




Los estudios de la Organización Mundial de la Salud indican que las personas viudas muestran menores índices de salud física y mental que el resto de la población de la misma edad cronológica. Por otra parte, la viudez y el aislamiento social son condiciones frecuentes en los mayores; sin embargo, las mujeres son las que presentan mayor incidencia, dada su mayor esperanza de vida media. Así, el objetivo del presente trabajo fue crear un modelo animal de soledad durante la vejez para estudiar en éste los cambios en el perfil cognitivo, conductual e inmunitario a los que conduce el aislamiento social en esta etapa de la vida.
Materiales y métodosVeinte ratones hembras de la cepa C57b/129sv estabulados en grupos de 4 a 5 animales hasta su vejez (18 meses de edad) siguieron en grupo (grupo control, n=10) o se aislaron a partir de ese momento y hasta alcanzar 24 meses de edad (ratones aislados, n=10). A esta edad avanzada los animales realizaron una serie de pruebas para valorar neofobia (test de esquinas), ansiedad (campo abierto) y aprendizaje y memoria (laberinto acuático de Morris). Posteriormente, se sacrificó a los animales, se obtuvo el timo y de éste la suspensión celular en la que se analizó la capacidad citotóxica de las células natural killer (NK) frente a la línea tumoral murina YAC-1.
ResultadosEl estudio conductual puso de manifiesto que los animales aislados durante la vejez muestran un declive funcional y cognitivo, con aumento de la neofobia y de la ansiedad así como problemas de aprendizaje y memoria. Además, la soledad conduce a una disminución de la actividad de las células NK tímicas.
ConclusionesSe demuestra la importancia del aislamiento y de la soledad durante la vejez, lo que exacerba la involución mental e inmunitaria durante este período a pesar de haber mantenido una vida social normal durante las etapas vitales anteriores.
Several studies by the World Health Organization indicate that widows and widowers show lower physical and mental health indexes than the age-matched general population. In addition, widowhood and social isolation are common in the elderly, with women being more affected than men due to their longer life span. Thus, the aim of the present study was to create an animal model of solitude in old age to study the behavioral, cognitive and immunological changes induced by social isolation at this late stage of life.
Material and methodsTwenty female C57b/129sv mice, housed in groups of 4–5 until their old age (18 months), remained in groups (controls, n=10) or were isolated after reaching the age of 18 months and until they reached the age of 24 months (isolated, n=10). At this advanced age, the animals were submitted to a battery of tests to assess neophobia (corner test), anxiety (open-field test), and learning and memory (Morris water maze). Thereafter, the animals were sacrificed and the thymus was removed. The natural killer (NK) activity of the thymic cells against the YAC-1 murine tumor cell line was evaluated.
ResultsAnimals isolated during old age showed functional and cognitive decline, with increased neophobia and anxiety as well as learning and memory deficits. In addition, isolation reduced the NK activity of thymic cells.
ConclusionsWe demonstrate the importance of social isolation and solitude during old age. Both social isolation and solitude exacerbate mental and immunological involution during this period, despite normal social life during previous stages of life.
Es bien conocido que el envejecimiento conduce a un deterioro conductual muy marcado, de modo que los individuos de edad avanzada muestran dificultades para afrontar situaciones novedosas de la vida diaria (aunque éstas no supongan peligro alguno) y muestran mayores grados de ansiedad, lo que los autores de este artículo junto con otros autores han demostrado tanto en humanos1,2 como en ratones3,4. De igual forma, numerosas aportaciones científicas han evidenciado el declive cognitivo en el aprendizaje y la memoria que se produce en ambos al avanzar la edad5,6.
Por otra parte, el sistema inmunitario parece ser especialmente vulnerable al proceso de envejecimiento7, mientras que su deterioro se encuentra íntimamente relacionado con la morbimortalidad asociada a la edad debido al declive de las funciones defensivas esenciales que este sistema ejerce en el organismo. El riesgo de presentar infecciones y la gravedad de éstas aumentan en la vejez, al igual que la susceptibilidad al cáncer8. Esto último puede ser, en parte, consecuencia del deterioro en la función antitumoral de las células natural killer (NK) que se da con la edad9,10. Además, la capacidad de preservar una función inmunológica adecuada ha demostrado ser un buen marcador de salud y predictor de longevidad11, lo que queda evidenciado en el caso de los sujetos centenarios12.
El aislamiento social es una situación frecuente en los mayores. El anciano siente una falta de compañía, de afecto y de apoyo que se agrava por la carencia de relaciones sociales de calidad. Entre las posibles causas se encuentran varias circunstancias a las que el anciano debe hacer frente en esta etapa de la vida: la jubilación, la dificultad para encontrar amistades nuevas, la existencia de cierto rechazo por parte de la sociedad y, de forma muy destacada, la viudez y la pérdida de otros seres queridos. Todo esto conduce a una privación de apoyo emocional, físico y económico ante la que el anciano tiene menor capacidad de respuesta.
Estudios de la Organización Mundial de la Salud indican que las personas viudas muestran menores índices de salud física y mental que el resto de la población de la misma edad cronológica13; sin embargo, las mujeres son las más afectadas debido a su mayor esperanza de vida media14. Así, el aislamiento social se considera un factor de riesgo en el desarrollo de enfermedades en los mayores15 o en las discapacidades que se presentan como resultado de éstas16 (en consecuencia, también es un factor de riesgo de morbimortalidad17). Dada la íntima comunicación que hay entre los sistemas nervioso e inmunológico18, la desregulación inmunitaria que se produce en los mayores, tanto per se como por consecuencia del deterioro del sistema nervioso, puede ser un elemento clave que deteriora la salud a la hora de afrontar todas estas emociones negativas mencionadas con anterioridad19.
