediada principalmente por la disminución del pH.
Únicamente B. amyloliquefaciens aislado 60 (tabla 2) fue capaz de crecer con el sustrato que contenía ACC como única fuente de N, lo que puede atribuirse a la actividad de la ACC desaminasa. La ACC desaminasa regula la producción de etileno a través de la degradación del ACC, precursor intermediario de la biosíntesis de etileno, en α-cetobutirato y amonio34,35,82. La presencia de ACC desaminasa ha sido mencionada para numerosas especies microbianas de bacterias gram negativas, gram positivas y hongos81, y, si bien los estudios para el género Bacillus son escasos, Read et al.78 la reportaron para este género bacteriano. En el caso de. B. amyloliquefaciens aislado 60, además fue productor de AIA, de modo que habría un efecto adicional, ya que el AIA activa a la ACC sintasa, esto favorecería la producción de ACC, y por ende, de etileno, el cual es un inhibidor del crecimiento de raíces. Sin embargo, las bacterias que producen la ACC desaminasa reducen el etileno y consecuentemente la inhibición del crecimiento de las raíces, tomando ventaja de esto y utilizando el ACC como una fuente de N84. De esta forma, en presencia de ambos mecanismos, las bacterias tendrán un mayor potencial como PGPR.
Los resultados de este estudio demostraron que bacterias rizosféricas y endófitas de E. nitens estimulan la emergencia y el crecimiento de plántulas de esta especie en condiciones de invernadero. Algunas bacterias PGPR presentaron más de un mecanismo de acción asociado a la promoción del crecimiento en plantas.
Estos resultados permiten predecir el potencial de estas bacterias para ser utilizadas como estimuladores de crecimiento de E. nitens en vivero y, de esta manera, favorecer la posterior supervivencia tras su establecimiento en terreno.
Responsabilidades éticasProtección de personas y animales. Los autores declaran que para esta investigación no se han realizado experimentos en seres humanos ni en animales.
Confidencialidad de los datos. Los autores declaran que en este artículo no aparecen datos de pacientes.
Derecho a la privacidad y consentimiento informado. Los autores declaran que en este artículo no aparecen datos de pacientes.
Conflictos de interesesLos autores declaran no tener ningún conflicto de intereses.
A Forestal Mininco S.A. por el financiamiento y las facilidades para realizar los muestreos y proporcionar las semillas para los ensayos, a la Lic. Evelyn Bustos del Laboratorio de Química de Productos Naturales de la Facultad de Ciencias Naturales de la Universidad de Concepción, por la colaboración con los análisis de HPLC, al Dr. Gonzalo Durán de la Universidad de Basel (Suiza), por la ayuda con el análisis estadístico y, finalmente, a la Dra. Marta Fernández del Centro de Biotecnología de la Universidad de Concepción por la ayuda con el estudio filogenético.