En vista de lo comentado anteriormente, el objetivo del presente trabajo fue crear un modelo animal de soledad durante la vejez y estudiar en éste los cambios en algunos parámetros del perfil cognitivo, conductual e inmunitario a los que puede conducir el aislamiento social en esta etapa de la vida.
Material y métodosModelo animalSe emplearon 20 ratones hembras de la cepa C57b/129sv estabulados en el Departamento de Psiquiatría y Medicina Legal del Instituto de Neurociencias de la Universitat Autònoma de Barcelona. Los animales vivieron en grupos de 4 a 5 hasta su vejez (18 meses de edad). A partir de ese momento se mantuvieron en grupo (grupo control, n=10) o aislados hasta alcanzar 24 meses de edad (ratones aislados, n=10).
En las condiciones de estabulación se usaron jaulas makrolon iv estándares (35×35×25cm) con un lecho de 10cm de serrín y con agua y comida ad libitum (dieta de acuerdo con las recomendaciones del American Institute of Nutrition para animales de laboratorio). Las condiciones de temperatura (22 ± 2°C), humedad (entre el 50 y el 60%) y luz (ciclos de 12h, con encendido de luces a las 9.00) se controlaron diariamente.
Los animales se trataron en todo momento de acuerdo con los principios descritos en las European Community Council Directives (1201/2005 EEC).
Estudios conductualesA la edad avanzada de 24 meses, los animales realizaron una serie de pruebas conductuales estandarizadas, útiles para detectar alteraciones conductuales en roedores20 y que valoran neofobia (test de esquinas), ansiedad (campo abierto) y aprendizaje y memoria (laberinto acuático de Morris). El día anterior al inicio de estas pruebas, los animales se identificaron mediante rotulación de la cola con un número blanco. El orden de los animales en las distintas pruebas se contrabalanceó por la situación «ratones en grupo y ratones solos». Dentro de los animales en grupo, se eligió uno de cada jaula antes de escoger otro animal de la misma jaula con el fin de minimizar la manipulación repetida. Las pruebas conductuales se realizaron en la misma sala de estabulación (test de esquinas) o en una sala contigua (campo abierto y laberinto acuático de Morris) y siempre dentro de un rango horario estrecho (entre las 10.00 y las 13.00) para evitar efectos debido al ritmo circadiano.
Test de esquinasEl test de esquinas utilizado para evaluar la neofobia o el miedo a la novedad consiste en colocar al animal en el centro de una jaula cuadrangular idéntica a las que se utilizan para la estabulación. La jaula está descubierta y llena de un lecho de 10cm de serrín. El experimentador observa la conducta espontánea del animal y anota el número de esquinas que el animal visita en un período de 30s. Se utiliza una jaula distinta para cada animal, para evitar interferencias en esta conducta debido al rastro de olor de otro animal.
Campo abiertoUna vez finalizado el test de esquinas, el animal permanece en la jaula y se lo traslada a una sala contigua pintada de negro en la que se evalúa durante 5min su conducta en un «campo abierto» (caja de madera de fabricación artesanal de 50×50×25cm) de color blanco e iluminado (20lx). Estas circunstancias no resultan cómodas para el animal, por lo que genera cierta aversión frente a éstas. Así, el tiempo que el ratón necesita para salir del área central (latencia del centro) o para iniciar la conducta de acicalamiento (latencia de grooming) son medidas estándares que permiten valorar su grado de emotividad y de ansiedad. Después de efectuar la prueba a cada animal, la caja se limpia rigurosamente con agua y alcohol.
Laberinto acuático de MorrisEl laberinto acuático de Morris21 permite realizar distintas tareas de aprendizaje y de memoria espacial. El laberinto consiste en una piscina de 90cm de diámetro llena de agua a temperatura templada (25°C) hasta 15cm por debajo del límite superior de la pared y en la que es posible colocar una plataforma no visible sumergida 1cm por debajo de la altura del agua, que permite al animal subir a ésta a descansar y así acabar la prueba.
Cada día, los animales se trasladan a la sala en la que se realiza esta prueba y en la que hay 20 jaulas individuales (35×35×25cm) con 10cm de serrín, una para cada animal. En cada sesión, se introduce el animal con delicadeza en la piscina en uno de los puntos cardinales (norte, sur, este y oeste) escogido previamente de forma aleatorizada. La primera tarea que realizan los animales en el laberinto es el «aprendizaje de la posición de la plataforma». Consiste en aprender y recordar dónde está la plataforma (fija en medio del cuadrante sudeste) durante 4 sesiones diarias espaciadas en 15min (memoria a corto plazo) y durante varios días consecutivos (memoria a largo plazo), hasta que los animales encuentran la plataforma en un mínimo de tiempo que además es idéntico (llegan a la asíntota) durante 2 días consecutivos.
Una vez que los animales han aprendido en qué consiste la prueba y dónde está la plataforma, se los expone a 2 pruebas nuevas22 que consisten en 4 ensayos consecutivos de «memoria de trabajo»23 en los que el animal se introduce en la piscina 5s después de haber realizado el ensayo anterior. En cada una de éstas, la posición de la plataforma es distinta a la que tuvo durante la prueba anterior y se les concede un máximo de 90s para encontrarla (dado que se espera que inicialmente los animales continúen buscando la plataforma allí donde estuvo durante los 4 días previos y, por tanto, necesitan más tiempo para encontrarla).
En la primera prueba, la plataforma sumergida ha cambiado de posición respecto a los días anteriores, se encuentra ahora a 180° (en medio del cuadrante noroeste) y los animales entran en la piscina desde el sur. El segundo día, la plataforma se encuentra en el centro de la piscina y los animales entran por el este. Para finalizar, se realiza una cuarta prueba de «aprendizaje de plataforma visible» para contrastar diferencias de atención o de motivación, que consiste en 4 ensayos distanciados por 15min (entradas por el oeste, sur, norte y este, respectivamente), en la que se mide el tiempo que el animal necesita para subir a la plataforma. Tienen un máximo de 60s cuando la plataforma está visible (elevada 0,5cm respecto a la altura del agua), en posición noroeste y señalizada de forma vistosa con una bandera a rayas negras y blancas. En todas las pruebas, cuando el animal no encuentra la plataforma en el tiempo estipulado, el experimentador lo coloca en ésta durante 10s, exactamente el mismo tiempo que permanecen los animales en ésta cuando la encuentran por sí solos.
Estudios inmunológicosObtención de las células inmunitariasUna vez finalizadas las pruebas de conducta, se sacrificó a los animales por dislocación cervical de acuerdo con las directrices descritas en las European Community Council Directives (86/6091 EEC). Se obtuvo el timo de forma aséptica, se liberó de las impurezas visibles y se trasladó inmediatamente en medio RPMI 1640 frío (PAA, Laboratories GmbH, Pasching, Austria) al Departamento de Fisiología (Fisiología Animal II) de la Universidad Complutense de Madrid. Una vez allí, se homogeneizó de forma manual en un medio de solución salina de fosfato (PBS), presionando con suavidad a través de una malla metálica (Sigma, St. Louis, EE. UU). La suspensión de las células tímicas resultante se lavó y se determinó su número mediante microscopia óptica. Se ajustó posteriormente a 106células/l en medio RPMI 1640 sin rojo fenol (PAA, Laboratories GmbH, Pasching, Austria) para la realización del análisis. La viabilidad celular, medida de forma habitual antes y después de cada experimento a través del test de exclusión Azul Tripán (Sigma, St. Louis, EE. UU), fue superior al 98% en todos los casos.
Actividad antitumoral de las células natural killerSe empleó un método colorimétrico enzimático para medir la lisis de las células diana (Cytotox 96 TM Promega, Boeringher Ingelheim, Alemania) basado en la determinación de la actividad de la enzima lactatodeshidrogenasa mediante sales de tetrazolio10. Brevemente, el análisis consistió en incubar las células diana (células YAC-1 procedentes de un linfoma murino; 104 células/pocillo) junto con las células efectoras (células del timo; 105 células/pocillo) en placas de cultivo de 96 pocillos con fondo en «U» (Nunclon, Dinamarca). Las placas se centrifugaron a 250g durante 4min para facilitar los contactos celulares y se incubaron durante 4h a 37°C en una atmósfera húmeda y al 5% de dióxido de carbono. Se empleó medio RPMI 1640 sin rojo fenol que no interfiere con la determinación colorimétrica. Tras la incubación, se midió la actividad de la enzima lactatodeshidrogenasa en 50μl del sobrenadante de cada pocillo mediante la adición del sustrato de la enzima y la posterior medida de la absorbancia a 490nm. Además, se llevaron a cabo varias medidas de control para conocer la lisis espontánea de las células diana y de las células efectoras, así como la lisis total de las células diana.
Los resultados se expresaron como porcentaje de lisis. Para su cálculo, se siguió la ecuación que se indica a continuación: lisis (%)=[(E-ES-TS)/(M-TS)]×100 donde E es la media de las absorbancias en presencia de células efectoras y de células diana, ES es la media de las absorbancias en los pocillos de células efectoras solas, TS es la media de las absorbancias en los pocillos de células diana solas y M es la media de las absorbancias máximas obtenidas tras la incubación de las células diana con solución de lisis.
EstadísticaEn primer lugar, se comprobó la normalidad de las muestras mediante la prueba de Kolmogorov–Smirnov. Las diferencias entre los 2 grupos experimentales se analizaron mediante el test de la t de Student para muestras independientes. En el laberinto acuático de Morris, las diferencias entre grupos durante los distintos ensayos o durante los distintos días se evaluaron con el análisis ANOVA (análisis de la variancia) para medidas repetidas, mientras que las comparaciones entre grupos en un ensayo concreto se evaluaron con el test de la t de Student para muestras independientes. En todos los casos, la significación estadística se consideró a partir de p<0,05.
ResultadosEstudios conductualesEl estudio conductual pone de manifiesto que los animales aislados durante la vejez muestran un declive funcional y cognitivo con un aumento en pautas de neofobia y ansiedad, así como problemas de aprendizaje y memoria.
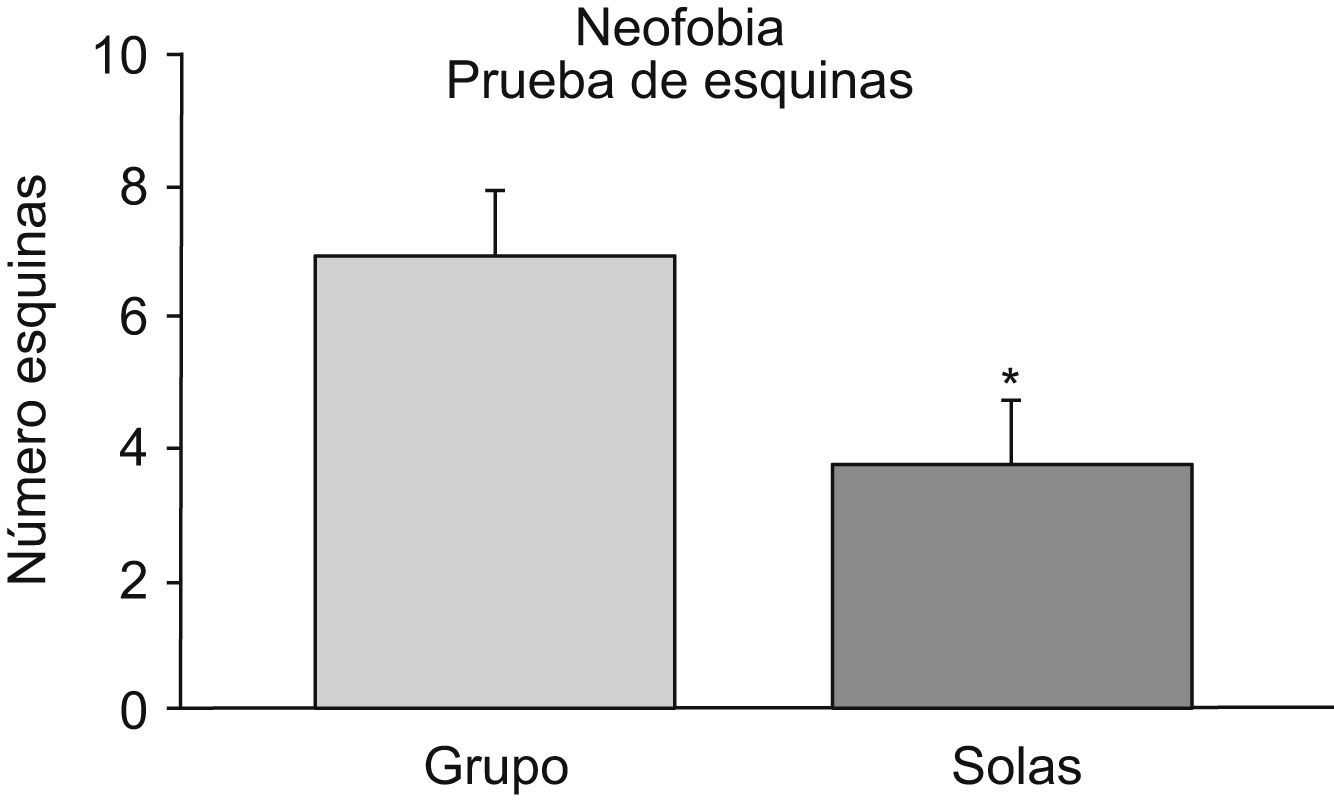
Como se indica en la figura 1, los animales que se aislaron en la vejez realizaron menos exploraciones de las esquinas de la jaula nueva que aquellos que continuaron estabulados en grupo (test de la t de Student; p<0,05).
 y aisladas en la vejez (solas). Cada columna representa la media±error estándar de 10 valores correspondientes a 10 animales. *p<0,05 respecto al valor obtenido en los animales mantenidos en grupo.")
Prueba de las esquinas para valorar la neofobia. Se representa el número de esquinas que visitan ratones hembras viejas mantenidas en grupo (grupo) y aisladas en la vejez (solas). Cada columna representa la media±error estándar de 10 valores correspondientes a 10 animales. *p<0,05 respecto al valor obtenido en los animales mantenidos en grupo.
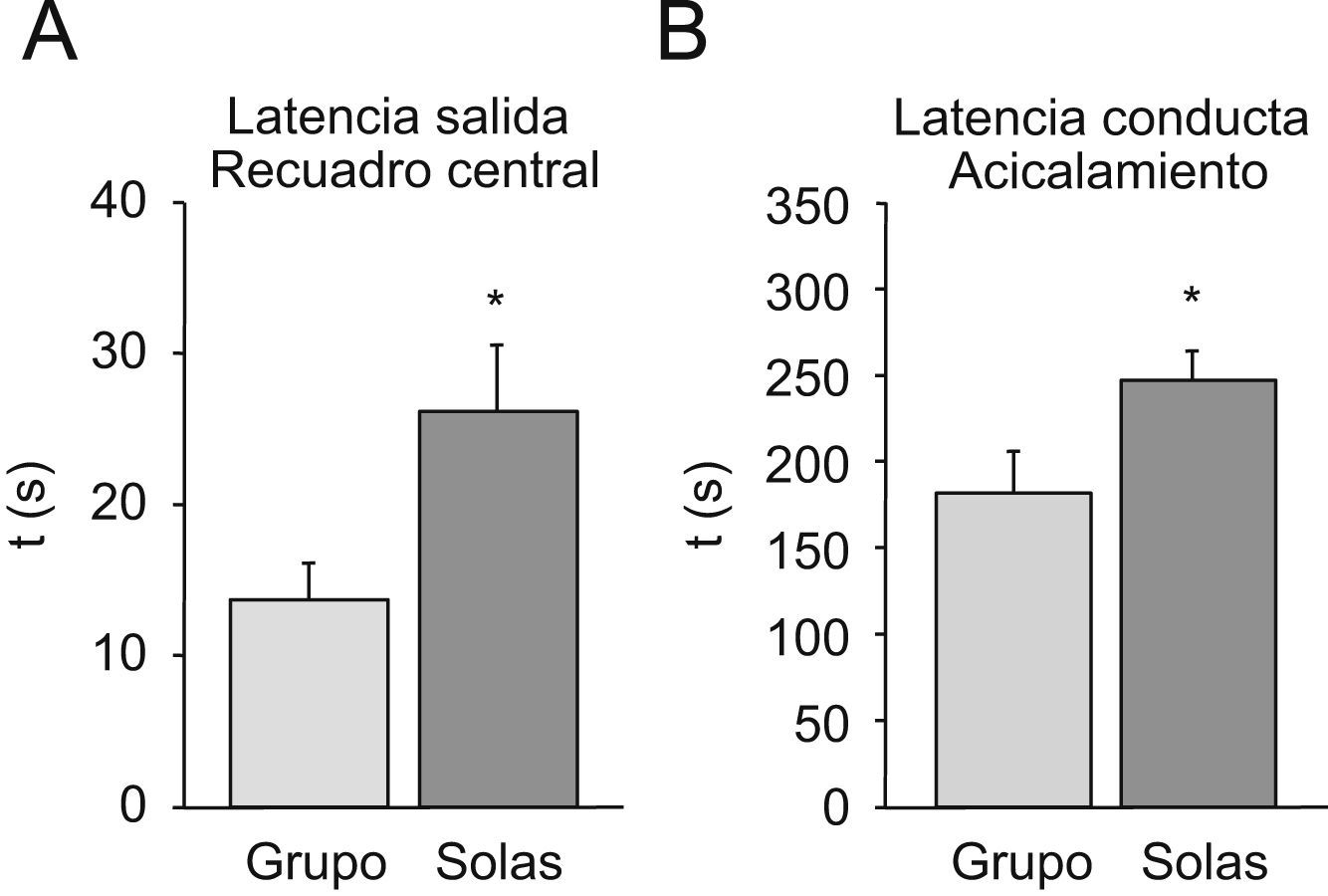
En el campo abierto (fig. 2), el análisis de las latencias de salida del recuadro central (fig. 2A) o de la conducta de acicalamiento (fig. 2B) indica que los animales mantenidos en aislamiento tienen valores superiores a los animales mantenidos en grupo (test de la t de Student; p<0,05 en ambos casos).
 Se representa la latencia en segundos de la salida del recuadro central. B) Se representa la latencia en segundos de la conducta de acicalamiento en ratones hembras viejas mantenidas en grupo (grupo) y aisladas en la vejez (solas). *p<0,05 respecto al valor obtenido en los animales mantenidos en grupo.")
Prueba de campo abierto para valorar la ansiedad y la emotividad. Cada columna representa la media±error estándar de 10 valores correspondientes a 10 animales. A) Se representa la latencia en segundos de la salida del recuadro central. B) Se representa la latencia en segundos de la conducta de acicalamiento en ratones hembras viejas mantenidas en grupo (grupo) y aisladas en la vejez (solas). *p<0,05 respecto al valor obtenido en los animales mantenidos en grupo.
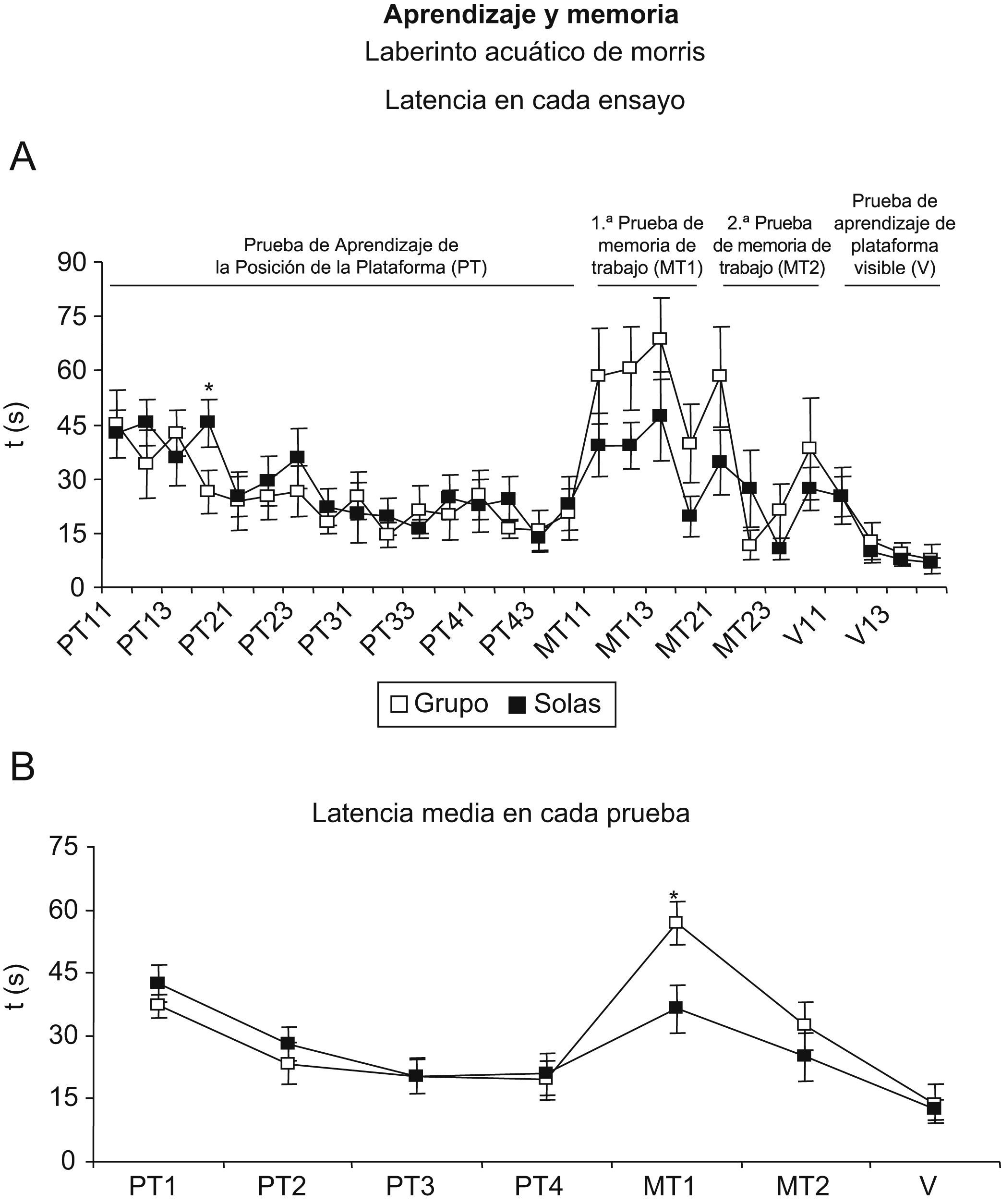
En la figura 3A se ilustran los resultados por ensayos diarios de las pruebas realizadas en el laberinto acuático de Morris. En la prueba de aprendizaje de la posición de la plataforma, los 2 grupos experimentales muestran curvas de aprendizaje similares durante los 4 días que dura la prueba, pero cuando se analizan los 4 ensayos realizados en cada uno de los 4 días se observa que la diferencia entre el primer y el último ensayo del primer día es positiva en los animales aislados, con una tardanza o un aumento del tiempo de 2,9 ± 5,3s respecto a la mejora de −18,9 ± 10,6s mostrada por los animales mantenidos en grupo. Esto se traduce en una diferencia estadísticamente significativa cuando se compara el tiempo que ambos grupos necesitan para encontrar la plataforma en este último ensayo del primer día (test de la t de Student; p<0,05). Además, en la primera prueba de memoria de trabajo, en la que se cambió la posición de la plataforma respecto a los 16 ensayos anteriores, los animales en grupo muestran mayores tiempos comparativamente a los animales aislados (test de la t de Student, p<0,05) mientras que en la segunda prueba de memoria de trabajo ya no se aprecian diferencias entre los 2 grupos experimentales (fig. 3B). Esto evidencia que los animales en grupo mantienen el recuerdo de la posición de la plataforma durante los 4 días anteriores y continúan buscándola allí donde estuvo en esos días, mientras que los animales aislados la encuentran con la misma dificultad que cuando realizaron la primera prueba en el laberinto. Una vez que los animales aprenden que la posición de la plataforma puede cambiar respecto a días anteriores, las diferencias entre ambos grupos desaparecen.
 en ratones hembras viejas mantenidas en grupo (grupo) y aisladas en la vejez (solas). Cada punto representa la media±error estándar de 7 a 9 valores correspondientes a ese mismo número de animales. *p<0,05 respecto al valor obtenido en los animales mantenidos en grupo. A) En cada ensayo. B) Media de cada prueba. MT1 y MT2) Pruebas de memoria de trabajo, días uno y 2, respectivamente; cada día consta de 4 sesiones. PT1, PT2, PT3 y PT4) Prueba de la adquisición de la plataforma, días uno a 4, respectivamente; cada día consta de 4 sesiones. V) Prueba de la plataforma visible que consta de 4 sesiones.")
Laberinto acuático de Morris para valorar el aprendizaje y la memoria (latencias por segundos) en ratones hembras viejas mantenidas en grupo (grupo) y aisladas en la vejez (solas). Cada punto representa la media±error estándar de 7 a 9 valores correspondientes a ese mismo número de animales. *p<0,05 respecto al valor obtenido en los animales mantenidos en grupo. A) En cada ensayo. B) Media de cada prueba. MT1 y MT2) Pruebas de memoria de trabajo, días uno y 2, respectivamente; cada día consta de 4 sesiones. PT1, PT2, PT3 y PT4) Prueba de la adquisición de la plataforma, días uno a 4, respectivamente; cada día consta de 4 sesiones. V) Prueba de la plataforma visible que consta de 4 sesiones.
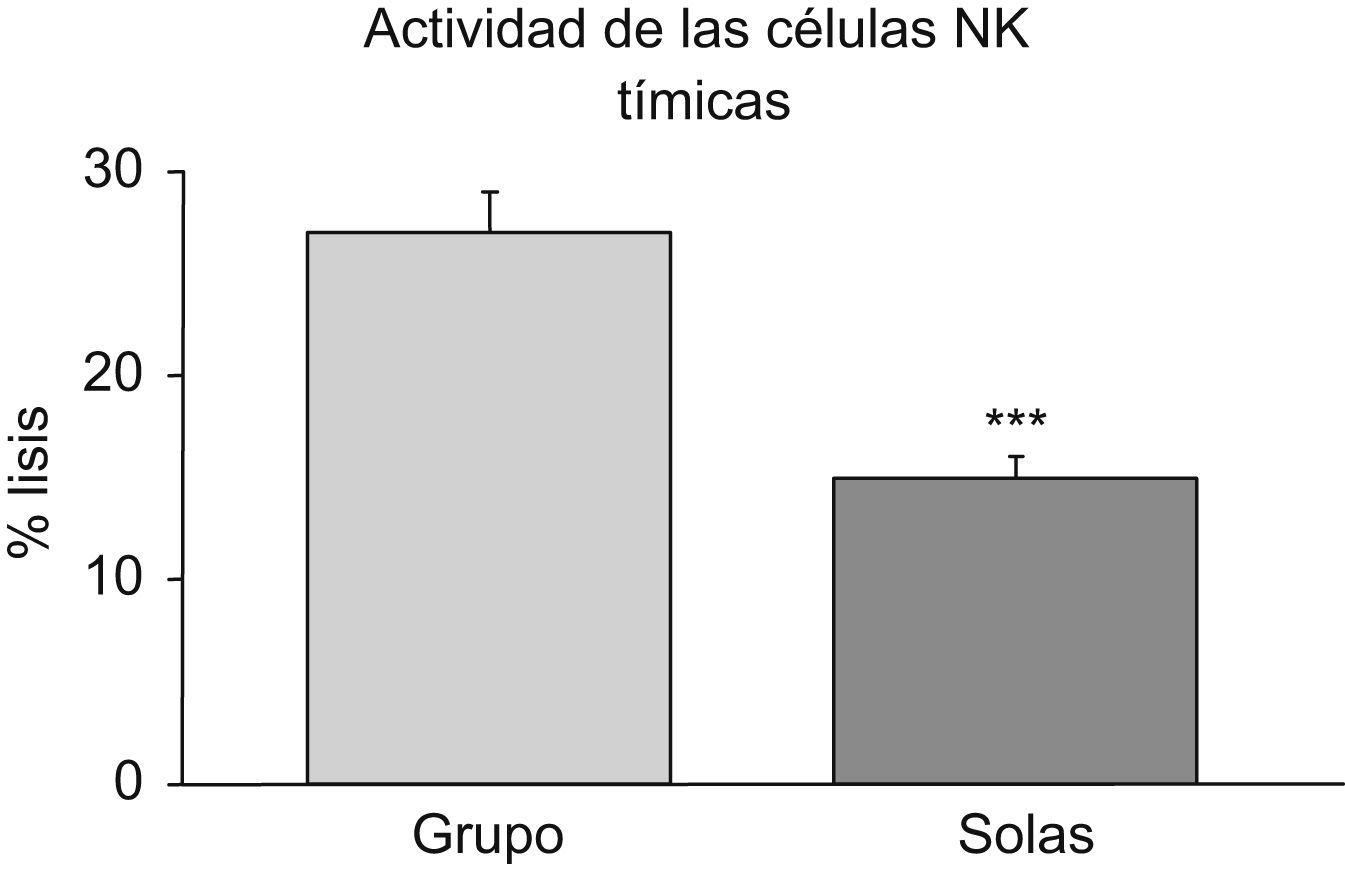
Por otra parte, la soledad en la vejez conduce a una disminución estadísticamente significativa de la actividad lítica de las células NK tímicas (grupo control: 27 ± 2%, ratones aislados: 15 ± 1%; p<0,001) (fig. 4).
 de las células natural killer (NK) tímicas procedentes de ratones hembras viejas mantenidas en grupo (grupo) y aisladas en la vejez (solas). Cada columna representa la media±error estándar de 7 a 10 valores correspondientes a ese mismo número de animales. Cada valor es la media de ensayos realizados por triplicado. *p<0,001 respecto al valor obtenido en los animales mantenidos en grupo.")
Actividad lítica frente a células tumorales (% lisis) de las células natural killer (NK) tímicas procedentes de ratones hembras viejas mantenidas en grupo (grupo) y aisladas en la vejez (solas). Cada columna representa la media±error estándar de 7 a 10 valores correspondientes a ese mismo número de animales. Cada valor es la media de ensayos realizados por triplicado. *p<0,001 respecto al valor obtenido en los animales mantenidos en grupo.
Este trabajo presenta el primer modelo animal que aborda experimentalmente una situación común para muchas personas mayores que por distintas razones experimentan la soledad durante su vejez tardía a pesar de haber gozado de una vida social activa en etapas anteriores.
En humanos, el declive cognitivo y físico asociado a la edad está caracterizado por mayores tiempos de reacción, disminución de la capacidad de atención y de concentración así como peor memoria a corto y largo plazo; la novedad y el grado de dificultad son los factores más relevantes que condicionan el declive24.
El modelo animal de soledad en el envejecimiento muestra disminución de respuestas en la prueba de las esquinas para la neofobia y aumento de latencias en el campo abierto. Esto es indicativo de que el individuo de edad avanzada aislado socialmente tiene más dificultades para desenvolverse con normalidad en ambientes nuevos o potencialmente ansiogénicos que el individuo que continúa viviendo en comunidad. Justamente estas variables son las mismas que se encuentran afectadas en modelos animales mutantes que muestran ansiedad y menor supervivencia20, y son también las variables que en ratones triple transgénicos 3xTgAD sirven para describir cambios similares a algunos de los síntomas neuropsiquiátricos típicos asociados a la enfermedad de Alzheimer, incluso se anticipan a la aparición de las primeras deficiencias cognitivas leves y avanzan en paralelo con el posterior curso de la demencia25,26. Todo esto demuestra que los individuos expuestos a la soledad en su edad avanzada pueden tener una mayor vulnerabilidad a los trastornos mentales.
La capacidad para adaptarse a situaciones nuevas que además implican funciones cognitivas superiores, como el aprendizaje y la memoria, también es menor en los animales viejos aislados. En el primer día de test en el laberinto acuático de Morris, los animales no muestran ninguna mejora durante los 4 ensayos, lo que significa que no consiguen entender en qué consiste la tarea que deben realizar o que no recuerdan con igual eficacia dónde está la plataforma sumergida. Sin embargo, la repetición de la tarea en días sucesivos ayuda a optimizar los resultados y reduce las diferencias entre los 2 grupos experimentales, probablemente porque la repetición de la tarea implica la reducción de la novedad y de la dificultad y ayuda a los procesos de consolidación de la memoria. Es decir, a pesar de presentar mayor dificultad inicial, los individuos viejos aislados mejoran con la práctica.
Estos resultados se reiteran cuando en la segunda y la tercera prueba se exige a los animales aprender una nueva localización de la plataforma. Se observa claramente que todos los animales requieren más tiempo para encontrarla porque no se localiza allí donde la encontraron durante los 16 ensayos realizados en los días anteriores. En general, los animales de edad avanzada mantenidos en grupo social necesitan más tiempo para encontrar la plataforma en la nueva localización que los animales aislados, probablemente porque recuerdan mejor la antigua posición e insisten más en buscarla en la zona en que estuvo.
En la tercera prueba la situación de posible cambio de la plataforma parece predecible y ambos grupos muestran igual eficacia en su búsqueda. Es interesante observar que cuando la plataforma está visible, los 2 grupos experimentales invierten exactamente el mismo tiempo en encontrar la plataforma, como ocurrió en los días 3 y 4 de su adquisición, lo que indica que en situaciones controladas y de baja dificultad (la plataforma se encontraba en un lugar conocido a causa de la práctica, visible y muy bien indicada) los animales viejos aislados no se diferencian de los animales mantenidos en grupo.
Por tanto, generalmente el aislamiento en la vejez afecta a la capacidad de respuesta a situaciones novedosas, potencialmente ansiogénicas y que requieren funciones superiores de aprendizaje y memoria, pero los individuos mejoran con la disminución de la novedad o con la disminución del grado de dificultad y se benefician de la práctica.
Paralelamente al deterioro cognitivo y físico asociado a la edad, la vejez comporta cambios funcionales en el sistema inmunitario que se conocen como inmunosenescencia. Dada la esencial función defensora frente a agentes extraños y antitumoral que este sistema ejerce en el organismo, el mantenimiento de una respuesta inmunitaria adecuada se considera como un excelente marcador de salud e incluso se ha relacionado con la longevidad de los individuos11. Es bien sabido que al envejecer aumenta la incidencia de infecciones y cáncer8; enfermedades relacionadas directamente con la funcionalidad de los leucocitos y más concretamente con la actividad citotóxica natural de las células NK en el caso de procesos víricos y tumorales. De hecho, esta función parece ser especialmente sensible al proceso de envejecimiento y demuestra ser un buen predictor de longevidad en modelos animales27.
Por otra parte, el sistema nervioso y el sistema inmunitario se encuentran íntimamente relacionados y forman una compleja red de interacciones bidireccionales que mantiene la homeostasis del organismo y, por tanto, la salud18. Dada esta comunicación estrecha entre ambos, factores ambientales como el estrés, ya sea por aislamiento, exposición a relaciones sociales excesivamente jerárquicas u otras situaciones que implican adaptación a cambios ambientales extremos, dañan la función inmunológica del individuo. Del mismo modo, la exposición continua a estos factores puede derivar en estados de ansiedad crónica, una alteración que tiene un fuerte impacto sobre las funciones leucocitarias claves, como lo es la actividad antitumoral de las células NK28.
De los resultados obtenidos en el presente trabajo se puede deducir que ambas condiciones (envejecimiento y aislamiento) unidas en las últimas etapas de la vida tienen un efecto considerable sobre la actividad de las células NK, ya que esta función disminuye muy significativamente en los animales viejos aislados al compararlos con los animales mantenidos en grupo, en los que el deterioro es menor. Esto puede conducir a un declive acelerado de la salud y por tanto a una muerte prematura de los individuos de edad avanzada que se encuentran solos, de donde se deduce la importancia de preservar lazos sociales de calidad en la vejez. Del mismo modo, estrategias de promoción de la función inmunológica, como el ejercicio moderado29 o el enriquecimiento ambiental30, es decir, mantener una vida activa tanto física como mentalmente, pueden ser beneficiosas en personas mayores que tengan que afrontar la soledad.
ConclusionesSe ha demostrado la importancia del aislamiento y de la soledad durante la vejez, lo que exacerba el deterioro mental e inmunitario durante este período a pesar de haber mantenido una vida social normal durante las etapas vitales anteriores. Además, los resultados sugieren que la realización de estrategias terapéuticas dirigidas a potenciar la interacción social y a promocionar la actividad física puede resultar beneficiosa para paliar el declive neuroinmunitario acelerado que se observa en los individuos aislados de edad avanzada.
FinanciaciónMEC (BFU2005-06777; SAF2006-13642), RETICEF (RD06/0013/0003) del ISCIII y una ayuda del vicerrectorado de investigación de la UAB para Grupos Emergentes (EME2006-140335) y de la UCM para el Grupo de Investigación (910379ENEROINN) han financiado este trabajo.
Este trabajo ha recibido el premio «Pañella Casas» 2007 a la mejor comunicación en forma oral del Área Biológica presentada durante el 49.o Congreso Nacional de la Sociedad Española de Geriatría y Gerontología, celebrado en Palma de Mallorca del 6 al 9 de junio de 2007.
